Pentti Niemistö 
Effect of growing density on biomass and stem volume growth of downy birch stands on peatland in Western and Northern Finland
Niemistö P. (2013). Effect of growing density on biomass and stem volume growth of downy birch stands on peatland in Western and Northern Finland. Silva Fennica vol. 47 no. 4 article id 1002. https://doi.org/10.14214/sf.1002
Highlights
- The thinning response in young downy birch stands was low and the mortality of merchantable stems was tolerable even in dense unthinned thickets during the rotation of 50 years
- The production of above-ground woody biomass and stem volume or even the production of pulpwood increased with stand density, with these values being greatest for unthinned or very lightly thinned plots.
Abstract
The objective of the study was to ascertain the relationship of thinning intensity of downy birch (Betula pubescens Ehrh.) stands with height, crown, and diameter development as well as pulpwood, stem volume, and biomass increment using long-term (20−30 years) field experiments. Diameter growth of birches increased with thinning intensity during the first 15 years from thinning in all development phases, though after that it did so only for the youngest stands. The thinning response was low. Thinning intensity had no influence on increase in height. In terms of stem volume with bark, the mortality in unthinned stands during the study period was 30–45 m3 ha-1. The mean stem number in unthinned birch thickets fell from 25 000 ha-1 at a dominant height of 7 m to 3000 ha-1 at 18 m. The stem volume increment over the first 15 years was highest (5–6 m3 ha-1 a-1) on the very lightly thinned or unthinned plots, but later there was no significant difference between initial thinning intensities. The maximum above-ground leafless biomass (over 100 Mg ha-1) was achieved on very lightly thinned plots. Also, the total production (including thinning removal) of biomass or stem volume or even the production of pulpwood increased with stand density, with these values being greatest for very lightly thinned or unthinned plots. During 50-year rotation, the highest leafless above-ground biomass production was 2.5 Mg ha-1 a-1 as a mean value from the experiments. The highest mean annual production of pulpwood (d > 6.5 cm) was 3.2 m3 ha-1 a-1, and, in practice, no saw timber or veneer timber was produced, because of the small size and low quality of the stems. A thinning in downy birch stands increased slightly the size of stems to be removed in future cuttings, but with exception for very light thinning it decreased the production of biomass and merchantable wood.
Keywords
yield;
thinning;
silviculture;
mortality;
pulpwood;
stand density;
crown;
energy-wood
-
Niemistö,
Finnish Forest Research Institute, Parkano Unit, FI-39700 Parkano, Finland
E-mail
pentti.niemisto@metla.fi
Received 12 April 2012 Accepted 31 May 2013 Published 24 October 2013
Views 212085
Available at https://doi.org/10.14214/sf.1002 | Download PDF

1 Introduction
Downy birch (Betula pubescens Ehrh.) is one of the most common tree species in Northern Europe, growing mostly as a mixture in pine- or spruce-dominated stands. The economic value ascribed to downy birch is not very high, because of the lower growth capacity and stem quality in comparison with conifers or silver birch (Betula pendula Roth) (Koivisto 1959; Fries 1964; Braastad 1967; Cameron et al. 1995; Verkasalo 1997; Hynynen et al. 2010).
Downy birch stands represent a common and typical low-budget forestry pattern in northern and western Finland, especially on peatlands and wet mineral soils. Natural regeneration of dense birch stands is favourable and low-priced. At the same time, the economic value of downy birch is low, because of the small stem size and low stem quality, but new interest is being turned to energy-wood production.
Downy birch is slightly more commonplace in cooler, more northern and humid areas than silver birch (Hultén and Fries 1986). On drained peatlands and wet mineral soils downy birch is often the first pioneer species after drainage and can be competitive with conifers (Keltikangas and Seppälä 1977; Saramäki 1977; Moilanen 1985). In western and northern Finland, pure birch stands of this sort are quite common (Niemistö and Korhonen 2008). In addition, a downy-birch-dominated stand can result from a failed regeneration operation or overdue tending treatment. The density of young stands dominated by downy birch is often very high, tens of thousands of stems per hectare (Ferm and Kauppi 1990; Ferm 1993). The practical recommendation in Finland for pulpwood production has been quite dense sapling stands (2000–2500 stems ha–1) and one light commercial thinning in the course of the rotation. The knowledge, so far, has been insufficient for estimating biomass production and inclusive management patterns over the rotation.
Control of stand density through thinnings has been a tool frequently used in increasing per-tree growth and improving stem quality. While thinning from below may increase merchantable volume (saw timber and even pulpwood), it has usually decreased the total volume increment per unit area (e.g., Assman 1970; Zeide 2001; Mäkinen and Isomäki 2004a and 2004b). Many studies have shown the merchantable volume increment not to decline as stand density falls, over a wide range of stand densities (e.g. Eriksson and Karlsson 1997). In downy birch stands on peatland, such a result has not been achieved, but both total and merchantable volume increment has increased with stand density (Niemistö 1991). Saw timber or plywood production has been very low in thinned downy birch stands (Verkasalo 1997) because of small stems and low quality.
