Outi H. Manninen  ,
Rainer Peltola
,
Rainer Peltola
Effects of picking methods on the berry production of bilberry (Vaccinium myrtillus), lingonberry (V. vitis-idaea) and crowberry (Empetrum nigrum ssp. hermaphroditum) in Northern Finland
Manninen O. H., Peltola R. (2013). Effects of picking methods on the berry production of bilberry (Vaccinium myrtillus), lingonberry (V. vitis-idaea) and crowberry (Empetrum nigrum ssp. hermaphroditum) in Northern Finland. Silva Fennica vol. 47 no. 3 article id 972. https://doi.org/10.14214/sf.972
Highlights
- Berry production of bilberry, lingonberry and crowberry was studied after picking the berries by plastic hand rake, long-handed metal rake, and powerful picking by long-handed metal rake
- Berry production was not affected by the damage caused by any of the picking method
- Current commercial picking methods do not endanger the berry production of the berry species at least in short-term.
Abstract
The effect of commercial wild berry picking on berry yields is under a strong public debate in Finland. Especially high concern has been arisen over damages caused by metal rakes used in commercial picking to subsequent berry production. We studied the berry production of bilberry (Vaccinium myrtillus L.), lingonberry (V. vitis-idaea L.) and crowberry (E. nigrum ssp. hermaphroditum (Hagerup) Böcher) after picking the berries by 1) plastic hand rake, 2) long-handed metal rake and 3) powerful picking by long-handed metal rake, in northern Finland during 2010–2012. In the powerful long-handed metal rake treatment the aboveground vegetation was raked twice to the moss layer after berry picking. Biomass, which was removed from the vegetation by rakes was collected and used as a measure of the damage. We assumed that picking by plastic hand rake would result in lowest, long-handed metal rake intermediate and powerful picking by long-handed metal rake highest biomass loss from vegetation. The amount of biomass loss should in turn be reversely reflected into berry production. However, only the powerful picking by long-handed metal rake removed higher amount of biomass than other picking methods in bilberry and lingonberry. In crowberry, the amount of biomass removed by rakes increased from treatment to treatment. Contrary to our assumption, berry production of bilberry, lingonberry and crowberry was not affected by the damage caused by any of the picking method. We conclude that long-handed metal rake used in commercial picking is comparable to hand rake in terms of berry production.
Keywords
berry yields;
commercial picking;
wild forest berries
-
Manninen,
Agrifood Research Finland MTT, Eteläranta 55, FI-96300 Rovaniemi, Finland
E-mail
outi.h.manninen@mtt.fi
- Peltola, Agrifood Research Finland MTT, Eteläranta 55, FI-96300 Rovaniemi, Finland E-mail rainer.peltola@mtt.fi
Received 29 January 2013 Accepted 16 May 2013 Published 11 September 2013
Views 91958
Available at https://doi.org/10.14214/sf.972 | Download PDF

1 Introduction
Wild berry picking and utilization has long tradition in many European countries (Seeland and Staniszewski 2007; Turtiainen and Nuutinen 2012). In Finland, berry picking is a popular leisure activity (Turtiainen and Nuutinen 2012). Wild berries may also have high economical significance as additional incomes for households which collect berries for sale (Salo 1995; Saastamoinen et al. 2000; Jonsson and Uddstål 2002). Bilberry (Vaccinium myrtillus L.) and lingonberry (Vaccinium vitis-idiae L.) are the most widely collected berry species in Finland (Maaseutuvirasto 2012).
In Finland, the average annual (1997–2008) yields of bilberry and lingonberry are approximately 180 and 260 million kg, respectively (Turtiainen et al. 2011). Despite the high between year variation in wild berry yields (Miina et al. 2009; Turtiainen et al. 2011), the national utilization rates of the berries are relatively constant between good and poor yields, being up to 6% of the annual yield for bilberry and 10% for lingonberry in Finland (Salo 2008; Turtiainen et al. 2011). These rates are based on data collected before migrant pickers were introduced into Finland in 2005 to meet the increasing wild berry demand and may underestimate the present use of wild berries (Turtiainen et al. 2011). However, newer estimations tell that 3.1 million kg of bilberry and 8.5 of lingonberry was bought by biggest berry companies in Finland in 2011, which are more than average for the last three decades (Maaseutuvirasto 2012). The yield of crowberry (Empetrum nigrum L.) is rather even between years (Wallenius 1999), but the annual purchase of crowberries was only 0–0.4 million kg during last decade (Maaseutuvirasto 2012). As these numbers do not include the current household use of wild berries, the actual use of berries is higher.
