Jaana Luoranen 
Autumn versus spring planting: the initiation of root growth and subsequent field performance of Scots pine and Norway spruce seedlings
Luoranen J. (2018). Autumn versus spring planting: the initiation of root growth and subsequent field performance of Scots pine and Norway spruce seedlings. Silva Fennica vol. 52 no. 2 article id 7813. https://doi.org/10.14214/sf.7813
Highlights
- Conifer seedlings planted after mid-September generally have poor rooting, which causes poor root egress during the following spring
- Although Scots pine and Norway spruce seedlings planted in late autumn may have a slightly reduced growth, it is possible to plant them if weather conditions are favorable in late-autumn, without increased mortality.
Abstract
There is a need to extend the planting season of conifer regeneration into periods where the soil remains unfrozen due to a lack of available labor and the mechanization of planting. This study investigated how the summer- (August) and autumn-, especially late autumn (mid-September to mid-October) plantings affect the field performance of Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karst.) container seedlings. This study examined the timing of root growth just after planting, shoot flush and the start of root growth the following spring, and subsequent field performance. Seedlings of both species were planted in a nursery field trial, and in a clearcut reforestation site from August to October and the following May. The root growth of planted seedlings declined in September and ceased after mid-September. In the following spring, seedlings which were planted in early-autumn started their root growth faster than late-autumn-planted seedlings in both species. There was no difference in the timing of shoot flush for various planting dates. During the initial two years after planting, the shoot growth of spring-planted seedlings was lower, compared to autumn-planted seedlings. In conclusion, it is possible to plant conifer seedlings in the boreal forest zone up to October under non-limiting field conditions.
Keywords
bud burst;
planting date;
root growth;
shoot growth
-
Luoranen,
Natural Resources Institute Finland (Luke), Production systems, FI-77600 Suonenjoki, Finland
E-mail
jaana.luoranen@luke.fi
Received 20 September 2017 Accepted 15 March 2018 Published 19 March 2018
Views 81490
Available at https://doi.org/10.14214/sf.7813 | Download PDF

1 Introduction
Conventionally, conifer seedlings are planted in spring from May through early June. This short period is busy, both in the nurseries and forests. In addition, the number of forest laborers is decreasing. The mechanization of planting work is one solution to the above mentioned problems (Nilsson et al. 2010). For example, Norway spruce (Picea abies (L.) Karst.) seedlings are suggested to plant from May to the end of September (Luoranen et al. 2005, 2006) and Scots pine (Pinus sylvestris L.) seedlings in May and late August – early September in Finland (Luoranen and Rikala 2013). On the other hand, in western Canada, the planting season is recommended to end in the middle of August (Grossnickle and Folk 2003; Tan 2007). For the purpose of machine planting, these periods are too short. To be cost-efficient, the work period of a machine should be a whole period with unfrozen soil.
There are several studies in which planting dates in container conifer seedlings has been studied (Valtanen et al. 1986; Kinnunen 1989; Luoranen et al. 2006). In these studies, however, the planting season ended in mid-September. In the studies of Luoranen and Rikala (2013) with Scots pine and Wallertz et al. (2016) with Norway spruce, the planting season was extended in October and November. Despite these studies, we do not know all the after-effects of late autumn plantings.
Several studies have shown that new root growth is important for seedling survival (Grossnickle 2005 and reference therein). Seedlings should be planted at that point in their cycle when they have the ability to grow roots to ensure the best chance of field performance (Grossnickle 2005 and reference therein). From earlier studies, we know that root growth in many tree species grown in the areas with low winter temperatures cease in late autumn (Lyr and Hoffmann 1967). For example, the results of Wallertz et al. (2016) showed that root growth ceased in Norway spruce seedlings when they were planted in October in the southern part of Sweden and Norway. The study of Wallertz et al. (2016) was undertaken in a more southern zone with warmer autumns and milder winters, compared to climatic conditions in Finland. Root growth probably ceases earlier in northern conditions and affects the field performance of conifer seedlings more. Before a recommendation to plant in late-autumn, we have to know better how much poor rooting in late autumn affects the field performance of seedlings.
The planting date has been observed to have some after-effects in the following year. For example, the earlier flush of summer planted seedlings in the following spring (Luoranen and Rikala 2015), the reduced growth of autumn planted seedlings (Luoranen and Rikala 2013; Wallerz et al. 2016) and the reduced ability to recover from damage (Wallerz et al. 2016). It is not known the reason for these after-effects. Luoranen et al. (2006) speculated that well-rooted, summer-planted seedlings may commence root growth earlier than newly planted seedlings in spring, which can then affect the later growth of seedlings. October-planted seedlings do not grow roots after planting (Wallertz et al. 2016), and are probably comparable to spring-plantings. Unrooted seedlings are also more susceptible to stresses (Grossnickle 2005). These factors may then affect the root growth of autumn-planted seedlings in the following spring. Unrooted seedlings may also have an increased risk of spring drought. All these factors can affect subsequent shoot growth of the seedlings.
The aim of the study was to investigate the field performance of late summer- and autumn-plantings in Norway spruce and Scots pine seedlings, in comparison to spring planting. Trials were established, both in a former nursery field and in a regeneration site with higher risks of damage. My hypothesis were that i) root growth after planting is minimal in both tree species at the end of September and in October, and ii) well-rooted, i.e. late-summer-planted seedlings, start their shoot and root growth earlier in the following spring than late-autumn planted seedlings. The hypothesis was also that poor rooting iii) increases the seedlings risks of damage in winter and spring, as well as reducing the seedlings’ ability to recover from damage, and iv) reduces seedling growth over the following years, and that v) these post-planting effects are stronger in Scots pine seedlings than in Norway spruce.