In addition to the weak response to thinning, also some ecological aspects corroborate also the favour of light thinning in downy birch stands. Birches are considered shade-intolerant pioneer tree species (Nygren and Kellomäki 1983), but downy birch is able to survive and grow also with little exposure to light (Atkinson 1984). Modest rising of the water table has been seen in peaty soil after light thinning of pine stands (Päivänen and Sarkkola 2000). For birch stands, especially after heavier thinning, this influence will probably be greater and reduce the growth response (Niemistö 1991).
The demand for energy-wood has increased over the last decade. Also, the prices of energy-wood and pulpwood have come closer to parity. This calls for a new approach to management in downy birch stands. According to the prevailing hypothesis the highest total production is achieved in unthinned stands, but the benefits of thinning can be lower mortality and higher pulpwood production in this study. In addition to a traditional thinning intensity study of stem and merchantable volume production this study provides a new approach for biomass production, with the aid of 19 long-term field experiments established in dense birch stands at dominant heights ranging from four to 15 metres. This study is a sequel to a report of Niemistö (1991), which is based on the seven experiments used also in this work.
The objective of this study was to assess thinning intensity in relation to tree height, stem diameter, and crown length development as well as stem volume and biomass increment in different phases of stand development. Most of the experiment stands have been studied for 25–30 years and either are approaching maturity or are already mature. Therefore, the study was able to monitor the thinning removals, mortality, and total stem volume and biomass production during the entire stand rotation. Total biomass of growing stock and removals in downy birch stands were studied in terms of separate components: stem wood, stem bark, living branches, dead branches and stumps.
2 Material and methods
2.1 Experiments
The material was collected from 19 experiments in Ostrobothnia and western Lapland, established by the Finnish Forest Research Institute. The experiment stands were divided into three classes according to the mean height of the birches at the onset of the experiments (Table 1). Nine experiments were established at the sapling stand stage (SS) at a mean height of 3–8 m, mostly in the years 1984–1990. Seven experiments were started at a mean height of 8–11 m and three experiments at a mean height of 12–13 m, mostly in 1975–1983. The height class 8–11 m is considered to be the energy-wood thinning stage (EW) and 12–13 m to be the pulpwood thinning stage (PW).
| Table 1. Mean characteristics of the experiments at establishment in the three height classes: Sapling stand stage (SS: H0 < 8m), Energy wood thinning stage (EW: 8m < H0 < 11m) and Pulpwood thinning stage (PW: 12m < H0 > 13m). View in new window/tab. |
The stands were pure or almost pure downy birch stands on drained peatlands and were mostly seed-originated, apart from two experiments that involved partial growth from sprouts and a whole young coppice. The sites were herb-rich type or vaccinium myrtillus type, typical for birch-dominated drained peatlands. The site index (H50, dominant height at the age of 50, Table 1) ranged from 13 to 19 (in metres). Downy birches have mainly born following the drainage and in most cases the ditch network maintenance took place prior to establishment of an experiment.
The experiment design involved mostly 2–3 replicates of 3–4 thinning intensities, controlled for the stem number of the growing stock, as well as unthinned plots. A second thinning 10–15 years after the first one, took place on 15 plots in EW and PW height classes both (19% and 28% of the total number of plots, respectively). The area of a rectangular plot (from ditch to ditch) ranged from 800 to 1400 square metres. Each experiment included 5–30 plots; i.e., in the main, the treatments were replicated 3–4 times in a randomised block design, in some cases even 5–6 times. However, some experimental stands included only one or two replicates because of small area.
2.2 Measurements and statistical analysis
After establishment, the experiment stands were re-measured 3–6 times, mostly in five-year periods (Table 1). At the time of final measurement, the stand age was 30–54 for SS height class, 40–65 for EW, and 67–84 for PW. Tree species, stem diameter at breast height (d1.3), any damage, and mortality were recorded for each tree on the plot. If the stem number was very high (a situation seen mostly on control plots), 1–3 small circular sample plots were measured instead of the whole plot area. In the selection of sample trees (20–40 per plot), the probability of a tree being selected was proportional to its diameter, but these trees were distributed randomly over a plot. Tree height (h) was measured for each sample tree from the beginning of the experiments. Height to living crown base and stem diameter at a height of 6 m (d6.0) were measured after the age at which the trees reached a height of approximately 10–11 metres.