Nowadays, commercial berry picking relies on foreign labour in Finland and Sweden (Richards and Saastamoinen 2010; Rantanen and Valkonen 2011). The phenomenon of foreign pickers has raised numerous sociological and ecological questions. From ecological point of view, the replacement of plastic hand picking rakes by long-handed rakes with metal tines has raised concern over sustainability of berry picking. As flower buds of bilberry, lingonberry and crowberry develop year before flowering, powerful autumn harvest may injure developing flower organs and thereby decrease subsequent berry production. Apart from the direct damaging effect on flower organs, biomass loss by rakes may reduce the number of reproductive units of the plant species (Salemaa et al. 1999; García and Ehrlén 2002) or enhance the vegetative growth which in turn leads to lower investments to berry production in following years (Tolvanen et al. 1993; Hegland et al. 2005; Stark et al. 2010).
Although current wild berry resources exceed their utilization in Finland (Turtiainen et al. 2011), strong public debate on possible negative effect of the commercial picking on the subsequent berry production clearly indicates the need for closer examination of the issue. We conducted a factorial experiment with two factors for bilberry, lingonberry and crowberry (E. nigrum ssp. hermaphroditum (Hagerup) Böcher) in northern Finland. The factors were year (three levels: 2010, 2011 and 2012) and picking treatments (four levels: unpicked control, picking by plastic hand rake, picking by long-handed metal rake and powerful picking by long-handed metal rake). In the powerful metal raking treatment the aboveground vegetation was raked twice to the moss layer to specify the vulnerability of berry production. To measure the damage caused by picking rakes on the wild berry species, their biomass which was removed by rakes were collected with berries, and the berry production was later monitored in two consecutive years. Berry production was evaluated by measuring the number and fresh weight of the berries. We assumed that the level of damage would gradually increase from hand raking to metal raking and to powerful metal raking, which in turn should be reversely reflected into berry production.
2 Material and methods
2.1 Study sites
The study was carried out in northern boreal forest zone in Rovaniemi, northern Finland during 2010–2012. The study was conducted at three locations: the bilberry study site was located at Petäjävaara (66°19´N, 25°29´E), and lingonberry and crowberry sites at Kivalo (66°21´N, 26°39´E and 66°20´N, 26°40´E, respectively). The studies of bilberry and lingonberry were conducted in closed coniferous forests, representing moist Hylocomium-Myrtillus forest type (Cajander 1949). Location of the study plots within the study sites were visually determined based on the dominance of the species, i.e. bilberry dominated the study plots in bilberry, and lingonberry in the lingonberry study site. The study of crowberry was conducted at open heath habitat at the top of a smooth hill where crowberry forms almost pure understorey vegetation. Nomenclature follows Hämet-Ahti et al. (1998).
Mean monthly temperature and precipitation are presented based on the measurements by the Finnish Meteorological Institute (Apukka weather station, 66°58´N, 26°01´E, Fig. 1). The weather station is located approximately 35 km from the bilberry study site and 40 km from the lingonberry and crowberry study sites. Mean monthly temperature during the growing season (May-September) was higher in 2011 than in other years (Fig. 1). Mean monthly precipitation decreased towards to the end of the growing season after July in 2010. Mean monthly precipitation was high in the beginning of the growing season in 2011 and 2012 (Fig. 1).
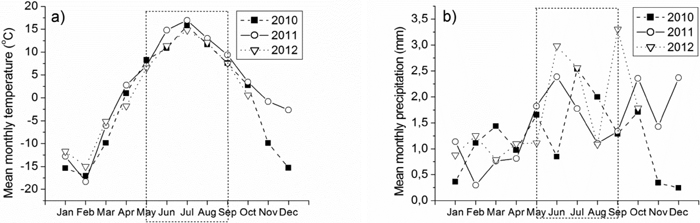
Fig. 1. a) Mean monthly temperature (°C) and b) mean monthly precipitation (mm) in Rovaniemi during 2010–2012. Weather data for November and December are not available in 2012. Growing season is represented by rectangles in the figures.
2.2 Experimental design
The experiment was conducted in a factorial design with two factors for each berry species during 2010–2012. The factors were year (three levels) and picking treatments (four levels). Treatments were applied in randomly assigned, permanent 1 x 1 m study plots, established at least five meters apart in 2010. The plots were further randomly assigned to the piking treatments (n = 8). Picking treatments were (1) unpicked control, (2) picking by plastic hand picking rake, (3) picking by long-handed metal rake and (4) powerful picking by long-handed metal rake. In the powerful metal rake picking treatment, the aboveground vegetation was raked twice to the moss layer. These treatments are referred to hereafter as control, hand raking, metal raking and powerful metal raking, respectively. More detailed descriptions of the rakes used in the experiment are given in Fig. 2.

Fig. 2. Picture of a) the plastic hand rake and b) long-handed metal rake used in the experiment. The width of the plastic hand rake was 21.5 cm with flat, pointed, gradually widening plastic tines. The space between the tines was 1 cm at the point and 0.5 cm at the base of the tines. The width of the metal rake was 20 cm and had evenly distributed round (diameter 2.5 mm) metal tines, located 0.5 cm apart.
