Hannu Hökkä  ,
Hanna Mäkelä
,
Hanna Mäkelä
Post-harvest height growth of Norway spruce seedlings in northern Finland peatland forest canopy gaps and comparison to partial and complete canopy removals and plantations
Hökkä H., Mäkelä H. (2015). Post-harvest height growth of Norway spruce seedlings in northern Finland peatland forest canopy gaps and comparison to partial and complete canopy removals and plantations. Silva Fennica vol. 48 no. 5 article id 1192. https://doi.org/10.14214/sf.1192
Highlights
- Norway spruce seedlings’ height growth recovered within four years after the cutting of canopy gaps
- Growth was linearly related to tree height, being highest for tallest seedlings
- Seedlings in the 20 m diameter gap and in the central and northern parts in the 15 m diameter gap showed the best growth
- In gaps early height growth was 60% of that in peatland spruce plantations but 2–3 times higher than in uneven-aged cut forests.
Abstract
Recent studies have shown the establishment of Norway spruce (Picea abies (L.) Karst.) to be successful in small canopy gaps cut in drained spruce mire stands in northern Finland. The aim of this study was to quantify seedling height growth in gaps and compare it to that observed in other canopy cuttings and plantations. We sampled spruce crop seedlings (maximum density ca. 3000 ha–1) in the spring of 2013 in a field experiment in which canopy gaps of 10, 15 and 20 m in diameter had been cut in winter 2004. The total seedling height in 2013 and the length of annual shoots over the past five years (2012–2008) were recorded in the survey. Seedling height varied from 20 cm to 2.7 m, with an average of 65 cm. The average annual height growth was 7.1 cm. A mixed linear model analysis was carried out to investigate seedling height growth variation. Seedling height was linearly and positively related to growth. Height growth started to increase in the fifth growing season after cutting. Seedling height growth in the 20 m gap was slightly better than in the smaller ones. In the 15 m gap, both the centrally located seedlings and those located at the northern edge grew best. In the 20 m gap, southerly located seedlings grew more slowly than seedlings in all other locations. The average seedling height growth in this study was about 60% of that in peatland plantations, but comparable to that in mineral soil gaps, and 2–3 times higher than in uneven-age cut stands.
Keywords
Picea abies;
height growth;
peatlands;
growth models;
advance growth;
canopy gap
-
Hökkä,
Finnish Forest Research Institute, Rovaniemi Unit, Eteläranta 55, FI-96300 Rovaniemi, Finland
E-mail
hannu.hokka@metla.fi
- Mäkelä, Lapland University of Applied Sciences, Jokiväylä 11 C, FI-96300 Rovaniemi, Finland E-mail hanna.makela@edu.ramk.fi
Received 14 May 2014 Accepted 25 November 2014 Published 2 January 2015
Views 144644
Available at https://doi.org/10.14214/sf.1192 | Download PDF

1 Introduction
Traditionally, highly productive spruce mire stands have been regenerated using clear-cutting and effective site preparation methods like mounding or ploughing, and the planting of Norway spruce (Picea abies (L.) Karst.) seedlings (e.g. Moilanen et al. 1995). In clear-cut areas, however, problems related to competition from ground vegetation (Moilanen et al. 1995; Hånell 1992) and pioneer tree species, mainly pubescent birch (Betula pubescent Ehrh.), necessitate intensive early management by means of herbaceous control, tending and pre-commercial thinning. These measures increase the costs of the clear-cut method. Water protection issues related to clear-cut and site preparation (e.g. Nieminen 2004) have also raised interest in the possibilities of natural regeneration of drained peatland forests.
Recent studies suggest that in northern Finnish spruce mire stands, the cutting of small canopy gaps provides conditions where the natural establishment of Norway spruce seedlings (composing of both advance regeneration and germinated seedlings) can be sufficient within five years of cutting without any regeneration measures and costs (Hökkä et al. 2011, 2012). Seedling establishment, however, does not guarantee that successful regeneration has been accomplished. It is important to know the growth rate of the established seedlings; how much lower their growth is compared to that in clear-cut areas and how long the period of growth recovery is. If the seedlings continue their growth at a fairly low rate for decades, the regeneration period becomes excessively long, which may result in the form of significantly longer rotations and postponed cutting incomes. This may eventually compromise the benefits of non-existent regeneration costs.
A mass of studies have been investigating forest gap formation, seedling establishment, and seedling growth in gaps. However, the majority of those studies have been conducted in temperate or tropical forests and concern natural gap dynamics with different broadleaved tree species (cf. Yamamoto 2000; Coates and Burton 1997). In fact, there seems to be much more limited information available on timber harvesting methods that emulate natural gap dynamics as a way of regenerating boreal coniferous forests. Spruce seedling establishment and the density of regeneration stocking in natural and harvested gaps have been investigated in quite a lot of studies (e.g. Leemans 1991; Drobyshev and Nihlgård 2000; Hanssen 2003; Valkonen et al. 2011; Hökkä et al. 2012). However, very few studies have addressed the height growth dynamics of advance regeneration in canopy gaps.
One of the earliest studies on Norway spruce seedling height development after the cutting of canopy gaps in the boreal region was made by Cajander (1934). He found that in a southern Finland rich Oxalis-Myrtillus mineral soil site (according to Cajander (1926)), spruce advance regeneration showed slow recovery of growth in gaps larger than 0.01 ha, i.e., larger than ca. 12 m in diameter. If the gap diameter exceeded 25 m, all seedlings in the gap could respond and grow vigorously. Chantal et al. (2003) analysed light conditions and the early post-cutting development of Norway spruce and Scots pine seedlings that were seeded systematically in different locations in a 50 m diameter gap and under the canopy of the surrounding stand in a Myrtillus site (Cajander 1926). Due to the uneven distribution of light on the gap area, location-wise and species-wise differences in seedling biomass were found after two years. Drobyshev and Nihlgård (2000) showed that in natural gaps in southern boreal spruce stands, the growth of spruce seedlings was related to gap size as well as seedling size and location within the gap. Outside the boreal region, Coates (2000) concluded that several planted coniferous species showed an asymptotically increasing growth trend as the gap size increased from a small gap the size of couple of trees to one of 1000 m2 in the temperate coniferous forests of British Columbia. On boreal peatlands, the only information on spruce advance regeneration seedlings’ height growth in canopy gaps are the tentative results published by Hökkä and Repola (2012) from the same data used by Hökkä et al. (2011). The conclusion was that no differences in the average growth rate of the first five-year period in gaps of different sizes (78–314 m2) were found.