The characteristics of individual plots – including mean stem measurements, basal area, and stem volume in three dimensions: veneer logs, pulpwood, and small-sized (see below) round-wood – were calculated by means of the KPL software, developed at the Finnish Forest Research Institute (Heinonen 1994). The stem volumes with bark of the sample trees were calculated via volume functions (Laasasenaho 1982) from d1.3 and h (and d6.0 when measured). The height of the other trees was predicted as a function of diameter from the height curve of Näslund (1937), which was fitted to each plot with the aid of the tree heights measured for the sample trees. The stem volume with bark of other trees was calculated with smoothing functions (v/d1.32 = a0 + a1d1.3 + a2d1.32) fitted to the sample trees on each plot. The minimum length applied for pulpwood boles was 3.0 m and the minimum top diameter 6.5 cm over bark. Stem wood below this size was considered to be small-sized round-wood. The minimum length applied for veneer logs was 3.0 m, and the minimum top diameter 18.0 cm over bark.
The biomass of every tree was calculated in terms of five components (Repola 2008 and 2009): stem wood, stem bark, living branches, dead branches and stumps. The biomass models were based on tree species, diameter and height in the beginning, when live crown was not measured (multivariate model 1 in Repola 2008). Later on, crown base was measured, and the multivariate model 2 was used with additive variables age and crown ratio. According to Jaakko Repola (Finnish Forest Research Institute, pers. comm. 2009) no bias for stem and branch biomass, caused by stand density, existed when the models were tested in an independent data collected from downy birch stands with 5000−30 000 stems ha–1 (Ferm 1990a; Ferm and Kaunisto 1983; Ferm and Kauppi 1990; Hytönen and Aro 2012). For the stand level measures, the stem volume (m3 ha–1 with bark) and biomass (dry material: Mg DM ha–1) components of the trees were summed for each plot and measurement. To calculate mean annual increment, we took the difference between successive measurements and divided this value by the number of years between the measurements.
The treatments were specified and plots classified on the basis of the stem number of the growing stock after the initial thinning on the establishment of the experiments (see Fig. 1). The thinning groups used were ‘Heavy’, ‘Moderate’, ‘Light’, ‘Very light’, and the non-thinned ‘Control’. Other grouping was based on a second thinning (applied or not 10–15 years after first one), that decreased the stem number to half on the sub-sample of lightly or very lightly thinned plots. There existed differences in stem numbers for corresponding treatments between the height classes SS, EW, and PW, especially within the light thinnings and the control. The reason for this was the original stem number, which was very high in the sapling stands (SS: 11 000–40 000 stems ha–1) and low in pulpwood thinning stands because of earlier tending (PW: 1900–2500 stems ha–1).
The differences among the treatments were analysed through covariance analysis including random block effect (where a block was an experimental stand). The three height classes were analysed separately because of differences in age, thinning status (commercial or not), sites, and the length of the study period. The model used to test the treatment effects was
![]()
where Ysp is the dependent variable, µ is the overall mean, δT is the vector of treatment effect (first and second thinning), and β and Xsp are the vectors of regression coefficients and covariates. Parameters us and usp are the random effects for stand s and plot p. The differences among the plots were removed in most cases through application of a continuous covariate H50 based on dominant height at the age of 50 years (measured or interpolated in EW and PW height classes but extrapolated in SS). Dominant height is defined as the mean height of the 100 largest trees per hectare. In addition to H50, dominant height or age at the end of the study period were used as covariates, as was the length of the study period. The covariates used in the models are reported in the figures and tables, and the estimated mean values are adjusted by setting covariate effects to their mean values. For the statement of standard deviation in some tables, the original mean values are also reported besides the estimated ones.
The pairwise comparisons were performed through computing of generalised least-square means of the treatment effects. In the figures and tables, treatments denoted with the same letter are not significantly different from each other (p > 0.05). Restricted maximum likelihood (REML) estimation in the ‘Mixed linear’ procedure was used in the analysis (SPSS 16.0, PASW Statistics).
3 Results
3.1 Tree growth
The dominant height increment was examined in two periods, years 1 to 15 from first thinning and years 16 to final measurement. For the first period, the mean of dominant height increment was 25 cm a–1 in all three height classes: SS, EW, and PW. It was independent of thinning intensity (p = 0.8–0.9). In the second period, the increment was still the same in SS height class but 18 cm a–1 for EW and 11 cm a–1 in PW. No differences were found in relation to the initial thinning densities, nor did the second thinning have an influence on dominant height growth. The culmination of the height increment was at a dominant height of 13–14 m in EW height class. In the youngest stands (SS), the culmination could not be detected and in mature stands (PW) it was probably passed by.