In 2010, the biomass of the each species which was removed from the plots by rakes was collected, dried at 80 °C for 24 hours, and weighed. The total undamaged biomass of each berry species was measured by cutting aboveground biomass samples (n = 8) at randomly selected 1 x 1 m areas outside the study plots at each site, and treated as the biomass which was removed by rakes. Percentage values were calculated to determine the biomass which was removed by rakes (= the biomass removed from the picking treatment plots by rakes in 2010/the total undamaged biomass in 2010 x 100) for each berry species.
Prior to treatments in 2010 the berries in the control plots were counted without picking them and berries in hand raking, metal raking and powerful metal raking plots were hand-picked, dried at room temperature for two hours, counted and their pooled fresh weight was recorded.
In 2011, the number of berries in the control and picking treatment plots were counted in the field without picking them. In 2012, the berries in all plots were hand-picked and treated as in 2010. After this, the aboveground biomass was collected by cutting from the plots, dried at 80 °C for 24 hours, and weighed.
The treatments, biomass collections and counting the berries were carried out in the middle of August, in the middle of September and in the late of August for bilberry, lingonberry and crowberry, respectively.
Berry wholesalers generally freeze the stock material collected by berry pickers. This material contains berries, litter and a mixed plant material. The first refining step of the stock material is mechanical separation of frozen reject (= litter and mixed plant material) from the berries. To estimate the amount of bilberry biomass which is removed during commercial picking, frozen reject from berry refining plant was obtained from Korvatunturin Marja (Inc., Savukoski). From the reject, six samples (50 g fresh weight) were taken and bilberry ramets, bilberries, litter and other species biomass were separated. These fractions were kept at room temperature for five hours, and their fresh weight was measured. Samples were later dried at 80 °C for 24 hours, and weighed. The fresh weight and dry weight of bilberry ramet and bilberry biomass in reject samples were later used to compare the biomass loss as percentage values due to the genuine commercial picking to the results of this study.
2.3 Statistical analysis
The difference between the amounts of biomass removed from the plots by rakes in 2010 was tested using the one-factor ANOVA for each berry species. Differences between the number of berries of bilberry, lingonberry and crowberry between years (2010, 2011 and 2012) and picking treatments (control, hand raking, metal raking, and powerful metal raking) were tested using the repeated measures ANOVA. Year was treated as a within-subject factor and picking treatments as a between-subject factor in the analysis.
The mean fresh weight of the berries was calculated by dividing the pooled fresh weight of the berries by their number in each study plot before the statistical analysis. Berries were left in the control plots in 2010, therefore the differences between the mean fresh weights of the berries were possible to test only between picking treatments. This was done by using the repeated measures ANOVA, using year (2010 and 2012) as a within-subject factor and picking treatments (hand raking, metal raking, and powerful metal raking) as a between-subject factor. To test whether the mean fresh weight of the berries differed between the control and picking treatment plots in 2012, i.e. two years after the treatments, the one-factor ANOVA was applied. The biomass which was collected from the control and picking treatment plots in 2012 was tested by the one-factor ANOVA.
In the repeated measures ANOVA, when Mauchly’s test of sphericity was significant, Huynh-Feldt adjusted F values were used. The data were checked for the requirements of normality, and homogeneity of variances was tested by Levene’s test before statistical analysis. Log-transformed data were used to achieve the requirements of homogeneity when testing the amount of removed biomass from the plots by rakes. Tukeys’ post hoc-test (significance level p < 0.05) was used in multiple comparisons between the control and picking treatments. Statistical analyses were carried out using SPSS/PASW Statistics 17 for Windows (SPSS Inc. Chicago 2009).
3 Results
The biomass removed from the picking treatment plots by rakes differed significantly for all species (Fig. 3). The removed biomass was lower in the hand and metal raking treatments than in the powerful metal raking treatment in bilberry (F(2,21) = 6.911, p = 0.005) and lingonberry (F(2,21) = 47.577, p ≤ 0.001) (Fig. 3). In crowberry, the removed biomass was lowest in the hand raking treatment, intermediate in the metal raking treatment and highest in the powerful metal raking treatment (F(2,21) = 31.795, p ≤ 0.001) (Fig. 3). The total undamaged biomass of bilberry, lingonberry and crowberry was approximately 143.17 ± 17.3, 196.51 ± 20.5 and 308.92 ± 22.0 (dry weight m–2, mean ± SE), respectively, at the study sites in 2010. Bilberry lost approximately 0.03%, 0.16% and 1.15% of its biomass in the hand raking, metal raking and powerful metal raking treatments in 2010, respectively. Corresponding values for lingonberry are 0.08%, 0.07% and 2.20%, and for crowberry 0.34%, 2.28% and 6.79%, respectively.