2 Material and methods
2.1 Seedling material
Two-year-old Norway spruce (a mean height of 22 ± 3 cm) and one-year-old Scots pine (15 ± 2 cm) seedlings were used. Seedlings were grown in hard plastic Plantek (BCC, Iso-Vimma, Finland) 81F trays (81 cells per tray, 546 cells m–2, cell volume 85 cm3) at Suonenjoki Research Nursery of Natural Resources Institute Finland (62°39´N, 27°03´E, with an of altitude 142 m a.s.l.). Seeds of Norway spruce from seed orchard no. 374 (supplying Central Finland) were sown on the 15th of June in 2013. Norway spruce seedlings were grown in a greenhouse until October 2013. Then they were transferred to an outdoor area where they were during the winter and the following growing season. In mid-winter, seedlings were under snow cover. In the second growing season, the Norway spruce seedlings were given a commercial fertilizer solution (0.1% Kekkilä Forest Superex with 22N:5P:6K + micronutrients, Kekkilä Co., Tuusula, Finland) 16 times from 19th May to 29th August 2014 and they received a total of 102.5 g m–2 fertilizer solution.
Seeds of Scots pine from seed orchard no. 339 (supplying Central Finland) were sown on 6th of June 2014. The Scots pine seedlings were in a greenhouse until 11th of July 2014. Then they were transferred to the same outdoor growing area in which the Norway spruce seedlings were. Scots pine seedlings were fertilized eight times from 4th June to 14th August 2014 and they received a total of 47.0 g m–2 of 0.1% Kekkilä Forest Superex. Tilt (propiconatsol 250 g L–1) was applied to all Scots pine seedlings for protection against pathogens on the 11th and 30th of July 2014, and to the spring planted seedlings on the 18th of November 2014.
In both species, seedlings were irrigated 2–4 times per week, depending on the weather conditions. For all seedlings in each species, three-week short day (SD) treatment (14-hour nights and 10-hour days) was started on the 21st of July 2014 with the use of a blackout curtain in the outdoor growing compound. Fertilization was also continued during SD-treatment. A day before planting, the moisture of peat plugs was checked and, if necessary, watered. For planting, seedlings were taken directly from the growing compound just before planting (planted within hours) without any package in 2014. Seedlings intended for spring-planting were packed in cardboard boxes at the end of October and kept in a freezer storage (–3 °C) until the end of April 2015. After that, seedlings were taken from the boxes and kept with watering in the outdoor growing area until planting. All seedlings were treated with KarateZeon Technique (lambda-cyhalotrin 100 g L–1) against pine weevil on 15th of August 2014 and on 21st of April 2015 (spring-planted seedlings).
2.2 Planting trials
One planting trial was established at a former nursery field in Suonenjoki research nursery to follow intensively the phenology and growth of seedlings. The soil at the former nursery field was fine sand containing only few organic matter and its nutrient level was low. The nursery field was not fully equivalent to a real situation in the mounded regeneration sites, and seedlings were also planted in a reforestation site about 20 km from Suonenjoki. In order to get the nursery field closer to the real regeneration site conditions in the forest, it was prepared and fertilized with Varhaisviljelyn Y 2 (600 kg ha–1; N-P-K 12-4-13.3, B 0.15%; YaraSuomi Oy, Finland) before establishment. To reduce the competition of grasses and seed sown birches, field vegetation was cut with a lawnmower in the summer 2015.
The regeneration site (hereafter referred to as the forest site) was clear cut in July 2014 and spot mounded with an excavator in early August 2014. In the spot mounds, the mineral soil layer was on the top of a double humus layer. The soil was classified as a medium coarse soil with disintegrated stones and rocks. Pine and spruce seedlings were planted in separate trials. The trial with spruce was established to the lower part of hillside, and the other one with pine to the upper part with more coarse soil. Fireweed (Chamerion angustifolium (L.) Scop) and raspberry (Rubus idaeus L.) were the main ground vegetation species in regeneration sites.
For all trials, seedlings were planted on the same dates, on the 19th of August, 1st, 15th and the 29th of September, 13th of October 2014 and on 19th of May 2015. In the nursery field, seedlings of both species were planted in rows with 1 m between seedlings in a row and between rows. At each planting date (D) and tree species (S), 45 seedlings in nine blocks (five seedlings in each block) using a split-plot design (planting date as a main plot and tree species as a subplot) were planted. For root growth measurements, three additional seedlings of both species were planted between seedlings in each plot on each planting date and block in the nursery field.
Tree species differ in their optimal growing conditions: pine seedlings normally grow on coarser soil types than spruce. Thus, the tree species were in separate trials in the forest site. For pine, there were 43 groups (blocks) of mounds, each group containing six mounds that were selected to be as similar as possible, nearby each other. Planting dates were randomized to these six mounds within a block. For spruce, the experimental design was similar, but with 45 groups of mounds. In all trials, seedlings were planted to a depth of 6 cm (the length of the stem under the soil surface). In the forest, seedlings were planted to the middle of mounds and at least 10 cm from nearest humus edge.
The quality of the mounds was determined at the forest site to assure that similar kinds of mounds within blocks for different planting dates were selected. The surface material of the mounds was classified as mineral, mineral-humus, and humus, and the soil texture of mound surface material was a normal mineral soil, with a mixture of mineral and gravel (mostly mineral) and gravel components. Most (86%, varying from 56% to 93% among planting dates) of the Norway spruce seedlings were planted in mounds covered by mineral soil and 14% (7–24%) were covered by a mixture of mineral soil and humus. Mineral soil on mounds contained gravel in 2% (0–9%) of cases. Mineral soil was a fine sandy soil in 58% (42–89%) with a mixture of fine sand and gravel in 40% (11–58%) of the mounds. Although we tried to select similar mounds and randomized to the different planting dates, more mounds with a mixture of gravel and fine sandy soil were in August- and May-plantings.