The mean stem number for unthinned birch thickets decreased from 25 000 ha–1 at a dominant height of 7 m to 3000 stems ha–1 at a dominant height of 17–18 m (Fig. 1). On lightly thinned plots, the number of live trees decreased to 1000 ha–1 at approximately 20 m dominant height. The mean length of live crowns in full dense young birch thickets was below 3 m for a dominant height of 10 m (Fig. 2). Later it increased to above 5 m at a 15 m dominant height because of dying small trees. Increasing thinning intensity extended the crown length linearly with time. For heavily thinned stands, the mean crown length increased from five metres at the beginning to nine metres at the end of the study period.
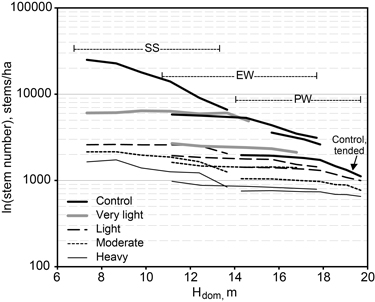
Fig. 1. The mean stem number (on a logarithmic scale) of live trees by first thinning intensity and dominant height during the study period, when the thinning was carried out at the sapling stand stage (SS), at the energy wood thinning stage (EW), and at the pulp wood thinning stage (PW).
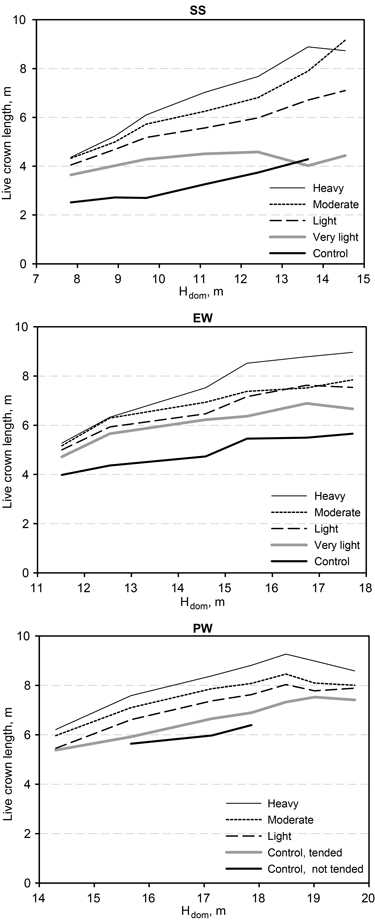
Fig. 2. Estimated live crown length of growing stock as a function of dominant height and treatment of downy birch stands. Dominant height was used as the covariate in Eq. 1 for crown length (i.e., tree height less live crown base). The mean dominant height at each measurement and in each height class (SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage) was used in calculation of the estimated means.
At the start of the experiments, the length of live crowns was 50–60% of tree height. In unthinned control plots and very light thinning, this crown ratio fell to 40% in the course of the study period. In light thinning, it stayed at the same level all the time. After moderate thinning, the crown ratio rose at first but then returned to its initial level. After heavy thinning, it showed an increase of 10 percentage points from its initial value.
Because the development of dominant height was independent of thinning, the site index (H50) based on it was used as a covariate in modelling of the influence of thinning intensity on other growth and yield variables. Thus the influence of variation among sites could be eliminated from the results. Two preceding periods, 1–15 years and year 16 to final measurement (or ‘16–Fm’), were used in the analyses. So the second thinning affected only results for the latter period. The effects of the second thinning will be presented later in section 3.4.
The mean diameter increment of all living trees on the plots increased with decreasing stand density (Table 2). The differences between thinning intensities were statistically significant in the 1–15-year period after first thinning in all three height classes. In the latter period 16−Fm, there still existed significant differences associated with the initial thinning intensities in SS height class and between heavy thinning and other treatments in EW – but not in PW. The unthinned control showed a divergent trend above the curve for SS height class because of several dead small trees in the latter period excluded from calculations of mean diameter.