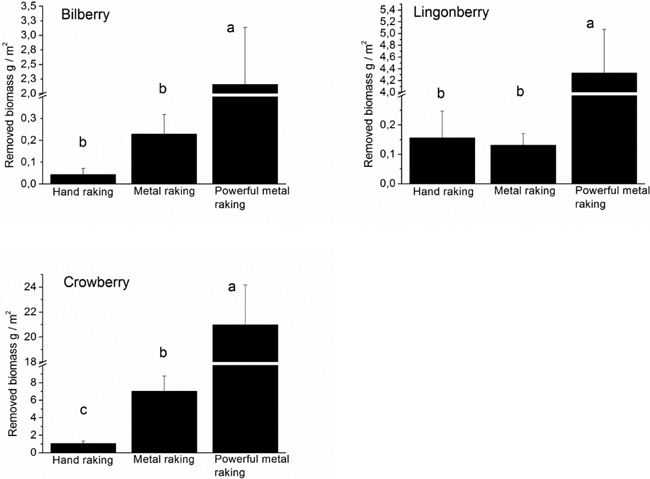
Fig. 3. The amount of the biomass of bilberry, lingonberry and crowberry (dry weight g/m2, mean ± SE) which was removed from the picking treatment plots by rakes in 2010. Different legends indicate significant differences between the treatments as revealed by the one-factor ANOVA followed by Tukey’s post hoc-test at the significance level p < 0.05.
Number of the berries of bilberry, lingonberry and crowberry varied between years in the plots, however, no differences were found between the picking treatments as revealed by the repeated measures ANOVA (Table 1). The number of berries of each species was highest in 2010 compared to other years (Fig. 4). The number of bilberry was lowest in 2011 compared to other years, whereas the number of lingonberry and crowberry decreased year by year during the study period (Fig. 4).
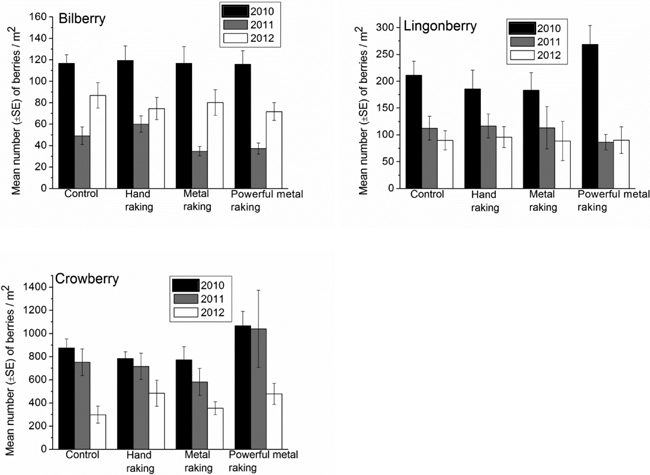
Fig. 4. Mean number of berries (± SE)/m2 of bilberry, lingonberry and crowberry in the control and the picking treatment plots during 2010–2012.
| Table 1. Results of the repeated measures ANOVA analyzing the effect of picking treatment on the number of the berries of bilberry, lingonberry and crowberry during 2010–2012. Picking treatment and year df = 3 and 2, respectively, and error df = 28 and 56, respectively. Year x picking treatment df = 6. | ||||||
| Year | Picking | Year x Picking | ||||
| F | p | F | p | F | p | |
| Bilberry | 79.080 | < 0.001 | 0.384 | 0.766 | 0.835 | 0.548 |
| Lingonberry | 43.517 | < 0.001 | 0.139 | 0.936 | 1.947 | 0.089 |
| Crowberry a) | 14.957 | < 0.001 | 2.179 | 0.113 | 0.528 | 0.753 |
| a) Huynh-Feldt adjusted F values were used in the analysis. | ||||||
| Table 2. Results of the repeated measures ANOVA analyzing the difference between picking treatments on the fresh weight of the berries of bilberry, lingonberry and crowberry in 2010 and 2012. Picking treatment and year df = 2 and 1, respectively, and error df = 21 and 21, respectively. Year x picking treatment df = 2. | ||||||
| Year | Picking | Year x Picking | ||||
| F | p | F | p | F | p | |
| Bilberry | 115.271 | < 0.001 | 1.017 | 0.379 | 2.893 | 0.078 |
| Lingonberry | 297.142 | < 0.001 | 0.854 | 0.440 | 2.374 | 0.118 |
| Crowberry | 5.428 | 0.030 | 0.162 | 0.851 | 0.349 | 0.710 |
The mean fresh weight of the berries in the treatment plots in 2010 were significantly higher than in 2012, however, no differences were found between the picking treatments as revealed by the repeated measures ANOVA (Tables 2 and 3). In 2012, two years after the treatments, both the mean fresh weight of the berries (Table 3) and the aboveground biomass of the berry species were similar in the control and picking treatment plots (one-factor ANOVAs, statistics not shown).