In the Scots pine, 86% (70–98%) of mounds were covered by mineral soil and 13% (2–30%) were a mixture of humus and mineral soil. The proportion of mounds with humus cover was only 1% (0–2%). The soil on mounds was gravel in 8% (0–40%) of cases, with the highest proportion in the May planting. Pure sandy soil covering mounds was on average 26% (12–37%) of the mounds and most (66%; 23–84%) of the mounds were a mixture of sandy soil and gravel.
2.3 Seedling measurements
At planting and at the end of each growing season, the seedling height was measured from the ground surface to the top of the terminal buds (accuracy 0.5 cm) and the diameter (accuracy 0.1 mm) was measured at about 2 cm above the ground or mound surface. The leader growth (height growth in a measuring year) was measured, as well as the cause (drought, frost, pine weevil, black spruce beetle, vole, frost heaving, field vegetation, etc.) and degree of damage (healthy, weakened, dead) was visually assessed at the end of each growing season for two years after planting. If a seedling had two or more leaders, or the leader growth was disturbed in some way, it was scored as having multiple leaders. In the first spring after planting (2015), just the cause and degree of damage (using the same classifications as above) and especially the existence of frost heaving (whether the peat plug had risen above the soil surface or not) was assessed.
One randomly selected seedling (from additional seedlings) per planting date, block and tree species, a total of 9 seedlings per date and species, was dug up from the trial in the nursery field three weeks after planting, and three times (on the 6th of May, on 20th of May and on 7th of October 2015) during the first growing season after planting. On the 6th and 20th of May, only seedlings planted during the previous autumn were sampled. Before digging, the shoot height, length of the leader growth, and diameter 2 cm above the ground surface were measured and the degree of damage was recorded. From the sampled seedlings, the roots grown out from the peat plug (longer than 5 mm) were counted each time and in autumn the total length of these roots was also measured (accuracy 1 mm). The dry mass of stem and needles was determined from the same seedlings from which roots were counted in autumn 2015. The shoot was divided as stem and needles, dried at 60 °C for 48 h, and dry masses were weighed (accuracy 0.01 g).
The start of the vegetative growth in both species was determined as described by Hänninen (2016) with few modifications. From the 22th of April 2015, bud burst in Norway spruce seedlings and the length of the new leader shoot in Scots pine seedlings (measurement was started from the same point at the base of bud each time; accuracy of 1 mm) were assessed in the nursery field. Two seedlings for each planting date and block, being a total of 18 seedlings in each species, were measured three times a week. In the spruce, buds were scored as flush when individual needle tips were visible. Bud burst was determined to occur on the date when 50% of observed seedlings had flush terminal buds on a planting date and block (the median day of bud burst). In Scots pine, the beginning date of shoot growth was determined to start when the height of a new shoot was at least 10 mm (median day of the beginning of height growth). For statistical analysis, the date was changed to the day of the year (DOY), but results are presented as a calendar date.
2.4 Weather conditions
At the weather station in Suonenjoki Research Unit of Luke, the monthly mean temperatures were 15.9, 10.3, 2.3, –0.4 and –3.9 °C for the months of August through December 2014, and –6.9, –2.2, –0.1, 3.2, 8.6, 12.4, 14.4, 16.0, 11.2 and 3.8 °C for the months of January through October 2015, respectively. The respective monthly precipitations were 101, 44, 53 and 31 mm for the months of August through November 2014, and 38, 71, 68, 90, 48, 63 and 26 mm for April through October 2015. Compared to a 15-year monthly average of temperatures and precipitations, autumn 2014 was consistent with the long-term average, except for August, which had an above average rainfall (long term 71 mm). In 2015, the temperatures were close to the average, but in May (long-term average 54 mm) and September (55 mm), the precipitation records were greater than average, while August and October (52 mm) were drier than the long-term average.
The first autumn frosts occurred on the 31st of August 2014, where the minimum temperatures at the soil surface were –0.2 °C. Snow cover was observed for the first time on 11th of December 2014 (5 cm) and it had melted by 11th of April 2015. The winter minimum temperatures (–23 °C) were measured on the 6th of January 2015, and on that day the depth of snow cover was 25 cm. In spring 2015, the last frosty night was on the 25th of May, where temperatures dropped to –1.9 °C on the soil surface.
The soil temperature at the forest site was measured by using two Hobo loggers (TMC20-HD). There were problems with the loggers, but at least one of them worked from 14th of August 2014 to November 2015. Temperatures were measured at depths of 6 and 12 cm (Fig. 1). In the spring and summer, the soil temperatures at the depth of 6 cm (the upper surface of a peat plug when seedlings were planted at the depth of 6 cm) were higher than temperatures at the depth of 12 cm, but they were lower in the autumn. The daily variation in temperatures was greater near the soil surface than in deeper in the soil profile throughout the season.
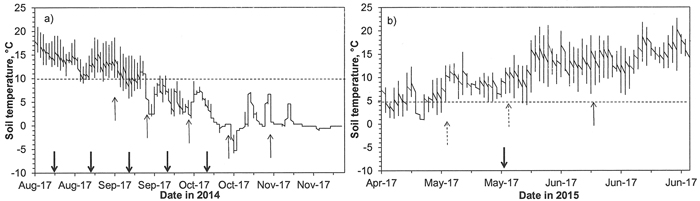
Fig. 1. Soil temperature measured at the depth of 12 cm (the bottom of a peat plug in a seedling planted to the depth of 6 cm) in a mound in a forest site a) just after planting in 2014, and b) in spring 2015. The thick arrows indicate the planting dates, thin arrows the dates when root growth three weeks after planting was assessed, and arrows with broken lines the dates when the initiation of root growth was measured in spring 2015. Broken horizontal lines are for +10 °C in a) and for +5 °C in b).