| Table 2. Annual mean diameter increment of live trees in two periods for plots of different initial thinning intensities and height classes. Treatments marked with the same letter (within a height class and growth period) are not significantly different (p > 0.05). Site index, H50, was used as the covariate in Eq. 1, and the estimated mean increment was calculated on the basis of the overall mean of H50 (15.3 m) across the entire dataset. | ||||||
| Treatment | Mean diameter growth (mm a–1) in years 1−15 | Mean diameter growth (mm a–1) years 16−Fm (final measurement) | ||||
| Mean for the data | Standard deviation | Estimated mean | Mean for the data | Standard deviation | Estimated mean | |
| SS, sapling stand stage | ||||||
| Heavy | 3.63 | 0.90 | 3.74a | 2.68 | 0.47 | 2.79a |
| Moderate | 2.98 | 0.78 | 2.94b | 2.28 | 0.53 | 2.35ab |
| Light | 2.35 | 0.51 | 2.32bc | 1.71 | 0.60 | 1.83bc |
| Very light | 1.62 | 0.33 | 1.64cd | 1.09 | 0.52 | 1.16c |
| Control | 0.77 | 0.40 | 0.87d | 1.84 | 0.78 | 1.85bc |
| EW, energy-wood thinning stage | ||||||
| Heavy | 3.47 | 0.63 | 3.54a | 1.82 | 0.58 | 1.88a |
| Moderate | 2.76 | 0.53 | 2.79b | 1.15 | 0.35 | 1.37b |
| Light | 2.24 | 0.61 | 2.43bc | 1.14 | 0.36 | 1.23b |
| Very light | 1.92 | 0.64 | 2.15c | 0.87 | 0.11 | 1.22b |
| Control | 0.87 | 0.30 | 1.02d | 1.06 | 1.07 | 1.02b |
| PW, pulpwood thinning stage | ||||||
| Heavy | 2.64 | 0.53 | 2.59a | 1.02 | 0.27 | 1.00a |
| Moderate | 1.95 | 0.47 | 2.12b | 0.74 | 0.07 | 0.78a |
| Light | 1.89 | 0.49 | 1.73c | 0.71 | 0.17 | 0.75a |
| Control. tended | 1.55 | 0.26 | 1.34d | 0.60 | 0.16 | 0.64a |
| Control. not tended a) | 0.72 | 0.14 | 1.17cd | |||
| a) Includes only two plots in one experimental stand. | ||||||
Because the difference in the mean increment values may be caused by differing numbers of stems/ha, mean stem volume with bark at final measurement was calculated for the 600 largest trees per hectare (Fig. 3). The mean volume of the largest trees tended to increase as stand density decreased, but significant differences existed only between heavy or moderate thinning, on one hand, and light thinning or the unthinned control, on second hand, in SS height class, as well as between heavy thinning and the unthinned control in EW. In mature (PW) stands, no significant differences were seen on the basis of the first thinning intensities.
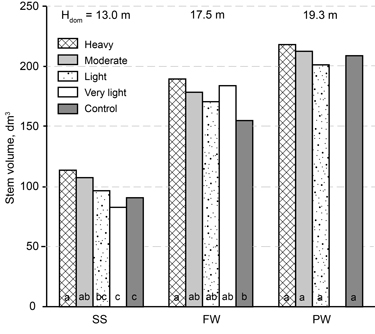
Fig. 3. The influence of thinning intensity on the mean volume of the 600 largest birch stems per hectare at the final measurement in height classes: SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage. Treatments denoted by the same letter (within a height class) are not significantly different (p > 0.05). Site index (H50) and stand age at final measurement were used as the covariates in Eq. 1, and the estimates were calculated through the use of their mean values separately for each height class.
The total stem volume increment on the plots increased with stand density (Table 3). Differences between stands with different thinning intensities were statistically significant in the 1–15-year period after first thinning, in all three height classes. In the latter period, the same trend could be seen but significant differences existed only in class EW between heavy or moderate thinning, on one hand, and the unthinned control.
| Table 3. Mean annual stem volume increment in two periods for plots with different initial thinning intensities and height classes. For the models, covariates, estimated means and statistical significance, refer to Table 2. | ||||||
| Treatment | Mean volume increment (m3 ha–1 a–1) in years 1−15 | Mean volume increment (m3 ha–1 a–1) in years 16−Fm (final measurement) | ||||
| Mean for the data | Standard deviation | Estimated mean | Mean for the data | Standard deviation | Estimated mean | |
| SS, sapling stand stage | ||||||
| Heavy | 3.01 | 0.84 | 2.97a | 4.37 | 1.50 | 4.81a |
| Moderate | 3.60 | 1.10 | 3.54a | 5.24 | 1.60 | 5.37a |
| Light | 4.27 | 1.25 | 4.40b | 5.26 | 1.11 | 5.56a |
| Very light | 4.89 | 1.35 | 5.38c | 5.84 | 1.50 | 6.17a |
| Control | 5.73 | 1.59 | 5.97c | 6.65 | 1.99 | 7.00a |
| EW, energy-wood thinning stage | ||||||
| Heavy | 4.14 | 1.10 | 3.89a | 4.16 | 1.18 | 4.05a |
| Moderate | 5.04 | 1.18 | 4.65b | 3.99 | 0.83 | 3.96a |
| Light | 5.33 | 1.00 | 5.07bc | 5.22 | 1.75 | 4.85ab |
| Very light | 6.48 | 1.58 | 6.14d | 5.49 | 2.99 | 5.50ab |
| Control | 5.81 | 0.84 | 5.74cd | 6.14 | 2.27 | 5.71b |
| PW, pulpwood thinning stage | ||||||
| Heavy | 4.73 | 1.21 | 4.20a | 2.71 | 0.70 | 2.41a |
| Moderate | 4.85 | 1.15 | 4.69ab | 2.78 | 0.99 | 2.94a |
| Light | 5.18 | 0.94 | 4.79ab | 2.96 | 0.99 | 3.00a |
| Control. tended | 6.13 | 1.07 | 5.30b | 3.41 | 0.70 | 3.36a |
| Control. not tended a) | 5.88 | 0.47 | 6.87c | |||
| a) Includes only two plots in one experimental stand. | ||||||
Because of mortality, the mean net volume increment (the volume of dead trees removed from the volume growth) on the plots was lower than the total increment, especially for dense tree stands (Fig. 4). The same trend as in total volume growth existed in the time 1–15 years after the first thinning except in unthinned control plots, where the net volume increment was lower than that with light thinning in SS and EW height classes. In the latter period, the net volume increment was independent of first thinning intensity in all height classes.