| Table 3. Pooled fresh weight of the berries (g/m2) and mean fresh weight of the berry in treatment plots in 2010 and 2012. | ||||||
| Bilberry | Lingonberry | Crowberry | ||||
| Mean ± SE | Mean ± SE | Mean ± SE | ||||
| Pooled fresh weight | Fresh weight of the berry | Pooled fresh weight | Fresh weight of the berry | Pooled fresh weight | Fresh weight of the berry | |
| 2010 | ||||||
| Control | not measured | not measured | not measured | not measured | not measured | not measured |
| Hand rake | 37.32 ± 4.6 | 0.31 ± 0.01 | 49.38 ± 7.5 | 0.27 ± 0.01 | 222.21 ± 20.8 | 0.28 ± 0.01 |
| Metal rake | 31.25 ± 4.7 | 0.21 ± 0.01 | 52.01 ± 8.3 | 0.28 ± 0.01 | 212.24 ± 31.1 | 0.27 ± 0.01 |
| Powerful metal rake | 32.31 ± 5.6 | 0.27 ± 0.02 | 68.93 ± 7.5 | 0.27 ± 0.01 | 287.94 ± 25.6 | 0.28 ± 0.01 |
| 2012 | ||||||
| Control | 16.45 ± 3.3 | 0.17 ± 0.02 | 18.37 ± 3.1 | 0.21 ± 0.00 | 91.00 ± 24.3 | 0.30 ± 0.03 |
| Hand rake | 13.38 ± 2.5 | 0.17 ± 0.02 | 20.19 ± 4.0 | 0.21 ± 0.00 | 122.66 ± 26.8 | 0.26 ± 0.02 |
| Metal rake | 14.70 ± 2.5 | 0.18 ± 0.01 | 17.57 ± 6.6 | 0.20 ± 0.01 | 91.76 ± 16.6 | 0.25 ± 0.01 |
| Powerful metal rake | 13.28 ± 2.4 | 0.18 ± 0.01 | 17.21 ± 4.8 | 0.19 ± 0.01 | 128.46 ± 25.9 | 0.27 ± 0.01 |
The amount of dry biomass of bilberry which was removed in the powerful metal raking treatment (approximately 2 g m–2, Fig. 3) coarsely corresponded to 4 g m–2 fresh weight biomass loss in 2010. In the same year, the average pooled fresh weight of the bilberries in the hand rake, metal rake and powerful metal rake treatments (values are shown in Table 3) was approximately 34 g m–2. The total fresh weight biomass loss of bilberry, i.e. the fresh weight of the biomass which was removed by rakes and the fresh weight of the bilberries, was therefore approximately 38 g m–2 in the powerful metal rake treatment in 2010. This means that the amount of bilberry ramet biomass expressed as fresh weight was approximately 10% from the total fresh weight removed by rake in the powerful metal rake treatment in this study in 2010.
4 Discussion
Contrary to our assumption, the berry production of bilberry, lingonberry and crowberry was not affected by damage caused by any of the treatments during the study period. Even the powerful metal raking did not result to diminished berry production. Corresponding studies on the effects of picking methods on wild berry production are missing. However, the response in berry production to picking may be similar to those found for herbivory given that both berry picking and grazing by herbivores remove certain amount of biomass of target species (Bråthen and Oksanen 2001). Both natural and artificial herbivory experiments suggest that herbivory reduces the berry production of wild berry species, especially in bilberry (Fernández-Calvo and Obeso 2004; Hegland et al. 2005, 2010; Mathisen et al. 2010 but see also Pato and Obeso 2012). Biomass loss caused by herbivory directly decreases the amount of reproductive units of plant, decreasing thereby future reproduction (Garcia and Ehrlén 2002; Côte et al. 2004). Moreover, biomass loss may indirectly enhance the allocation of resources to vegetative growth at the expense of sexual reproduction, postponing the berry production of plant (Tolvanen 1993; Hegland et al. 2006; Stark et al. 2010).
The fact that damages caused by picking did not affect the berry production of wild berry plants in this study may be explained by the small amount of biomass removed by treatments, and the timing of the damage. In this experiment, the most severe picking treatment (powerful metal raking) removed the biomass of bilberry, lingonberry and crowberry approximately 1, 2 and 7%, respectively. Berry production of bilberry has been found to decrease for two years after 20% biomass loss (Stark et al. 2010), suggesting that the intensity of the damage in this study was not severe enough to affect negatively the bud number even in the most powerful picking treatment. Contrary to the investigation of Stark et al. (2010), in which 20% biomass loss corresponded 5–10 cm cutting from the top of the bilberry ramets, Pato and Obeso (2012) reported that cutting 5–8 cm did not affect the number of flowers, berry production nor berry weight of bilberry. Moreover, it has been shown that damages occurring early in the growing season have the most detrimental effect on several plant species, whereas the biomass loss occurring late in the growing season is more easily compensated (Tolvanen and Taulavuori 1998; Salemaa et al. 1999; Whigham and Chapa 1999; García and Ehrlén 2002). Taken together, it seems that both low intensity as well as timing of the damage buffered the possible negative effect of picking on the berry production in this study.