2.5 Data Analysis
The differences in shoot height, stem diameter, root growth, dry masses, the DOY when buds burst (or shoot growth started), probabilities for multiple leaders, and mortality, among planting dates within a trial were analyzed in IPM SPSS Statistics Version 22. Although the tree species were planted to the same field trial in a split-plot design in the nursery field, the tree species were analyzed separately. A repeated linear mixed model (MIXED) was applied to data for total shoot heights and diameters at the end of each year (the time since planting was repeated) and a linear mixed model for the root growth (each sampling time was analyzed separately), dry masses and the dates when buds burst or shoot growth started. A normal distribution was used in the MIXED model.
Probabilities for unburst buds, multiple leaders and mortality were analyzed with a generalized linear mixed model (GENLINMIXED). In the analyses, planting date (D) was considered as a fixed effect, while block was considered as the random effect. I employed a binomial distribution with a logit-link function in the GENLINMIXED models. Multiple comparisons were based on Least Significant Difference. Differences with a p-value <0.05 were considered to be significant.
Normal distribution was tested with the Kolmokorov-Smirnov test and the homogeneity of variances with Levine’s test. Log-transformation was used to homogenize variances in the number of roots in pine seedlings in 2015. For the first sampling (three weeks after planting), data was analyzed without transformation, although variances were heterogeneous (the transformation did not help due to zero values in the two last planting dates in autumn).
3 Results
3.1 Nursery field trial
3.1.1 Root growth and morphology
During a three week period after planting, the growth of the new roots slowed down when Norway spruce and Scots pine seedlings were planted in the middle of September and ceased later in the autumn (p < 0.001 for both species; Fig. 2). In both species, only small differences in rooting immediately after planting between plantings in August, early September or the following May were observed.
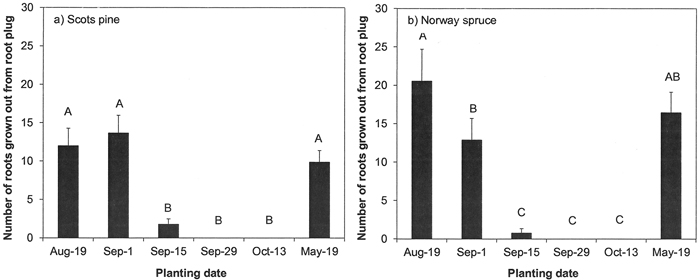
Fig. 2. Root growth of a) Scots pine and b) Norway spruce seedlings immediately after planting. Root growth was assessed as the number of roots grown out from a peat plug during a three-week period after planting in the nursery field. Vertical bars indicate standard errors of means (N = 9). Letters above bars indicate statistically significant (p < 0.05) differences among the planting dates.
In the following spring after autumn-plantings, on 6th of May 2015, no buds in Norway spruce had burst and the lengthening of new shoots had not started in Scots pine. On that day, the number of new root tips in Norway spruce was greater in the August-planted seedlings than in other autumn-planting dates (p = 0.003, Fig. 3b). In Norway spruce, the total number of roots grown out from peat plugs was on average 35, 20, 9, 11 and 8 from first to last planting date, respectively, and the two first dates differed statistically from the other dates (p < 0.001). On the same day, the number of new root tips in Scots pine seedlings did not differ among planting dates (p = 0.059), although the trend was that October-planted seedlings had the fewest number of new white root tips (Fig. 3a). The trend in the total number of roots grown out from peat plugs corresponded to the spruce and was on average 35, 27, 17, 9 and 10 from the first to last planting date, respectively, and the two first dates differed statistically from the other dates (p < 0.001).
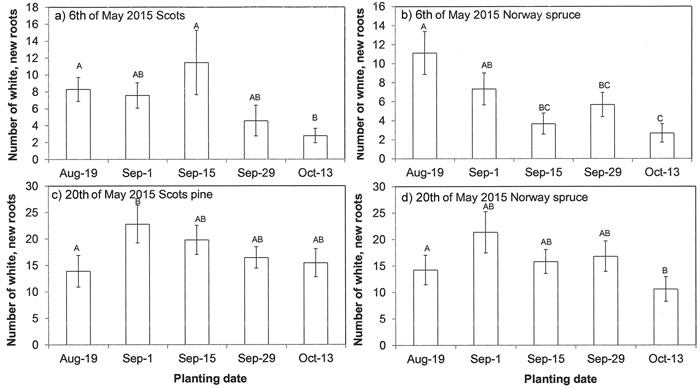
Fig. 3. Root growth of a, c) Scots pine seedlings and b, d) Norway spruce in the following spring after autumn plantings. Root growth was assessed as the number of new white roots grown out from a peat plug until a, b) 6th of May and c, d) 20th of May 2015 in the nursery field. Vertical bars indicate standard errors of means (N = 9). Letters above bars indicate statistically significant (p < 0.05) differences among planting dates.
On the 20th of May 2015, at the time of spring planting, root growth was assessed again. On that day, no buds in spruce seedlings had burst, but 15% of seedlings had swollen buds. In pine, new shoots started to grow. No statistically significant differences in the number of new roots among planting dates were found for either species (p = 0.414 for spruce and p = 0.216 for pine; Fig. 3c,d). The total number of roots grown out from a peat plug was higher in the two first planting dates in the autumn than in later autumn planted seedlings in both species (p < 0.001 for both species: data not shown)
At the end of the first growing season, no differences in the number and dry mass of roots grown out of a peat plug (new roots) or dry mass of roots inside a peat plug (old roots) or the dry mass of needles or stems among planting dates were found for either tree species (p-values for D 0.171–0.943; Fig. 4).
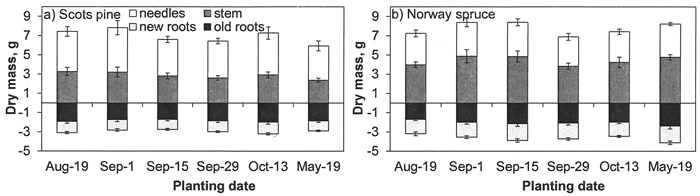
Fig. 4. Dry mass of needles, stems and new (roots grown out from peat plugs after planting) and old (roots inside a pleat plug) roots in a) Scots pine and b) Norway spruce seedlings in the nursery field at the end of first growing season. Vertical bars indicate standard errors of means (N = 9).