The size of the gap, as well as the seedling’s location within the gap due to the uneven distribution of light over the gap area, influence the seedling growth rate (e.g. Coates 2000; Drobyshev and Nilhgård 2000; Chantal et al. 2003). It can be assumed that three factors are related to the growing conditions in gaps of different sizes. The first is the higher amount of radiation, i.e. light and temperature available for trees in the large gaps, which has a positive impact on tree growth in places were radiation levels significantly increase (Page and Cameron 2006). Total precipitation reaching the ground also increases when compared to the uncut forest resulting in higher soil moisture in gaps (Page and Cameron 2006). Secondly, increased light also enhances competition for free resources. On a highly productive site, ground vegetation covers the site within two to four years (Moilanen et al. 1995) and may form a serious constraint for seedling growth and survival (Hånell 1992; Hånell 1993; Nilson and Lundqvist 2001). Rapidly-growing pioneer tree species like pubescent birch also benefit from increased light and create another aspect of competition (Moilanen et al. 1995; Roy et al. 2000). The third factor is the edge effect of the uncut forest, i.e., the root competition of trees growing at the gap edge also extends to the gap area (Kuuluvainen 1993; Chantal et al. 2003). The observed growth rate of advance growth reflects a combination of these factors which are not equally distributed over the gap area.
The cutting of canopy gaps in order to release the growth of advanced regeneration can be compared to other release cuttings, e.g., when shelter trees are removed to enable the development of natural advance growth. A decrease in competition and a change in light availability may however be significantly larger than in the case of gap cuttings, depending on the completeness of overstory removal. Results from studies from mineral soil sites have indicated rather long periods of slow spruce height growth after overstory removal. Koistinen and Valkonen (1993) and Valkonen (2000) proposed that spruce advance growth shows a clearly lower growth rate for four to five years after release cutting when compared to planted spruce seedlings in mineral soil sites in southern Finland. A similar lag in height growth has also been reported by Cajander (1934), Skoglefald (1967) and Bergan (1971). According to Valkonen (2000), it will take another five or more years for height growth to fully recover to a level similar to that of planted spruce. Örlander and Karlsson (2000) studied a shallow-peated drained spruce stand in southern Sweden and found that after cutting the spruce seedlings eight-year height growth rate was related to seedling height, height growth prior to cutting, and density of the retained overstory trees.
Another point of comparison could be found from cuttings in uneven-aged stands, in which selected dominant and co-dominant trees are harvested to enhance the growth of sub-canopy trees and improve conditions for the establishment of natural seedlings. Eerikäinen et al. (2014) investigated the growth rate of seedlings growing in uneven-aged cut stands in southern Finland and concluded that spruce seedling height growth was very slow, only a couple of centimetres annually and strongly related to tree height. They also found that cutting intensity influenced the growth of the seedlings. Lundqvist (1989) has reported comparably slow growth rates in Sweden after uneven-aged cutting.
The third point of comparison is spruce plantation in a clear-cut area, where the availability of resources (radiation, water and nutrients) is not limited by the overstory trees or the forest edge. The early growth rate of planted seedlings is supposedly the fastest in spite of the competition from ground vegetation and fast-growing pioneer species, which may be more severe than under canopy trees or in gaps.
Based on the previous results, it can be expected that the early height growth rate of established seedlings in small canopy gaps is rather low and it will take several years before height growth starts to recover. From the silvicultural point of view, the success of gap regeneration can be evaluated by comparing seedling height growth rate to that observed after other methods of regeneration, given that regeneration density is sufficient. Such comparisons are lacking, possibly due to the fact that seedling dynamics in harvested gaps appears to be poorly documented in the boreal region. There is more information available on the height growth of Norway spruce advance regeneration after complete or partial canopy removal, which has enabled comparisons with spruce plantations in mineral soil sites, for example (e.g., Valkonen 2000). Information from drained peatlands on the height development of spruce plantations has recently become available (Siipilehto et al. 2014).
The aims of this study were to quantify the early height growth rate of Norway spruce seedlings in small canopy gaps in a northern boreal spruce mire and the effect of gap size and different growing locations within the gap on growth by means of regression modelling. The observed height development was compared to that reported after different canopy removal cuttings from mineral soil sites and the average height development observed in peatland plantations. This study was based on measurements of annual shoot lengths of selected crop trees. The data originated from the same stand as that used in Hökkä et al. (2012).
2 Materials and methods
2.1 Study site
The study site was located in Tervola, northern Finland (N = 7341008, E = 440177) and represented a eutrophic, shallow-peated spruce swamp (Laine et al. 2012) with peat thickness varying from 10–50 cm. In terms of timber productivity, the site is comparable to a rich mineral soil site (Oxalis-Myrtillus site (Cajander 1926)) with average annual growth varying between 8–10 m3 ha–1 a–1 in southern Finland and 5–6 m3 ha–1 a–1 in northern Finland (Laine et al. 2012). The average annual temperature sum (with a 5 °C threshold) between 2000 and 2010 was 1076 dd °C (Venäläinen et al. 2005) and the altitude was 105 m above sea level. The site was drained for the first time in the 1960s, and complemented at 1980s. Currently the ditches are in satisfactory or poor shape with respect to their ability to transport water effectively. The present tree stand was composed of mature Norway spruce with a variable admixture of pubescent birch. The stand dominant height (mean height of 100 thickest trees on hectare) varied from 17 to 18 m, stem number from 735 to 2930, and stand volume from 170 to 227 m3 ha–1.
The experimental design was composed of a total of four randomised blocks, each including the three different gap sizes replicated two to four times, i.e., the experimental design was unbalanced. The diameter of the largest gap was 20 m, the middle-size gap 15 m, and the smallest gap 10 m. The total number of gaps was 33. The cutting of the gaps was performed in November 2004 when the soil was frozen and covered in snow. A more detailed description of the experimental design and cuttings can be found in Hökkä et al. (2011) and Hökkä et al. (2012).