Mechanical separation of litter and a mixed plant material from frozen berries is the first refining step before berries are delivered to markets. Typically, share of bilberry ramet biomass is about 4–8% fresh weight in unrefined mass (Jari Huttunen, manager of Korvatunturin Marja, pers. comm. in 2013). This falls below the share of bilberry ramet biomass which was removed (10%) in the powerful metal rake treatment in our study. This suggests that the current commercial picking methods do not endanger berry production, as the most powerful raking in our experiment did not decrease berry production.
Biomass loss may change the competitive balance between plant species in long term. Mathisen et al. (2010) reported that long lasting simulated herbivory increased the sexual reproduction in the understorey in boreal forest in general. However, the change in the understorey vegetation structure due to the competitive interactions decreased the cover and sexual reproductive units of bilberry. Generally, deciduous species such as bilberry are resistant for moderate biomass loss due to their relatively fast growth rate and effective utilization of reserved carbohydrates (Tolvanen 1994; Tolvanen et al. 1994; Hegland et al. 2010). Evergreen species such as lingonberry and crowberry express slow growth rate (Ruiz-Robleto and Villar 2005 and references therein) and are therefore more sensitive to biomass loss than deciduous species. In this study, we did not measure the biomass of wild berry species one year after picking. However, plant species biomass exhibited full recovery in second year after raking, suggesting that biomass loss of wild berry species caused by rake picking have no long term influences on vegetation structure and competition balance between the species in this study. Furthermore, the finding that the mean fresh weight of the berries was similar in the control and picking treatment plots two years after raking supports the results that picking treatments had no effect on berry production. This is in accordance with the results by Pato and Obeso (2012), which showed that simulated herbivory by clipping did not affect the fruit mass of bilberry during the similar time period as in this study.
Berry production of wild berry species varies greatly between years (Wallenius 1999; Kudo and Suzuki 2002; Miina et al. 2009; Turtiainen et al. 2011) as also revealed in this study. In this study, between year variation in berry production was similar in the control and picking treatments, suggesting that other than picking method determined the berry production. According to generally accepted view, the variation in wild berry yields result from climatic conditions, such as temperature, precipitation and frost injuries during different stages of berry production (Raatikainen and Vänninen 1988; Tolvanen 1997; Wallenius 1999; Selås 2000; Krebs et al. 2009; Gerdol et al. 2013). In this study, wild berry production was highest in 2010 compared to other years. This result disagrees with the nationwide announcements of wild berry yields by Finnish Forest Research Institute in 2010–2012, and describes the difficulty of yield estimation of wild berry species due to the high regional variation. The yield of bilberry suffered from drought in July in Finland in 2011 (Finnish Forest Research Institute) which probably explain the low number of the berries of bilberry in our experiment in that year. Contrary to bilberry, the number of berries of lingonberry and crowberry decreased year by year in this study. Different between year variation in berry production suggests that the response of berry yields to climatic variables may be species specific (Krebs et al. 2009). Moreover, both current and previous year’s climatic variables may affect the berry production of long-lived wild berry species (Raatikainen and Vänninen 1988; Wallenius 1999; Selås 2000; Krebs et al. 2009). It has been observed that weather conditions during flower organ development in the autumn are equally important for berry production than weather conditions during the other stages of flower and berry development (Selås 2000). Climatic variables also affect indirectly through pollinator activity to the berry production (Jacquemart and Thompson 1996). It also has been suggested that the berry production of Vaccinium-species may fluctuate regularly (Vander Kloet and Cabilio 1996; Wallenius 1999; Selås 2000). For example, Selås (2000) reported peak in the production of berries at intervals of 2–5 years for bilberry, and this periodicity was dependent on previous year’s berry production and weather conditions.
In summary, plastic hand rake and metal rake removed equal amount of bilberry and lingonberry biomass except when the metal rake usage was exceptionally powerfully. Crowberry was more sensitive to picking by metal rake than bilberry and lingonberry measured by biomass loss. However, even the powerful usage of metal rake did not decrease berry production of any of the wild berry species in this study. Therefore we conclude that long-handed metal rake used in commercial wild berry picking seems to be comparable to plastic hand rake in terms of berry production. However, in this study picking treatments were conducted only once, and although their effects were monitored in two consecutive years, the study provides only the short-time insight on the issue. If same areas are continuously under high picking pressure, repeated biomass loss may affect negatively the berry production of wild berry species for example through deleterious effect on plant resource level (Fröborg 1996; Kudo and Suzuki 2002; Strengbom et al. 2003) or through plant competitive interactions (Mathisen et al. 2010).
Acknowledgements
We thank Esko Saastamoinen, Hanna-Leena Juntti, Aarno Niva and Pasi Rautio for assistance, and two anonymous reviewers for useful comments and suggestions. This work was financed by European Agricultural Fund for Rural Development.
References
Bråthen K.A., Oksanen J. (2001). Reindeer reduce biomass of preferred plant species. Journal of Vegetation Science 12: 473–480. http://dx.doi.org/10.2307/3236999.
Cajander A.K. (1949). Forest types and their significance. Acta Forestalia Fennica 56. 71 p.