3.1.2 Bud burst and shoot growth
Scots pine flushed earlier in May than Norway spruce. The average day when buds of Norway spruce seedlings burst in different planting dates varied from the 27th to the 31st of May 2015, without statistically significant differences among planting dates (p = 0.104). The spring-planted seedlings flushed the latest. In pine, the planting date affected the beginning of flush (p = 0.009). For seedlings planted during the previous autumn, the beginning date varied from the 13th to the 17th of May 2015 without statistically significant differences among planting dates. However, spring-planted pine seedlings flushed significantly later, on the 21st of May, compared to autumn plantings.
In pine, the height of the seedlings was the same in all planting dates (Table 1, Fig. 5a). At the end of second growing season, spring-planted pine seedlings were 2.5–4.5 cm shorter than seedlings planted on the 19th of August or the 1st of September. October-planted pine seedlings were thinner than seedlings on other planting dates (Table 1, Fig 5c). After the first year, spring-planted pine seedlings were still thinner than seedlings in other planting dates. At the end of the second season, the thickest seedlings were on the two first planting dates (19th of August and 1st of September).
| Table 1. Repeated (measuring year Y) mixed model analysis of variance for planting date (D) on the shoot height and diameter of Norway spruce and Scots pine seedlings planted in the nursery field. The variance of random effects of each planting year and block are also presented. | ||||||||
| Source | Spruce | Pine | ||||||
| Height | Diameter | Height | Diameter | |||||
| Fixed effects | F | p-value | F | p-value | F | p-value | F | p-value |
| Intercept | 12 978 | <0.001 | 13 958 | <0.001 | 12 335 | 0.004 | 17 800 | <0.001 |
| Planting date (D) | 0.5 | 0.796 | 3.0 | 0.012 | 2.5 | . | 10.8 | <0.001 |
| Year (Y) | 1107 | <0.001 | 2120 | <0.001 | 2823 | <0.001 | 3526 | <0.001 |
| Y × D | 1.7 | 0.079 | 1.7 | 0.082 | 3.1 | 0.067 | 8.5 | <0.001 |
| Variance estimates for repeated planting year effect and block effect | ||||||||
| At planting | 3.49 (0.94) | 0.06 (0.01) | 3.3 (45) | 0.01 (0.01) | ||||
| First year | 19.13 (2.30) | 0.54 (0.06) | 7.3 (7.9) | 0.7 (0.1) | ||||
| Second year | 53.05 (5.26) | 2.98 (0.27) | 34.8 (7.6) | 4.0 (1.1) | ||||
| Block effect | 7.26 (<0.01) | 0.09 (<0.01) | 8.24 (<0.01) | 0.07 (<0.01) | ||||
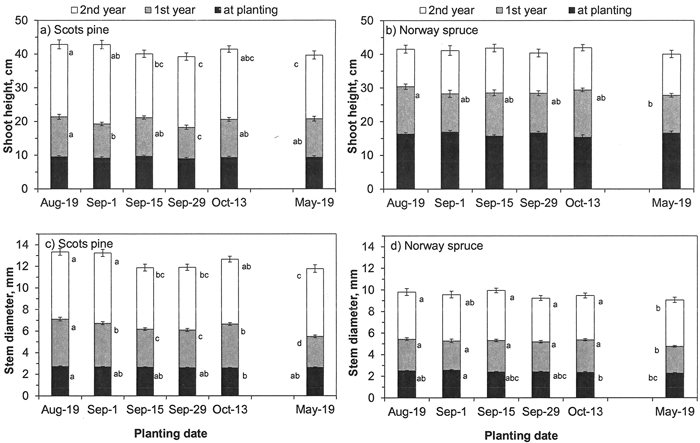
Fig. 5. a, b) Shoot height and c, d) stem diameter of a, c) Scots pine and b, d) Norway spruce seedlings planted in the nursery field in Suonenjoki biweekly in autumn 2014 and once in spring 2015. Height and diameters were measured at planting and at the end of two growing seasons after planting. Vertical bars indicate the standard errors of means and different letters indicate statistically significant differences (p < 0.05) among planting dates within a measuring year (N = 5 seedlings in 9 blocks).
In spruce, no statistically significant differences in height development among planting dates were found in the nursery field (Table 1, Fig. 5b). At planting, spring-planted seedlings were thinner than seedlings planted in August and the beginning of September and they also grew less than all the autumn planted seedlings during the first year (Fig. 5d). In the second year, no statistically significant differences in diameter were found among planting dates.
In autumn 2016, 0–7% of seedlings had multiple leaders without statistically significant differences among the planting dates (p = 0.916 for pine and p = 0.999 for spruce). All spruce seedlings survived, and only two spring-planted pine seedlings were dead at the end of the second season.
3.2 Trials in forest
The mortality of seedlings was low. No spruce seedlings died and 3% (varying between 0and 7%) of pine seedlings died without a statistically significant difference among the planting dates (p = 0.947). No frost heaved seedlings were observed during the first spring.