2.2 Seedling survey and measurements
For the height growth analysis, a subsample of spruce seedlings was taken in spring 2013, after eight growing seasons had passed since the cutting, based on the same design of seedling survey plots as in Hökkä et al. (2011) and (2012). Seedlings were sampled from five circular survey plots, the largest of which was located at the centre of the gap (10 m2 in size) and four smaller (5 m2) ones at a distance of 1.5 m from the edge of the gap in each cardinal direction (Fig. 1). Only crop seedlings were selected, with the aim of achieving a maximum density of approximately 3000 seedlings ha–1 in each canopy gap. The selected crop seedlings had to be vigorous and healthy, were in a dominant position, located at least 0.6 m distance from each other, and were at least 20 cm tall at the time of the survey. To obtain the target density, a maximum of four seedlings were selected from the largest central survey plot and a maximum of two seedlings from each of the smaller plots, with the limitation that the maximum number of seedlings in a gap was nine, which would give 2997 ha–1 as the maximum crop seedling density. The majority of the sampled seedlings originated from the advance growth, i.e., they had been established before cutting, but part of the smallest seedlings had been established after cutting.
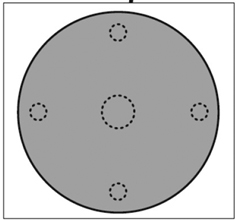
Fig. 1. Location of seedling survey plots in the canopy gaps.
From the selected seedlings the following characteristics were measured: distance and location from the survey plot centre to map seedlings exact position, seedlings total height, and annual height growth of the past five-year period with an accuracy of 1 cm. Not all selected seedlings included all five height growth observations, because some were only 20 cm tall at the time of data collection and had been established after cutting. The total number of crop seedlings in the data was 162 and their average density was 1809 ha–1 (1645 ha–1 if the three gaps in which no crop seedlings were found are included) (Table 1). In total, there were 716 height growth observations in the data.
| Table 1. Characteristics of the crop seedling data measured in spring 2013 from spruce mire canopy gaps. The gaps were cut in 2004. | ||||
| Mean | Min | Max | sd | |
| h | 64.7 | 20 | 270 | 43.5 |
| ih | 7.1 | 1 | 31 | 4.7 |
| N | 1809 a) | 333 | 2997 | 873 |
| h = seedling height, cm ih = seedling height growth, cm a–1 N = seedling density, ha–1 a) 1645 ha–1 if three gaps with no crop seedlings are included. | ||||
2.3 Statistical analyses
The height growth variation of crop seedlings was studied by means of growth model analysis. Eerikäinen et al. (2014) also used growth model analysis as the method to investigate factors affecting the height growth of spruce seedlings. Annual shoot length was used as the response variable. Since it was not normally distributed, logarithmic transformation was made after adding 0.5 to each growth observation. The data were hierarchically structured at block, gap, survey plot and seedling levels. Further, annual shoot lengths were sequentially correlated within each tree. To account for these correlations, the mixed linear model approach (e.g., Snijders and Bosker 2003) was applied in the analysis. The following model was used to explain height growth variations in the data (Snijders and Bosker 2003):
![]()
where
ihlijkt = height growth of tree k in year t in survey plot j in gap i in block l
β0 = model intercept
βh = fixed model parameters to be estimated
xhijkt = vector of explanatory variables measured at different levels of hierarchy (e.g., tree attributes, gap size, location, time since cutting)
fl = random effect of block l, NID (0, δf 2)
uli = random effect of gap i in block l, NID (0, δu2)
vlij = random effect of survey plot j in gap i in block l, NID (0, δv2)
mlijk = random effect of seedling k in survey plot j in gap i in block l, NID (0, δm2)
elijkt = random residual error of year t in tree k in survey plot j in gap i in block l, NID (0, δe2)
The covariance structure of the successive annual height growth observations was assumed to follow the first-order autoregressive (AR-1) structure. The model fixed parameters and variances of the random effects were estimated simultaneously using the maximum likelihood method as implemented in the mixed procedure of SAS (SAS Institute Inc. 2002–2008). The t-test was used to assess the significance of fixed parameters. Minimum 2xlog-likelihood and AIC were used to select the best model.
3 Results
3.1 Modelling height growth variation
The mean height growth of the crop seedlings during the five–year period from the third to the eighth year after cutting was rather low, only 7.1 cm year–1. There was an increasing trend from 2009 onwards (Fig. 2a). Improving height growth was also seen in the average height development over the study period (Fig. 2b).
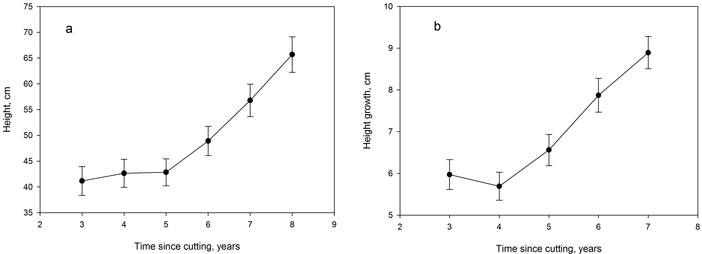
Fig. 2. Average observed height (a) and height growth (b) of spruce crop seedlings in the canopy gaps at different years after cutting (2004) calculated from the 2013 survey data. Vertical lines indicate the standard error of means.
In the model’s fixed part, the crop seedling’s height growth was linearly related to tree height at the beginning of each growing season (Table 2). There was a non-linear temporal trend comprising a negative linear term and a positive term with exponent 1.3. This corresponds to the average temporal trend seen in Fig 2a. In the largest gaps the negative first-order effect was significantly less than in the smaller gaps (Table 2).
| Table 2. Model for Norway spruce advance growth seedling annual height growth (ln(ih+0.5)) after cutting of canopy gaps in a drained spruce mire (see Eq. 1). | |||
| Variable | Estimate | Std. error | p-value |
| Intercept | 2.0717 | 0.3600 | <0.0001 |
| Height | 0.006656 | 0.000775 | <0.0001 |
| Time | –0.6391 | 0.2590 | 0.0139 |
| Time_20m | 0.04613 | 0.01340 | 0.0010 |
| Time13 | 0.3356 | 0.1227 | 0.0064 |
| 15m_C | 0.3491 | 0.1059 | 0.0017 |
| 15m_N | 0.2815 | 0.1340 | 0.0390 |
| 20m_S | –0.4110 | 0.1265 | 0.0017 |
| var(vij) | 0.02041 | 0.0119 | 0.0431 |
| var(mijk) | 0.03774 | 0.01350 | 0.0026 |
| AR1 | 0.2975 | 0.05173 | <0.0001 |
| var(eijkt) | 0.1324 | 0.009278 | <0.0001 |
| Mean bias | –0.84 cm | ||
| Height = seedling height at the beginning of growing season, cm Time = time since gap cutting, years Time_20 = dummy variable; for 20 m gap = 1, otherwise = 0 Time13 = (time)1.3 15m_C = dummy variable; for central location of 15 m gap = 1, otherwise = 0 15m_N = dummy variable; for northern location of 15 m gap = 1, otherwise = 0 20m_S = dummy variable; for southern location of 20 m gap = 1, otherwise = 0 | |||
There were also a few effects related to the gap size and seedling location within the gap that turned out to be significant in the model. On average, height growth was best in the largest (20 m diameter) gaps. The height growth of seedlings located in the middle or at the northern edge of the medium size gaps (15 m diameter) showed better growth than seedlings in other locations. Seedlings located in the southern part of the largest gap (20 m diameter) showed poorer growth than seedlings in other locations.