Côte S.D., Rooney T.P., Tremblay J.-P., Dussault C., Waller D.M. (2004). Ecological impacts of deer overabundance. Annual Review of Ecology and Systematics 35: 113–147. http://dx.doi.org/10.1146/annurev.ecolsys.35.021103.105725.
Fernández-Calvo I.C., Obeso J.R. (2004). Growth, nutrient content, fruit production and herbivory in bilberry Vaccinium myrtillus L. along an altitudinal gradient. Forestry 77: 213–223. http://dx.doi.org/10.1093/forestry/77.3.213.
Finnish Forest Research Institute. Metsäkasvien fenologia ja satoennusteet muuttuvassa ilmastossa tiedotteet. [In Finnish]. http://www.metla.fi/hanke/3517/tiedotteet.htm. [Cited 23 Jan 2013].
Fröborg H. (1996). Pollination and seed production in five boreal species of Vaccinium and Andromeda (Ericaceae). Canadian Journal of Botany 74: 1363–1368. http://dx.doi.org/10.1139/b96-165.
García M., Ehrlén J. (2002). Reproductive effort and herbivory timing in a perennial herb: fitness components at the individual and population levels. American Journal of Botany 89: 1295–1302. http://dx.doi.org/10.3732/ajb.89.8.1295.
Gerdol R., Siffi C., Iacumin P., Gualmini M., Tomaselli M. (2013). Advanced snowmelt affects vegetative growth and sexual reproduction of Vaccinium myrtillus in a sub-alpine heath. Journal of Vegetation Science 24: 569–579. http://dx.doi.org/10.1111/j.1654-1103.2012.01472.x.
Hämet-Ahti L., Suominen J., Ulvinen T., Uotila P. (eds.). (1998). Retkeilykasvio (Field Flora of Finland). Finnish Museum of Natural History, Botanical Museum, Helsinki. 656 p.
Hegland S.J., Rydgren K., Seldal T. (2006). The response of Vaccinium myrtillus to variations in grazing intensity in a Scandinavian pine forest on the island of Svanøy. Canadian Journal of Botany 83: 1638–1644. http://dx.doi.org/10.1139/b05-132.
Hegland S.J., Jongejans E., Rydgren K. (2010). Investigating the interaction between ungulate grazing and resource effects on Vaccinium myrtillus populations with integral projection models. Oecologia 163: 695–706. http://dx.doi.org/10.1007/s00442-010-1616-2.
Jacquemart A.L., Thompson J.D. (1996). Floral and pollination biology of three sympatric Vaccinium (Ericaceae) species in the upper Ardennes, Belgium. Canadian Journal of Botany 74: 210–221. http://dx.doi.org/101139/b96-025.
Jonsson L., Uddstål R. (2002). En beskrivning av den svenska skogsbärbranschen. [In Swedish]. Skog & trä. Swedish University of Agricultural Sciences, Vindeln Experimental Forest, Umeå. 25 p. ISBN 91-973519-5-4.
Krebs C.J., Boonstra R., Cowcill K., Kenney A.J. (2009). Climatic determinants of berry crops in the boreal forest of the southwestern Yukon. Botany 87: 401–408. http://dx.doi.org/10.1139/B09-013.
Kudo G., Suzuki S. (2002). Relationships between flowering phenology and fruit-set of dwarf shrubs in Alpine fellfields in northern Japan: a comparison with a subarctic heathland in northern Sweden. Arctic, Antarctic, and Alpine Research 34: 185–190. http://dx.doi.org/10.2307/1552470.
Maaseutuvirasto. [Agency for Rural Affairs]. (2012). Luonnonmarjojen ja -sienten kauppaantulomäärät vuonna 2011. [In Finnish]. [Market supply of wild berries and edible mushrooms in 2011]. Seinäjoki. 40 p.
Mathisen K.M., Buhtz F., Danell K., Bergström R., Skarpe C., Suominen O., Persson I.-L. (2010). Moose density and habitat productivity affects reproduction, growth and species composition in field layer vegetation. Journal of Vegetation Science 21: 705–716. http://dx.doi.org/10.1111/j.1654-1103.2010.01180.x.
Miina J., Hotanen J.-P., Salo K. (2009). Modelling the abundance and temporal variation in the production of bilberry (Vaccinium myrtillus L.) in Finnish mineral soil forests. Silva Fennica 43: 577–593.
Pato J., Obeso J.R. (2012). Fruit mass variability in Vaccinium myrtillus as a response to altitude, simulated herbivory and nutrient availability. Basic and Applied Ecology 13: 338–346. http://dx.doi.org/10.1016/j.baae.2012.05.003.
Raatikainen M., Vänninen I. (1988). The effects of the 1984–1985 cold winter on the bilberry and lingonberry yield in Finland. Acta Botanica Fennica 136: 43–47.