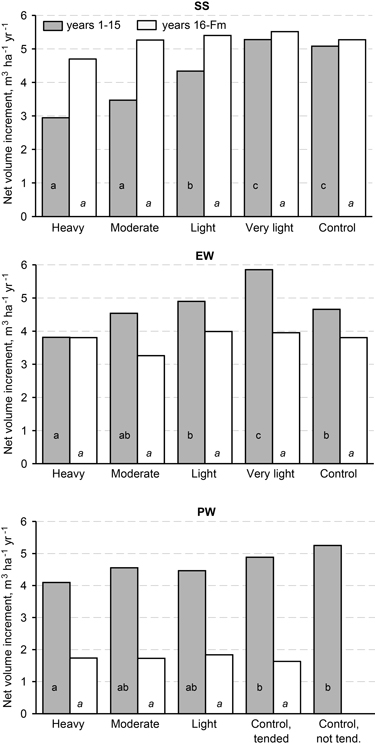
Fig. 4. Mean net volume increment estimates on plots of different thinning intensity and height class in two periods in height classes: SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage. For the model used, covariates, estimated means and statistical significance, refer to Table 2.
3.2 Biomass development
The estimated mean development of the main biomass components of live trees on the plots is presented in Fig. 5. Biomass components increased and differences between stands of different thinning intensities decreased step by step over the 20–30-year study period. The differences were still significant at the end of the study period, except for the unthinned control versus very light thinning in SS and EW height classes and moderate thinning versus light thinning in EW and PW. The other exception was the biomass of branches, which increased rapidly on thinned plots in the first half of the study period but not in the second half.
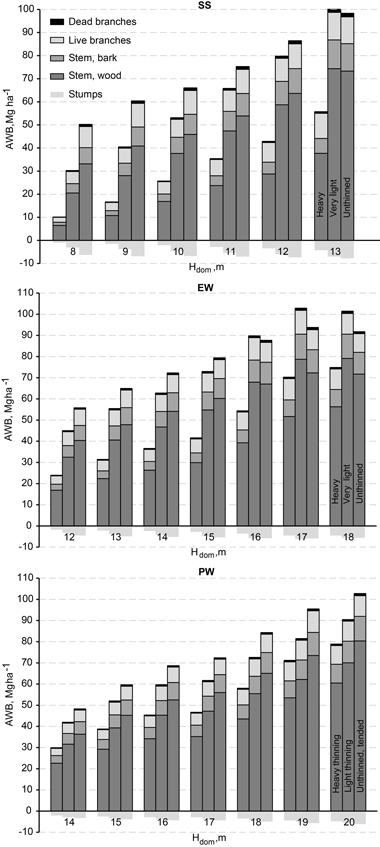
Fig. 5. Mean above-ground woody biomass (AWB) development + stumps of growing stock in downy birch stands by dominant height and treatment in height classes: SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage. Dominant height was used as the covariate in Eq. 1, and the models were formulated, and the biomass estimates calculated, through consideration of the mean dominant height separately for each height class at each measurement.
Most of the above-ground leafless biomass was accounted for by stem wood (65–77%) increasing clearly with stand age. The percentage of stem bark was 12%, regardless of age or treatment. Compared to the above-ground woody biomass, the relative amount of stumps was also stable, 7%. The proportion of branches was 11–23%, decreasing with stand age and density.
The amount of above-ground woody biomass (AWB) in growing stock is an interesting variable from the perspective of harvestable wood potential. AWB showed a straight linear positive correlation with dominant height (Fig. 5) in light thinning, but it curved downwards for heavy thinning and slightly upwards for unthinned control. The differences in AWB related to thinning intensity remained almost stable throughout the study period, except for very light thinnings in SS and EW height classes, for which the growth was faster and the biomass of the growing stock exceeded that in the unthinned control. In all three height classes, the mean of live above-ground biomass reached 100 tons ha–1 in dense plots, but culmination was seen only in EW.