The block level variation related to the experimental design appeared to be non-significant and was omitted from the model. In addition, the random effect of the gap was non-significant. Most of the unexplained variation was at the residual level, i.e., among the annual shoot lengths (Table 2). The serial correlation coefficient was rather low, 0.2969.
3.2 Model application
The model shown in Table 2 was used to predict a height growth of 10 cm initial height seedlings from the beginning of the third post-harvest year to a point when breast height was achieved (Fig. 3). Based on height development predicted by the model, the seedlings reached 1.3 m in height when 11 or 12 growing seasons had passed since cutting, depending on the location of the seedling. In terms of height growth speed, two groups could be distinguished: seedlings located in the middle and north part of a 15 m gap and seedlings at a 20 m gap (excluding the southernmost edge) showed the best growth, while a lower growth rate was observed in other locations. There were also some minor differences in growth rates within the two groups. Predictions for any further heights should be inspected with caution, because there were very few trees taller than 1 m in the data (< 10%) and the power term for time since cutting may produce biased growth estimates for trees taller than 1.3 m.
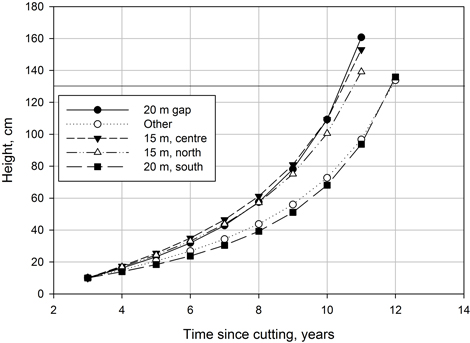
Fig. 3. Height development of spruce crop seedlings as a function of time since cutting in gaps of different sizes and in different locations within the gap according to the height growth model (Table 2). Other = all other gaps and locations except those shown by a separate curve. Initial seedling height in year 3 is 10 cm. Horizontal line indicates breast height.
3.3 Spruce seedling height growth after different canopy removal cuttings and in plantations
The observed spruce seedling mean height growth of this study was compared to results from earlier studies that reported on complete removal (release of advanced growth) and partial (thinning, shelter tree cutting, uneven-aged harvest) removal of Norway spruce overstory and to that observed in peatland Norway spruce plantations in the boreal region (Table 3). Different cutting treatments were compared separately to gap cutting. When possible, a similar study period and seedlings of a similar size (41 cm tall at the beginning of the third growing season after cutting in this study) were compared. Site types were mostly mineral soils sites and represented high or good productivity in all studies.
| Table 3. Description of height growth comparison studies. | ||||
| Study | Treatment | Location | Site type | Average annual height growth |
| This study | Gap cutting | Northern Finland | Herb-rich drained peatland site a) | 7.1 cm 5-year mean for 41 cm seedlings (3rd to 7th year after cut) |
| Cajander (1934) | Gap cutting | Southern Finland | Oxalis-Myrtillus mineral soil site b) | 8.8 cm for 41–60 cm seedlings (3rd to 7th year after cut) |
| Complete overstory removal | 21.1 cm for 41–60 cm seedlings (3rd to 7th year after cut) | |||
| Koistinen and Valkonen (1993) | Complete removal of overstory | Southern Finland | Oxalis-Myrtillus and Myrtillus mineral soil site b) | 20 cm (first 9 year mean) for 2.7 m tall seedlings |
| Örlander and Karlsson (2000) | Variable intensity removal of overstory by shelter tree cutting | Southern Sweden | Drained productive shallow-peated mineral soil site | 11–12 cm (first 8 year mean) for 20–50 cm seedlings with overstory density of 160 stems ha–1 or less |
| Nilson and Lundqvist (2001) | Variable intensity (30, 60, 85%) thinning of overstory from above or below | Northern and central Sweden | Vaccinium myrtillus low herb site and tall herb mineral soil site | 2–15 cm (first 7 year mean) for > 50cm seedlings depending on overstory density, time since cutting, and location |
| Eerikäinen et al. (2014) | Uneven-aged cutting | Southern Finland | Oxalis-myrtillus and Myrtillus mineral soil site | 1–4 cm (first 15 year mean) depending on tree size and time |
| Siipilehto et al. (2014) | Spruce plantation | Whole of Finland | Herb-rich, Vaccinium myrtillus and Vaccinium vitis-idaea drained peatland sites a) | 11.9 cm (first 9.2 year mean) c) |
| a) According to Laine et al. (2012) b) According to Cajander (1926) c) Calculated by dividing mean height (1.1 m) by mean age of plantations (9.2 years) in Siipilehto et al. (2014) data. | ||||
In canopy gaps cut in southern Finland, at the highly productive Oxalis-Myrtillus (OMT) mineral soil site, the annual height growth of the smallest (< 10 cm tall) seedlings remained under 10 cm in gaps of all sizes, while larger seedlings showed 20–50 cm height growth ten years after cutting in gaps larger than 0.04 ha in size (Cajander 1934). Calculated from the data published in the form of tables in Cajander (1934), the average height growth from year 3 to year 7 for seedlings of 41–60 cm in height in < 0.03 ha gaps was 8.8 cm year–1 (Table 3).
After complete overstory removal, the average annual height growth of spruce advance regeneration increased from less than 10 cm to 60 cm in ten years and varied according to initial seedling height (Cajander 1934). Average height growth calculated for 41–60 cm seedlings from year 3 to year 7 was 21.1 cm. Koistinen and Valkonen (1993) reported that the spruce advance regeneration average height growth after complete removal of overstory was 20 cm year–1 in the nine-year post-harvest period in OMT and Myrtillus types (MT) in southern Finland (Table 3).