At this site, the planting date affected the development of the shoot height and stem diameter of both Scots pine and Norway spruce seedlings (see Y × D in Table 2). In pine, differences were small at planting, but spruce seedlings planted in October and May were the tallest and thickest (Fig. 6). Up to the end of the second season, no big differences in shoot height among autumn-planting dates were found in pine, but the May-planted seedlings were 5–10 cm shorter than seedlings planted in August and at the end of September. In Norway spruce, seedlings planted on 29th of September and in May were 4–8 cm shorter than seedlings planted on August and earlier in September (Fig. 6b). May-planted spruce seedlings were also thinner than seedlings in other planting dates at the end of the second season (Fig. 6d).
| Table 2. Repeated (measuring year Y) mixed model analysis of variance for planting date (D) on the shoot height and diameter of Norway spruce and Scots pine seedlings planted at the forest site. Variances of random effects of each planting year and block are also presented. | ||||||||
| Source | Spruce | Pine | ||||||
| Height | Diameter | Height | Diameter | |||||
| Fixed effects | F | p-value | F | p-value | F | p-value | F | p-value |
| Intercept | 8489 | <0.001 | 12 621 | <0.001 | 3159 | <0.001 | 4806 | <0.001 |
| Planting date (D) | 3.3 | 0.007 | 4.9 | <0.001 | 0.6 | 0.686 | 1.7 | 0.124 |
| Year (Y) | 1107 | <0.001 | 2424 | <0.001 | 902 | <0.001 | 931 | <0.001 |
| Y × D | 6.5 | <0.001 | 7.1 | <0.001 | 3.0 | 0.002 | 4.6 | <0.001 |
| Variance estimates for repeated planting year effect and block effect | ||||||||
| At planting | 6.46 (0.92) | 0.01 (0.01) | 4.9 (0.4) | 0.1 (0.01) | ||||
| First year | 20.44 (2.13) | 0.63 (0.06) | 31.2 (2.8) | 1.0 (0.1) | ||||
| Second year | 102.75 (9.31) | 2.27 (0.18) | 266.2 (23.7) | 11.9 (1.1) | ||||
| Block effect | 4.05 (<0.01) | 0.11 (<0.01) | 0 | 0.008 | ||||
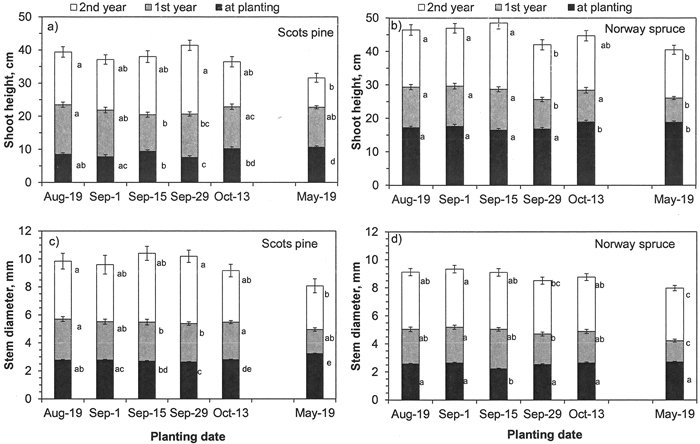
Fig. 6. a-b) Shoot height and c-d) stem diameter of a, d) Scots pine and b, d) Norway spruce seedlings planted in the forest site biweekly in autumn 2014 and once in spring 2015 and measured at planting and at the end of two growing seasons after planting. Vertical bars indicate the standard errors of means and different letters indicate statistically significant differences (p < 0.05) among planting dates within a measuring year (N = 43 for Scots pine and N = 45 for Norway spruce).
13% and 31% of pine seedlings had multiple leaders in the inventories in autumn 2015 and 2016, respectively, although no significant differences among planting dates were found (p = 0.074 in 2015; p = 0.856 in 2016). 6% and 8% of spruce seedlings had multiple leaders, in autumn 2015 and 2016, respectively. However there were no significant differences among the planting dates (p = 0.525 in 2015 and p = 0.656 in 2016). No clear reasons for multiple leaders were observed in the inventory.
4 Discussion
Root growth is important to the good field performance of seedlings. In this study, the most important result was the observation that the root growth in the previous autumn affected the initiation of the root growth in the following spring. There was a clear trend that root growth slowed down from August to September and root growth ceased in late-September and October. The root growth differences in the autumn correlated with the initiation of root growth in the following spring, when root growth in well-rooted seedlings, i.e. August-planted ones, started earlier than root growth in late-autumn planted, unrooted seedlings. However, these differences in root growth in autumn and the following spring affected the field performance of Scots pine and Norway spruce seedlings only little.
The root growth of conifers in natural soil conditions in areas having low winter temperatures usually ceases in September and October, but in some years root growth may continue into November and December in temperate forest zone (Lyr and Hoffmann 1967). According to Lyr and Hoffmann (1967), the minimum temperature for root growth of Norway spruce seedlings is 2–4 °C. However, root growth of newly planted seedlings ceases in higher temperatures. Similarly as Wray (1998) I observed that root growth slowed down at about the same time when the soil temperature dropped below +10 °C in September. In more southern areas with higher temperatures, a longer photoperiod and more amount of light in autumn, root growth may continue a few weeks longer as was the case in the study of Wallertz et al. (2016). The previous observations that the growth of roots in conifer seedlings can continue in hydroponic cultures in October (Iivonen et al. 1999; Kaakinen et al 2004) or in favorable temperature conditions throughout the year (Mattson 1986) is not possible in newly planted seedlings under a natural declining photoperiod, light and temperature conditions combined with planting stress in late-autumn plantings.
The rapid rooting of newly planted seedlings is important to ensure the best chance of field performance (Grossnickle 2005 and reference therein). In this study, the mortality of seedlings was exceptionally low. Thus, I did not receive any confirmation to the hypothesis that a poor rooting of autumn planted seedlings increased seedling damage or reduced the seedlings’ ability to recover from them. In the autumn, the planting stress of newly planted seedlings is probably lower than on other planting dates, since there is normally a high soil water content and low atmospheric evaporative demand. According to Grossnickle (2005), the existing root system is adequate to supply water to the shoot system under these conditions. Thus, under favorable weather conditions the risk of damage was low in this study. In practical scale plantings, however, the risk of failures are greater when the nursery-to-out planting process takes a much longer time than in this experiment and the seedlings are stressed at the time of planting. Valtanen et al. (1986), Kinnunen (1989) and Luoranen and Rikala (2013) observed that survival varies a great between autumn- and spring-plantings. Similarly, as in those previous studies, no differences in the survival of container conifer seedlings between planting dates were observed.