The increment of branch biomass is important from the standpoint of whole-tree harvesting – e.g., for energy-wood production. The variation in live-branch biomass on the plots at the time of establishment of the experiments was 1.5–8 Mg ha–1 for SS height class, 3–7 Mg ha–1 for EW, and 4–6 Mg ha–1 for PW and correlated with thinning intensity. Around 25 years after thinning, the biomass of live branches culminated at a level of 9–11 Mg ha–1 and differences between stands of different densities were not significant anymore. In the SS height class, culmination was not yet reached at the end of the study period. For class EW, this culmination occurred at a dominant height of 17 m and for PW at 19 m.
Stem wood was the largest biomass component of growing stock, at 65–78% of AWB, with the proportion continuously increasing, while the share of stem bark (11–13%) and of stumps (6–10%, not included in AWB) decreased slightly with increasing age and thinning grade. The percentage of branches (10–23%) decreased with age, but it was the higher the heavier thinning was. According to the stem volume and biomass models used (Repola 2008), the mean wood density of downy birch stems with bark 15 years after first thinning was 0.486 g cm–1 (stdev. 0.026), well compatible with results of Hakkila (1982).
3.3 Total yield, removal in thinnings, and mortality
Total production of woody biomass including mortality increased with density. As an exception, the highest values were achieved on very lightly thinned plots, but the differences between these and the unthinned control were not significant. Figures for above-ground woody biomass production that include both growing stock and removal in thinnings are important from the energy-wood angle (Fig. 6). Until the end of the study period, they were 90–130 Mg ha–1, depending on treatment and height class. For thinned stands, the figure increased with stand density in SS and EW height classes, but on unthinned control plots the amounts of this harvestable biomass were, respectively, 24 Mg ha–1 and 27 Mg ha–1 lower than in very light thinning. In PW height class, the influence of the first thinning was not significant, however, without the removal during tending. The maximum mean annual AWB production reached was 2.5 Mg ha–1 and 2.6 Mg ha–1 for SS and EW height classes respectively.
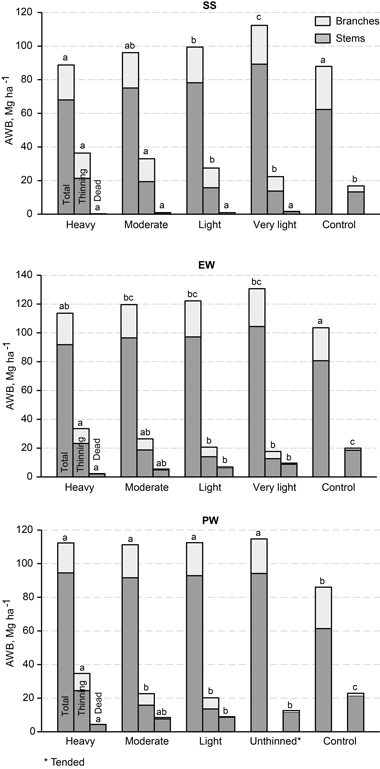
Fig. 6. Estimates of above-ground woody biomass (AWB) production until the end of the study period for different initial thinning intensities in height classes: SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage. The total AWB figures include live trees in the final growing stock and in first-thinning removal. Treatments denoted with the same letter (within a height class) are not significantly different (p > 0.05)). Site index, H50, and dominant height at final measurement were used as the covariates in Eq. 1 for total production, and their mean values for each height class (SS: H50 = 14.1 and Hdom = 12.9 m, EW: H50 = 17.2 and Hdom = 17.6 m, PW: H50 = 15.4 and Hdom = 19.0 m) were used in calculation of the estimated means. No covariate was used in models of the removal in thinnings, but the length of the study period was considered in the modelling of mortality: mean period 25 years for EW and PW, 22 years for SS.
The stem and branch biomass of first-thinning removal was 18–36 Mg ha–1, depending on thinning intensity, in all three height classes. The mortality of stem and branch biomass in the course of the study period was very low in heavy thinning but increased with stand density, reaching about 20 Mg ha–1 on unthinned plots.
The maximum mean total stem volume yield with bark, nearly 250 m3 ha–1, was reached with a density of 2000–2500 stems ha–1 in EW and PW height classes, but the difference from the unthinned control was not significant (Fig. 7). The maximum mean annual stem volume yield seen was 4.4 m3 ha–1 a–1 in EW. The stem volume of first-thinning removal was 35–60 m3 ha–1, depending on thinning intensity, in SS and EW height classes but it was 10 m3 higher in PW. The mortality of stem volume as a mean of unthinned control plots in EW and PW height classes during 25 years was 41 m3 ha–1, more than half being small-sized round-wood.