Spruce advance regeneration height growth after shelter tree cutting has been investigated by Örlander and Karlsson (2000). For the first eight post-cutting years following partial or complete overstory removal height growth varied according to the number of retained canopy trees and size class of the seedlings. Mean annual height growth varied from 5 to 25 cm when shelter tree density was 160 ha–1 (12.2 m2ha–1) or less (Table 3). With this treatment the average eight-year growth of 20–50 cm seedlings was 11–12 cm year–1. In uncut forest or dense shelter tree stands (320 ha–1), only trees taller than 1.0 m showed an average annual growth clearly higher than 5 cm. The site was located in southern Sweden and represented a productive drained spruce peatland with a shallow peat layer and an estimated site index of H100 = 30 m.
In Nilson and Lundqvist’s (2001) study on partial (30%, 60% or 85%) removal of overstory by thinning either from above or below, spruce advance growth showed very slow mean annual height growth rates of 2–10 cm for saplings taller than 50 cm during the first six years after harvest in northern and central Sweden (Table 3). The sites were of good productivity (V. myrtillus – low herb type in the north, tall herb type in the central) but were located at a relatively high altitude (425 and 470 m.a.s.l.). Only in the seventh year did the maximum height growth reach 15 cm in the northern site. At the experimental site in central Sweden, the growth response was slightly stronger (seventh year maximum annual growth > 20 cm). Comparable mean growth cannot be calculated, but the visual inspection of growth for the period between the third and the seventh year after cutting suggest lower mean growth in the northern site and similar growth for the southern site (Nilson and Lundqvist 2001) than growth in this study.
Eerikäinen et al.’s (2014) results on spruce height growth after uneven-aged harvest in mineral soil experimental sites in southern Finland also indicated very slow rates of growth. During a 15-year period after cutting for a medium growth forest site, the annual height growth varied from 1 to 4 cm (Table 3). Lundqvist (1989) has reported similar results with very slow annual height growth (2–4 cm) after cutting an uneven-aged forest in Sweden.
Siipilehto et al. (2014) reported average heights of spruce plantations established in peatland site clear-cuts based on a large sample plot data obtained from practical forest regeneration areas. The results are valid for a wide geographical area in Finland and the mean represent conditions prevailing in central Finland. At the average age of 9.2 years, the mean height of planted spruces was 1.1 m, which corresponds to an average annual height growth of 11.9 cm after planting (Table 3).
4 Discussion
Based on the results of this study, the growth rate of Norway spruce seedlings from the third to the eighth year after canopy gap cutting in a drained spruce mire stand in northern Finland was fairly low. Cajander’s (1934) results on height growth for a comparable post-harvest period for the same sized advance growth seedlings growing in gaps of a similar size (< 0.03 ha) were about 24% higher (8.8 cm) than the average 7.1 cm in this study. The difference may be explained by the fact that Cajander’s (1934) material was from southern Finland and this study was conducted in northern Finland. In a similar forest site type, a 25–30% difference in timber productivity can be due to climate (Gustavsen 1980), which is also reflected in the seedlings’ height growth.
When compared to studies on seedling growth recovery after complete overstory removal, the mean height growth in this study was much less than the average of 20 cm reported by Koistinen and Valkonen (1993) for a nine-year period after release cutting on mineral soil sites in southern Finland. The growth difference is caused by a combination of several reasons: i) the total removal of overstory trees provides clearly better growing conditions in comparison to those prevailing in canopy gaps; ii) the geographical difference between southern and northern study sites; and ii) greater mean height (2.7 m) of the seedlings at the time of cutting in the data from Koistinen and Valkonen (1993). On the basis of the comparison made by Valkonen (2000), the height growth rate after complete overstory removal in Cajander’s (1934) data was even faster than that reported by Koistinen and Valkonen (1993), perhaps because in the former study the oldest stunted seedlings were excluded from the analysis (Cajander 1934).
Partial overstorey removal studies have been conducted in Sweden. Örlander and Karlsson (2000) study on variable intensity shelter tree cutting in southern Sweden showed that a significant growth response of seedlings can be found if the density of retained trees was 160 stems ha–1 or less. Better growth than in this study may be explained by the southern location since the site types are quite similar. In addition, the average overstory density of the shelter tree stand was probably lower than the average in this study although overstory density in gaps was not defined. Nilson and Lundqvist’s (2001) results on spruce seedling growth rates after partial overstory removal were somewhat lower than or similar to those observed in this study in the northern study site, but slightly better in the central Sweden study site. In the heaviest canopy removal treatment, only 20–50 m3ha–1 standing volume was retained but even in those stands the growth response was not remarkable. Since the seedlings were also larger than in this study, the poorer average growth may be attributable to more unfavourable growing conditions in the high altitude forests in the Swedish study sites.
Comparisons with uneven-aged cuttings indicated clear differences in the height growth of advance regeneration in the post-harvest period. In this study the average growth was two to three times better than in uneven-aged harvest stands observed in southern Finland by Eerikäinen et al. (2014) or in Sweden (Lundqvist 1989). Based on simulations with the model constructed in this study, it would take 11–12 years for a 10 cm-tall spruce seedling to reach a height of 1.3 m. If an assumption is made that for a spruce seedling it takes three to five years to reach 10 cm (Hökkä et al. 2011), breast height can be reached in 20 years or less in the gaps of this study site. That would be significantly quicker when compared to the average of 60 years in an uneven-aged stand in southern Finland mineral soil sites (Eerikäinen et al. 2014), or 24–47 years as reported by Lundqvist (1989) from mineral soil sites in Sweden. One possible reason may be that after uneven-aged cutting in the more southerly located mineral soils sites, the retained stand remains rather stocked (Eerikäinen et al. 2014) and conditions more shady than after cutting canopy gaps in naturally more open (e.g., Heikurainen 1971; Norokorpi et al. 1997) spruce mire stands in northern Finland. Heterogeneous stand structures in drained peatland spruce mires persist for decades after drainage (Sarkkola et al. 2003). Such conditions allow more light to penetrate to the soil surface and advance growth may be capable to respond more quickly to increased light availability.