Previously, Wallertz et al. (2016) observed that late-autumn planted and poorly rooted, Norway spruce seedlings did not recover from pine weevil feeding as well as the earlier planted, and better rooted seedlings. In the same study, there were some signs that the late autumn planted seedlings suffered more winter desiccation than earlier planted ones. In my study, well-watered and insecticide-treated seedlings were carefully planted to the depth of 6 cm in mineral-soil dominated mounds or in the sandy nursery field and the frost heaving and other winter damage, as well as pine weevil feeding were totally prevented. The results parallel with the observation of Luoranen and Rikala (2013) that container-grown Scots pine seedlings can be planted even up to November in Central Finland conditions without any effects on survival if the soil is still unfrozen and further weather conditions are favorable.
Frost heaving is thought to be a risk to unrooted seedlings. The weather conditions did not favor frost heaving in this study. Seedlings were also planted deeply (6 cm) in sandy soil. Deep planting has been shown to reduce the risk of frost heaving (Schwan 1994; Sahlén and Goulet 2002). Frost heaving is also more probable in finer textured soils (Örlander et al. 1991; de Chantal et al. 2006) than was in this study. In autumn plantings, deep planting is good way to prevent frost heaving.
Differences in root growth in the previous autumn affected the initiation of the root growth in the following spring. To my knowledge, there are no previous studies, where the start of root growth of differently rooted seedlings has been investigated in the spring. August-planted and well-rooted seedlings started their root growth just after the soil warmed up by a few degrees in the following spring, before any visible sign of bud swelling. This agrees with the observations of Lyr and Hoffmann (1967) that root growth in Scots pine continues independently of shoot growth when the soil temperature is high enough in the spring. The soil temperatures at the time of the first root sampling in spring was about +5 °C. This is in contrast to the result of Vapaavuori et al. (1992), who observed that root growth was almost prevented if the soil temperature was +8 °C and starts properly when the soil temperature rises above +12 °C.
In mid-May, there was still a trend that seedlings planted later had fewer roots than earlier planted seedlings, but no differences in the dry masses of roots were found at the end of the first growing season. This differs from the results of Wallertz et al. (2016), who observed that differences in the dry mass of roots remain until the end of the first autumn.
The differences in the initiation of the root growth in the spring and the size of root systems probably explain why the shoot growth of seedlings planted after mid-September was slightly lower than it was in the August and early-September-planted ones. The trend in shoot growth in relation to the planting date was similar in both species. Poorer growth of late-planted seedlings has also been observed by Luoranen and Rikala (2013) in Scots pine seedlings, in machine-planted Norway spruce seedlings by Luoranen et al. (2011) and in Norway spruce seedlings by Wallertz et al. (2016). Although differences in shoot height were small, it has been observed in previous studies that a few centimeters difference in height growth two years after planting may remain the same or even increase in later years (Luoranen et al. 2006; Johansson et al. 2013).
I did not observe any differences in flush either of tree species in the first spring after planting. This differs to some extent from the results of Luoranen and Rikala (2015) who observed that summer-planted (July, early August) seedlings flush earlier than autumn- and spring-planted Norway spruce seedlings. In this study, the first planting was in mid-August and the later plantings in September and October, and thus the probable differences among autumn planting dates were small in flush. Late flush of spring-planted seedlings is probably caused by freezer storage in which the seedlings were until the end part of April. Seedlings were moved to the outdoor growing area in the nursery near the forest edge, where seedlings were in a shadow during a part of a day. Planted seedlings were in an open field in direct sunlight. Thus, the accumulated temperature sum was probably lower for spring-planted seedlings than for autumn-planted ones.
In this experiment, I did not find big differences in survival or damage between tree species. Previously, Johansson et al. (2015) observed that Scots pine seedlings had more damage than Norway spruce seedlings. The shoot growth of Scots pine seedlings was better than that in Norway spruce, which corresponds to the results of Johansson et al. (2015).
In conclusion, late-autumn plantings of Scots pine and Norway spruce container seedlings can be successful, at least without harsh weather conditions in winter and spring and when seedlings are planted carefully without any stressors before and after planting. Poorly rooted, late-autumn planted seedlings start their root growth later during the following spring, which reduces the shoot growth of seedlings in later years. Risks related to late plantings, especially in more harsh weather conditions, need more studies before recommendations to plant in very late autumn can be made.
Acknowledgements
This research was conducted in co-operation with forest owner Dr. Antti Lukkarinen who carried out soil preparation in the forest area. I thank Osmo Korhonen and Esa Mölkänen for planting the seedlings, Pekka Voipio, Auli Lehtinen, Maija Sianoja, Juhani Korhonen and Ilkka Taponen for the seedling measurements and Dr. Risto Rikala, Dr. Sirkka Sutinen and two anonymous referees for valuable comments to the manuscript. The language was checked by AAC Global Oy. The study was solely financed by the Natural Recourses Institute Finland (former Finnish Forest Research Institute) through projects 3554, 1300355400 and 41007-00100100.
References
de Chantal M., Rita H., Bergsten U., Ottoson Löfvenius M., Grip H. (2006). Effect of soil properties and soil disturbance on frost heaving of mineral soil: a laboratory experiment. Canadian Journal of Forest Research 36(11): 2885‒2893. https://doi.org/10.1139/x06-181.
Grossnickle S.C. (2005). Importance of root growth in overcoming planting stress. New Forests 30(2–3): 273–294. https://doi.org/10.1007/s11056-004-8303-2.
Grossnickle S.C., Folk R.S. (2003). Spring versus summer spruce stocktypes of western Canada: nursery development and field performance. Western Journal of Applied Forestry 18(4): 267–275.