Rantanen P., Valkonen J. (2011). Ulkomaalaiset metsämarjanpoimijat Suomessa. [In Finnish]. Ulkoministeriön raportti, EU:n Ulkorajarahasto. 84 p. http://formin.finland.fi/public/download.aspx?ID=88464&GUID=%7B33EAB400-3CF7-4B9D-9AB3-84C5EE8FE8D8%7D. [Cited 1 Jan 2013].
Richards R.T., Saastamoinen O. (2010). NTFP policy, access to markets and labour issues in Finland: impacts of regionalization and globalization on the wild berry industry. In: Laird S., McLain R., Wynberg R. (eds.). Wild product governance – finding policies that work for non-timber forest products. Earthscan, People and Plants International Conservation Series. p. 287–309.
Ruiz-Robleto J., Villar R. (2005). Relative growth rate and biomass allocation in ten woody species with different leaf area longevity using phylogenetic independent contrasts (PICs). Plant Biology 7: 484–494. http://dx.doi.org/10.1055/s-2005-865905.
Saastamoinen O., Kangas K., Aho H. (2000). The picking of wild berries in Finland in 1997 and 1998. Scandinavian Journal of Forest Research 15: 645–650. http://dx.doi.org/10.1080/02827580050216897.
Salemaa M., Vanha-Majamaa I., Gardner P.I. (1999). Compensatory growth of two clonal dwarf shrubs, Arctostaphylos uva-ursi and Vaccinium uliginosum in a heavy metal polluted environment. Plant Ecology 141: 79–91. http://dx.doi.org/10.1023/A:1009847728799.
Salo K. (1995). Non-timber forest products and their utilization. In: Hytönen M. (ed.). Multiple-use forestry in the Nordic countries. Gummerus Printing, Jyväskylä. p. 117–155. ISBN 951-40-1421-9.
Salo K. (2008). Luonnontuotteet. [In Finnish]. In: Rantala S. (ed.). Tapion taskukirja. Metsäkustannus Oy, Helsinki. p. 240–250. ISBN 978-952-5694-26-0.
Seeland K., Staniszewski P. (2007). Indicators for a European cross-country state-of-the-art assessment of non-timber forest products and services. Small-scale Forestry 6: 411–422. http://dx.doi.org/10.1007/s11842-007-9029-8.
Selås V. (2000). Seed production of a masting dwarf shrub, Vaccinium myrtillus, in relation to previous reproduction and weather. Canadian Journal of Botany 78: 423–429. http://dx.doi.org/10.1139/b00-017.
Stark S., Niva A., Martz F., Vuorela E. (2010). Sustainable gathering of juniper and bilberry shoots for natural product industry. [In Finnish with English summary]. MTT, Jokioinen. p. 27. ISBN 978-952-487-269-0.
Strengbom J., Olofsson J., Witzell J., Dahlgren J. (2003). Effects of repeated damage and fertilization on palatability of Vaccinium myrtillus to grey sided voles, Clethrionomys rufocanus. Oikos 103:133–141. http://dx.doi.org/10.1034/j.1600-0706.2003.12680.x.
Tolvanen A. (1994). Differences in recovery between a deciduous and an evergreen Ericaceous clonal dwarf shrub after simulated aboveground herbivory and belowground damage. Canadian Journal of Botany 72: 853–859. http://dx.doi.org/10.1139/b94-110.
Tolvanen A., Taulavuori K. (1998). Timing of deacclimation affects the ability to recover from simulated winter herbivory. Plant Ecology 135: 9–12. http://dx.doi.org/10.1023/A:1009784917648.
Tolvanen A., Laine K., Pakonen T., Saari E., Havas P. (1993). Above-ground growth response of the bilberry (Vaccinium myrtillus L.) to simulated herbivory. Flora 188: 197–202.
Tolvanen A., Laine K., Pakonen T., Havas P. (1994). Responses to harvesting intensity in a clonal dwarf shrub, the bilberry (Vaccinium myrtillus L). Vegetatio 110: 163–169. http://dx.doi.org/10.1007/BF00033396.
Turtiainen M., Nuutinen T. (2012). Evaluation of information on wild berry and mushroom markets in European countries. Small-scale Forestry 11: 131–145. http://dx.doi.org/10.1007/s11842-011-9173-z.
Turtiainen M., Salo K., Saastamoinen O. (2011). Variations of yield and utilisation of bilberries (Vaccinium myrtillys L.) and cowberries (V. vitis-idaea L.) in Finland. Silva Fennica 45: 237–251.
Vander Kloet S.P., Cabilio P. (1996). Ten year study of the annual variation in berry and seed production in a population of Vaccinium corymbosum L. American Midland Naturalist 135: 349–356. http://dx.doi.org/10.2307/2426718.
Wallenius T.H. (1999). Yield variations of some common wild berries in Finland in 1956–1996. Annales Botanici Fennici 36: 299–314.
Whigham D.F., Chapa A.S. (1999). Timing and intensity of herbivory: Its influence on the performance of clonal woodland herbs. Plant Species Biology 14: 29–37. http://dx.doi.org/10.1046/j.1442-1984.1999.00004.x.
Total of 40 references