In peatland spruce plantations (Siipilehto et al. 2014), seedling average height growth was 11.9 cm which was almost 70% higher than the average growth in this study. The difference may partly be due to the slight climatic differences between the plantation data and these data, but it is mostly due to different growth patterns of planted and advance growth seedlings (Valkonen 2000). According to Saksa (2011), planted spruce seedlings in mineral soil sites in central Finland grow height 10–11 cm annually during the first four years after planting on average, and after that growth will further increase. Advance growth seedlings’ full growth recovery may take 10–15 years after the removal of the canopy trees (Valkonen 2000). Valkonen (2000) concluded that advance growth seedlings which are 1.0–1.5 m tall at the time of canopy removal will achieve their inherent growth rate in 10 years and after that their growth is comparable to that of planted seedlings.
The height growth rate observed in this study is in line with earlier results obtained in canopy gaps by Cajander (1934). After release cutting in mineral soil sites (Cajander 1934; Koistinen and Valkonen 1993) growth was clearly better than in this study, because of the lack of competing canopy trees and partly due to the southern location and taller seedlings (Koistinen and Valkonen 1993). In partial overstory removal studies, growth was better (Örlander and Karlson 2000) or slightly poorer (Nilson and Lunqvist 2001) than that observed in this study, and in uneven-aged cuttings (Eerikäinen et al. 2014; Lundqvist 1989) much lower growth rates than in this study were observed. The seedlings’ growing environment in this study’s data may represent conditions where the change in light availability is something between complete removal of the overstory and partial cutting that is typical for uneven-aged harvest or shelter tree cutting. The observed growth differences that cannot be attributed to geographical or site deviations may reflect this difference.
The non-linear temporal trend in the annual height growth indicated a slight improvement in height growth after the fourth growing season. This corresponds well with the conclusions of Koistinen and Valkonen (1993) and Valkonen (2000) that Norway spruce height growth does not recover before the fourth or fifth growing season following overstory removal. A similar lag of three to four years in growth after release cutting was also reported by Örlander and Karlsson (2000) in a southern Swedish site as well as Nilson and Lundqvist (2001) in central and northern Sweden, and Metslaid et al. (2005) in Estonia. According to Valkonen (2000) it may take another five years or more before seedling growth rates are fully recovered and comparable to that of planted seedlings. Longer recovery periods have also been observed. Skoglefald (1967) reported a seven-year recovery time for Norway spruce in northern Norway and Groot and Hökkä (2000) found black spruce (Picea mariana (Mill.) BSP) advance regeneration height growth suppression effects to last 21 years. This lag in growth is due to the fact that suppressed seedlings need several years for their needles and root system to adapt and for resources to be reallocated to changed light conditions (Kneeshaw et al. 2002).
Kneeshaw et al. (2002) showed that the height growth of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) advance growth decreased in the first year after partial overstory removal, followed by an increasing growth trend in the following years. Because decreasing height growth several years after overstory removal has not been previously reported, the height growth minimum in the fifth year (2009) of this study may be due to the cool and wet summer of 2008. Norway spruce height growth has been observed to be poorly correlated with climatic variables, but it is likely that the previous year’s weather impacts on the growth of the coming year through the prevailing weather conditions at the time of bud formation (Levanic et al. 2009). For black spruce (Picea mariana (Mill.) BSP), the height growth of Arctic timberline trees was more strongly correlated with the weather of the previous year than that of the current year (Gamache and Payette 2004). With these data it was impossible to assess the impact of annual weather conditions in detail, because the year effect was mixed with the time effect.
The most important variable explaining variation in growth was tree height, i.e., better height growth was related to taller seedlings. This result is similar to that observed by Cajander (1934), Koistinen and Valkonen (1993), Örlander and Karlsson (2000) and Eerikäinen et al. (2014). Örlander and Karlsson (2000) explained the poor growth of the smallest seedlings by their superficial root system and lack of water in the topmost soil layer after a significant change in heat radiation reaching the ground after release cutting. In these data, it is not likely that the lack of water is a problem due to the nature of the site and relatively poor drainage conditions, despite ditches. The rise in the ground water table level and increased soil moisture is expected after harvest in drained spruce mire stands (Lundin 2000). Because of that, small seedlings may benefit from the moist soil surface conditions in peatland sites in terms of survival and growth.
As hypothesised, gap size had an influence on the growth of the seedlings. Height growth in the 20 m diameter gap was significantly better than in the smaller gaps. This is rather logical because in the largest gap most light is available for seedlings. Comparable growth rates were also observed at the middle and northern edge of the 15 m gap. Chantal et al. (2003) showed that in a 50 m diameter canopy gap in southern Finland the highest amount of radiation was observed at several metres north of the centre of the gap and the central location and northern edge are equal in terms of light availability. The asymmetry of light distribution is supposedly more pronounced in the site of this study because of the significantly lower sun angle due to the northern latitude. On the other hand, the surrounding forest is shorter (17–18 m). Good growth in middle and northern part of the 15 m gap may be related to the combined effects of light availability – which is best in the largest gaps – but also to differences in competition conditions. According to Hökkä et al. (2011), birch seedlings were most abundant in the 20 m gaps with a significant difference compared to 15 m and 10 m gaps (also Roy et al. 2000). It is therefore possible that in the 15 m gap competition from birch is less intense and in the middle and the northern edge there is also enough light available for good growth. Based on simulations, the growth rates in these locations were close to each other (Fig. 3). The fact that in the southern edge of 20 m gap growth rate corresponded to that of the rest of the data may be partly explained by the combined effect of the shady location even after cutting, and the higher competition from birch seedlings. Competition from birch or ground vegetation was not directly measured in these data, but is based on number of birch seedlings given in Hökkä et al. (2011).
Since the gaps were rather small on average, root competition from larger trees growing at the edge of the surrounding forest also extends to the gap area. In the 10 m gap in particular, the root systems of the trees growing outside the gap may cover a significant proportion of the rooting space in the gap. Despite of that, seedling growth in the 10 m gap was not much poorer than in the larger gaps (Fig. 3). This may partly reflect the openness of the canopy in the northern peatland spruce stands (Heikurainen 1971; Sarkkola et al. 2003) and possibly suggest that the effect of a small gap – created by removal of a couple of trees – on the sub-canopy light conditions may be proportionally greater than the actual size of the gap.
Variation in seedling height was rather narrow, i.e., > 90% of seedlings were between 0.2 m and 1.0 m in height at the time of the survey in spring 2013. Because of that the model developed here should not be applied to trees taller than 1.3 m, and more data should be collected to develop a more widely applicable height growth model. Generalisation of the results is also limited by the fact that the data were from only one study location. Nevertheless, the rather large number of canopy gaps, 33 in total, and the result that neither the blocks nor the gaps showed significant deviations from the modelled height growth, suggest that the captured pattern was rather uniform.