Hänninen H. (2016). Boreal and temperate trees in a changing climate. Modelling the Ecophysiology of Seasonality. Springer Science+Business Media B.V. Dordrecht. 342 p. https://doi.org/10.1007/978-94-017-7549-6.
Iivonen S., Rikala R., Ryyppö A., Vapaavuori E. (1999). Responses of Scots pine (Pinus sylvestris) seedlings grown in different nutrient regimes to changing root zone temperature in spring. Tree Physiology 19(14): 951–958. https://doi.org/10.1093/treephys/19.14.951.
Johansson K, Nilsson U., Örlander G. (2013). Comparison of long-term effects of scarification methods on the establishment of Norway spruce. Forestry 89: 91–98. https://doi.org/10.1093/forestry/cps062.
Johansson K., Hajek J., Sjölin O., Normark E. (2015). Early performance of Pinus sylvestris and Picea abies – a comparison between seedling size, species, and geographic location of the planting site. Scandinavian Journal of Forest Research 30(5): 388–400. https://doi.org/10.1080/02827581.2014.987808.
Kaakinen S., Jolkkonen A., Iivonen S., Vapaavuori E. (2004). Growth, allocation and tissue chemistry of Picea abies seedlings affected by nutrient supply during the second growing season. Tree Physiology 24(6): 707–719. https://doi.org/10.1093/treephys/24.6.707.
Kinnunen K. (1989). Effect of seedling type and site preparation on the initial development of Scots pine and Norway spruce seedlings. Folia Forestalia 727. (In Finnish with English summary). http://urn.fi/URN:ISBN:951-40-1040-X.
Luoranen J., Rikala R. (2013). Field performance of Scots pine (Pinus sylvestris L.) seedlings planted in disc trenched or mounded sites over an extended planting season. New Forests 44(2): 147–162. https://doi.org/10.1007/s11056-012-9307-y.
Luoranen J., Rikala R. (2015). Post-planting effects of early-season short-day treatment and summer planting on Norway spruce seedlings. Silva Fennica 49(1) article 1300. https://doi.org/10.14214/sf.1300.
Luoranen J., Rikala R., Konttinen K., Smolander H. (2005). Extending the planting period of dormant and growing Norway spruce container seedlings to early summer. Silva Fennica 39(4): 481–496. https://doi.org/10.14214/sf.361.
Luoranen J., Rikala R., Konttinen K., Smolander H. (2006). Summer planting of Picea abies container-grown seedlings: effects of planting date on survival, height growth and root egress. Forest Ecology and Management 237(1–3): 534–544. https://doi.org/10.1016/j.foreco.2006.09.073.
Luoranen J., Rikala R., Smolander H. (2011). Machine planting of Norway spruce by Bracke and Ecoplanter: an evaluation of soil preparation, planting method and seedling performance. Silva Fennica 45(3): 341–357. https://doi.org/10.14214/sf.107.
Lyr H., Hoffmann G. (1967). Growth rates and growth periodicity of tree roots. International Reviews of Forestry Research 2: 181–206. https://doi.org/10.1016/B978-1-4831-9976-4.50011-X.
Mattson A. (1986). Seasonal variation in root growth capacity during cultivation of container grown Pinus sylvestris seedlings. Scandinavian Journal of Forest Research 1(1–4): 473–482. https://doi.org/10.1080/02827588609382438.
Nilsson U., Luoranen J., Kolström T., Örlander G., Puttonen P. (2010). Reforestation with planting in northern Europe. Scandinavian Journal of Forest Research 25(4): 283–294. https://doi.org/10.1080/02827581.2010.498384.
Örlander G., Gemmel P., Wilhelmsson C. (1991). Effects of scarification, planting depth and planting spot on seedling establishment in a low humidity area in southern Sweden. Swedish University of Agricultural Sciences. Department of Silviculture. Report 33. 92 p. [In Swedish with English summary].
Sahlén K., Goulet F. (2002). Reduction of frost heaving of Norway spruce and Scots pine seedlings by planting in mounds or in humus. New Forests 24(3): 175–182. https://doi.org/10.1023/A:1021378228524.
Schwan T. (1994). Planting depth and its influence on survival and growth. Literature review with emphasis on Jack pine, black spruce and white spruce. Ontario Ministry of Natural Resources, Northeast Science & Technology, Technical Report TR-017. 35 p.
Söderström V. (1974). Influence of soil temperature on conifer plant growth – pilot study in the laboratory. Sveriges Skogsvårdsförbunds Tidskrift 72(5–6): 595–614. [In Swedish with English summary].
Tan W. (2007). Impact of nursery cultural treatments on stress tolerance in 1 + 0 container white spruce (Picea glauca [Moench] Voss) seedlings for summer-planting. New Forests 33(1): 93–107. https://doi.org/10.1007/s11056-006-9016-5.
Valtanen J., Kuusela J., Marjakangas A., Huurinainen S. (1986). Initial development of Scots pine and Siberian larch paper-pot seedlings planted at various times. Folia Forestalia 649. [In Finnish with English summary]. http://urn.fi/URN:ISBN:951-40-0732-8.
Vapaavuori E., Ryyppö A., Rikala R. (1992). Effects of root temperature on growth and photosynthesis in conifer seedlings during shoot elongation. Tree Physiology 10(3): 217–230. https://doi.org/10.1093/treephys/10.3.217.
Wallertz K., Hansen K.H., Hjelm K., Fløistad I.S. (2016). Effect of planting time on pine weevil (Hylobius abietis) damage to Norway spruce seedlings. Scandinavian Journal of Forest Research 31(3): 262–270. https://doi.org/10.1080/02827581.2015.1125523.
Wray P.H. (1998). Fall planting of bare root seedlings. Iowa State University, Forestry Extension Notes. 2 p.
Total of 26 references.
Send to email