The conclusions that can be made from the present study are that it will take four to five years for the growth Norway spruce seedlings’ growth rate to start recover after cutting of canopy gaps. Growth is linearly related to tree height, being highest for the tallest seedlings. Seedlings in the largest gaps and in some locations in the mid-size gap showed the best average growth.
In their review, Coates and Burton (1997) concluded that gap dynamics is common in conditions where large-scale high-intensive disturbances are rare, e.g., in high-elevation forests and low-elevation interior sub-boreal spruce stands that have escaped fire for a long time. Moist northern boreal peatland spruce forests are likely such an environment where high-intensive disturbances are rare and gap dynamics is a natural way of regeneration. The results on post-harvest height growth of spruce advance regeneration obtained in this study suggest that regeneration in canopy gaps does not last for decades. Seedling height growth in gaps was significantly lower than in plantations but comparable or slightly lower than in partially harvested stands and clearly better than in uneven-aged stands. To obtain a better understanding of the height growth rate and height development over time, monitoring should continue for at least another five years. Further, the competition factors (density and height of birch regeneration and that of ground vegetation) should also be assessed in some quantitative way to understand whether they have a significant role in the seedlings’ early height growth process.
Acknowledgments
The field experiment presented in the study was established in co-operation with Finnish Forest Research Institute and Metsähallitus. The comments of two anonymous referees greatly improved the manuscript.
References
Bergan J. (1971). Skjermforyngelse av gran samenlignet med plantning i Grane i Nordlund. Summary: Natural Norway spruce regeneration under shelterwood compared with plantations at Grane in Nordland. Meddelser fra det Norske Skogforsöksvesen 28(104): 194–211. [In Norwegian with English summary].
Cajander A.K. (1926). The theory of forest types. Acta Forestalia Fennica 29(3). 108 p.
Cajander E.K. (1934). Kuusen taimistojen vapauttamisen jälkeisestä pituuskasvusta. Über den Höhenzuwachs der Fichtenpflanzenbestände nach der Befreiung. Communicationes Instituti Forestalis Fenniae 19(5): 1–53. [In Finnish with German summary].
Chantal M., Leinonen K. Kuuluvainen T., Cescatti A. (2003). Early response of Pinus sylvestris and Picea abies seedlings to an experimental canopy gap in boreal spruce forest. Forest Ecology and Management 176: 321–336. http://dx.doi.org/10.1016/S0378-1127(02)00273-6.
Coates K.D. (2000). Conifer seedling response to northern temperate forest gaps. Forest Ecology and Management 127: 249–269. http://dx.doi.org/10.1016/S0378-1127(99)00135-8.
Coates K.D., Burton P.J. (1997). A gap-based approach for development of silvicultural systems to address ecosystem management objectives. Forest Ecology and Management 99: 337–354. http://dx.doi.org/10.1016/S0378-1127(97)00113-8.
Drobyshev I., Nihlgård B. (2000). Growth response of spruce saplings in relation to climatic conditions along a gradient of gap size. Canadian Journal of Forest Research 30: 930–938. http://dx.doi.org/10.1139/x00-008.
Eerikäinen K., Valkonen S., Saksa T. (2014). Ingrowth, survival and height growth of small trees in uneven-aged Picea abies stands in southern Finland. Forest Ecosystems 2014(1:5). 10 p. http://dx.doi.org/10.1186/2197-5620-1-5.
Gamache I., Payette S. (2004). Height growth response of tree line black spruce to recent climate warming across the forest-tundra of eastern Canada. Journal of Ecology 92: 835–845. http://dx.doi.org/10.1111/j.0022-0477.2004.00913.x.
Groot A., Hökkä H. (2000). Persistence of suppression effects on black spruce advance regeneration after overstory removal. Canadian Journal of Forest Research 30(5): 753–760. http://dx.doi.org/10.1139/x00-007.
Gustavsen H.G. (1980). Talousmetsien kasvupaikkaluokittelu valtapituuden avulla. Summary: Site index curves for conifer stands in Finland. Folia Forestalia 454. 31 p. [In Finnish with English summary].
Hånell B. (1992). Skogsfönyelse på högproduktiva torvmarker – plantering av gran på kalhygge och under skärmträd. Forest renewal on productive peatlands: Planting of Norway spruce on clear cuts and in shelterwoods. Swedish University of Agricultural Sciences, Department of Silviculture Report 34. Umeå 1992. 71 p. [In Swedish with English summary].
Hånell B. (1993). Regeneration of Picea abies forests on highly productive peatlands – clearcutting or selective cutting? Scandinavian Journal of Forest Research 8(1–4): 518–527. http://dx.doi.org/10.1080/02827589309382798.
Hanssen K.H. (2003). Natural regeneration of Picea abies on small clear-cuts in SE Norway. Forest Ecology and Management 180: 199–213. http://dx.doi.org/10.1016/S0378-1127(02)00610-2.
Heikurainen L. (1971). Virgin peatland forests in Finland. Acta Agralia Fennica 123: 11–26.
Hökkä H., Repola J., Moilanen M. Saarinen M. (2011). Seedling survival and establishment in small canopy openings in drained spruce mires in Northern Finland. Silva Fennica 45(4): 633–645. http://dx.doi.org/10.14214/sf.97.
Hökkä H., Repola J., Moilanen M. Saarinen M. (2012). Seedling establishment on small cutting areas with or without site preparation in a drained spruce mire – a case study in northern Finland. Silva Fennica 46(5): 695–705. http://dx.doi.org/10.14214/sf.920.
Hökkä H., Repola J. (2012). Five-year growth of Norway spruce advance regeneration following cutting of small canopy openings in a spruce mire. In: Proceedings of the 14th International Peat congress, Stockholm. International Peat Society. p. 163–168.
Kneeshaw D., Williams H., Nikinmaa E., Messier C. (2002). Patterns of above- and below-ground response of understory conifer release 6 years after partial cutting. Canadian Journal of Forest Research 32: 255–265. http://dx.doi.org/10.1139/x01-190.
Koistinen E., Valkonen S. (1993). Models for height development of Norway spruce and Scots pine advance growth after release cutting in southern Finland. Silva Fennica 27(3): 179–194. http://doi.org/10.14214/sf.a15671.
Kuuluvainen T. (1993). Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: a review. Annales Zoologi Fennici 31: 35–51.
Laine J., Vasander H., Hotanen J.-P., Nousiainen H., Saarinen M., Penttilä T. (2012). Suotyypit ja turvekankaat – opas kasvupaikkojen tunnistamiseen. Metsäkustannus Oy. ISBN 978-952-5694-89-5. 160 p. [In Finnish].
Leemans R. (1991). Canopy gaps and establishment patterns of spruce (Picea abies (L.) Karst.) in two old-growth coniferous forests in central Sweden. Vegetatio 93: 157–165. http://dx.doi.org/10.1007/BF00033209.
Levanic T., Gricar J., Gagen M., Jalkanen R., Loader N., McCarroll D., Oven P., Robertson I. (2009). The climate sensitivity of Norway spruce (Picea abies (L.) Karst.) in the southern European Alps. Trees – Structure and Function 23(1): 169–180. http://dx.doi.org/10.1007/s00468-008-0265-0.
Lundin L. (2000). Water environment care at peatland forestry practices. In: Rochefort L., Daigle J.-Y. (eds.). Sustaining our peatlands. Proceedings of the 11th International Peat Congress, Volume II. Canadian Society of Peat and Peatlands and International Peat Society. p. 952–961.
Lundqvist L. (1989). Blädning i ganskog – strukturförändring, volymtillväxt, inväxning och föryngring på försöksytor skötta med stamvis blädning. Swedish University of Agricultural Sciences, Institute of Silviculture, Umeå. 22 p. [In Swedish].
Metslaid M., Ilisson T., Nikinmaa E., Kusmin J., Jögiste K. (2005). Recovery of advance regeneration after disturbances: acclimation of needle characteristics in Picea abies. Scandinavian Journal of Forest Research 20(6): 112–121. http://dx.doi.org/10.1080/14004080510043352.
Moilanen M., Ferm A., Issakainen J. (1995). Kuusen- ja koivuntaimien alkukehitys korven uudistamisaloilla. Metsätieteen aikakauskirja – Folia Forestalia 1995(2): 115–130. [In Finnish].
Nieminen M. (2004). Export of dissolved organic carbon, nitrogen and phosphorus following clear-cutting of three Norway spruce forests growing on drained peatlands in southern Finland. Silva Fennica 38(2): 123–132. http://dx.doi.org/10.14214/sf.422.
Nilson K., Lundqvist L. (2001). Effect of stand structure and density on development of natural regeneration in two Picea abies stands in Sweden. Scandinavian Journal of Forest Research 16: 253–259. http://dx.doi.org/10.1080/713785124.
Norokorpi Y., Lähde E., Laiho O. Saksa T. (1997). Stand structure, dynamics, and diversity of virgin forests on northern peatlands. In: Trettin C.C., Jurgensen M.F., Grigal D.F., Gale M.R., Jeglum J.K. (eds.). Northern Forested Wetlands: Ecology and Management. CRC Press Inc.; Lewis Publishers. p. 73–87. ISBN 1-56670-177-5.
Örlander G., Karlsson C. (2000). Influence of shelterwood density on survival and height increment of Picea abies advance growth. Scandinavian Journal of Forest Research 15: 20–29. http://dx.doi.org/10.1080/02827580050160439.
Page L.M., Cameron A.D. (2006). Regeneration dynamics of Sitka spruce in artificially created forest gaps. Forest Ecology and Management 221: 260–266. http://dx.doi.org/10.1016/j.foreco.2005.10.006.
Roy V., Ruel J.-C., Plamondon A. (2000). Establishment, growth and survival of natural regeneration after clearcutting and drainage on forested wetlands. Forest Ecology and Management 129: 253–267. http://dx.doi.org/10.1016/S0378-1127(99)00170-X.
Saksa T. (2011). Kuusen istutustaimien menestyminen ja tukkimiehentäin tuhot eri tavoin muokatuilla uudistusaloilla. Metsätieteen aikakauskirja 2/2001: 91–105. [In Finnish]. http://www.metla.fi/aikakauskirja/full/ff11/ff112091.pdf.
Sarkkola S., Alenius V., Hökkä H., Laiho R., Päivänen J., Penttilä T. (2003). Changes in structural inequality in Norway spruce stands on peatland sites after water-level drawdown. Canadian Journal of Forest Research 33(2): 222–231. http://dx.doi.org/10.1139/x02-179.
SAS Institute Inc. 2003–2008. SAS OnlineDoc version 9.1 (computer program). SAS Institute Inc., Cary, N.C. https://support.sas.com/learn/statlibrary/.
Siipilehto J., Saarinen M., Hökkä H. (2014). Taimikoiden pituuskehityksen luotettavuus sovellettaessa MOTTI-ohjelmiston kangasmaiden ennustemalleja turvekankaille. Metlan työraportteja /Working Papers of the Finnish Forest Research Institute 294. 25 p. [In Finnish]. http://www.metla.fi/julkaisut/workingpapers/2014/mwp294.htm.
Skoglefald S. (1967). Fristilling av naturlig gjenvekst av gran. Summary: Release of natural Norway spruce regeneration. Meddelser fra det Norske Skogsforsöksvesen 23(8): 385–409. [In Norwegian with English summary].
Snijders T., Bosker R. (2003). Multilevel analysis. An introduction to basic and advanced multilevel modeling. Sage Publications Ltd. 266 p.
Valkonen S. (2000). Kuusen taimikon kasvattamisen vaihtoehdot Etelä-Suomen kivennäismailla: Puhdas kuusen viljelytaimikko, vapautettu alikasvos ja kuusi-koivusekataimikko. Metsäntutkimuslaitoksen tiedonantoja 763. 83 p. + 4 osajulkaisua. ISBN 951-40-1721-8. [In Finnish].
Valkonen S., Koskinen K., Mäkinen J., Vanha-Majamaa I. (2011). Natural regeneration in patch clear-cutting in Picea abies stands in Southern Finland. Scandinavian Journal of Forest Research 26: 530–542. http://dx.doi.org/10.1080/02827581.2011.611818.
Venäläinen A., Tuomenvirta H., Pirinen P., Drebs A. (2005). A basic Finnish climate data set 1961–2000 – description and illustrations. Finnish Meteorological Institute, Reports 2005:5. 27 p. ISBN 951-697-615-8.
Yamamoto S.-I. (2000). Forest gap dynamics and tree regeneration. Invited review. Journal of Forest Research 5: 223–229. http://dx.doi.org/10.1007/BF02767114.
Total of 44 references