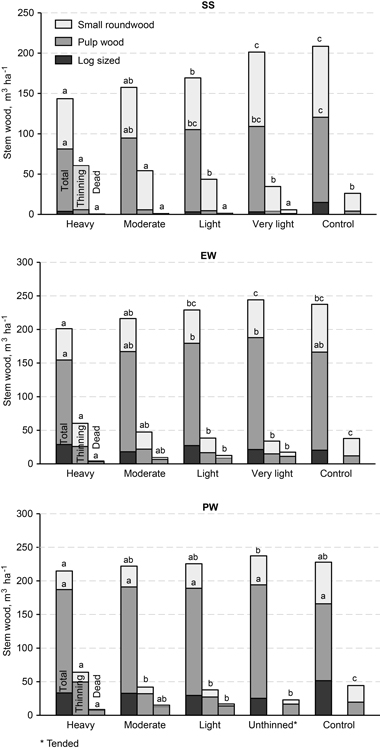
Fig. 7. Estimates of stem wood production until the end of the study period for different initial thinning intensities in height classes: SS = sapling stand stage, EW = energy wood thinning stage, and PW = pulp wood thinning stage. The stem wood was subdivided in to veneer log sized (> 18 cm), pulpwood and small round wood (< 6.5 cm). The total figures include live trees in the final growing stock and in first-thinning removal. For the model used, covariates, estimated means and statistical significance, refer to Fig. 6.
Traditionally, merchantable birch wood has consisted of pulpwood and veneer timber. However, the yield of veneer or sawn timber is very low and occasional in downy birch stands on peatlands because of the small stem size and low quality (Verkasalo 1991). That is why the production of stem volume was separated into three timber assortments: veneer log-sized material (diameter > 18 cm), pulpwood-sized material and small-sized round-wood (diameter > 6.5 cm). Volume production of small-sized stem wood excluding mortality in heavily, moderately and lightly thinned plots was 28–36 m3 ha–1 for PW, 47–50 m3 ha–1 for EW and, 61–64 m3 ha–1 for SS. In unthinned control it was the highest, 60–90 m3 ha–1 for PW, EW and SS respectively.
In PW height class, the production of merchantable stem volume (thinnings + final growing stock) totalled 190 m3 ha–1, on average, and was not dependent on the thinning intensity (Fig. 7). Mean annual yield was 2.5 m3 ha–1 over quite a long rotation, 75 years. In EW height class, the highest value for production of merchantable stem volume was 188 m3 ha–1, with an annual mean of 3.4 m3, on very lightly thinned plots. In SS height class, the yield of merchantable wood was highest in the unthinned control until the final measurement. However, the differences between cases of light or very light thinnings and the unthinned control were not statistically significant for any height class.
In the course of the rotation of 46–55 years the mean annual above-ground woody biomass increment (MAIAWB) was highest, 2.5 Mg ha–1 a–1, in unthinned and very lightly thinned stands. Excluding mortality, the net increment was 2.4 Mg ha–1 a–1 for very lightly thinned stands and 2.1 Mg ha–1 a–1 for unthinned ones. The mean annual stem volume increment (MAI) was highest in the experiments started at a young age (SS and EW), 4.3–4.5 m3 ha–1 a–1 for unthinned and very lightly thinned downy birch stands. Excluding the mortality, the highest annual net increment was 4.1 m3 ha–1 a–1 in lightly thinned stands, and 3.8 m3 ha–1 a–1 in unthinned ones. The mortality in dense stands accelerated toward the end of the study period. The cumulative sum of dead stem volume with bark was 40 m3 ha–1 in unthinned stands and 17 m3 ha–1 in very lightly thinned stands. For dead above-ground woody biomass, the equivalent numbers were 20–25 and 9–12 Mg ha–1, respectively.
The highest mean annual increment for merchantable wood (MAImerch) was 3.2 m3 ha–1 a–1 in very lightly thinned downy birch stands at a mean age of 55 years (EW). Against the hypothesis, the pulpwood production of younger stands (SS) was highest in unthinned thickets, 2.5 m3 ha–1 a–1, at a mean age of 46 years.
3.4 The effect of second thinning
The effect of second thinning could be analysed in plots within the energy-wood (EW) and pulpwood (PW) height classes that had previously been thinned either lightly or very lightly, with removal of half of the total stem number. This second thinning increased annual diameter growth at tree level significantly, but no influence on stem volume increment per hectare was detected (see Table 4). However, when mortality was reduced, the net stem volume increment increased in the PW height class after the second thinning, by 0.85 m3 ha–1 a–1. At the end of the study period, 10–20 years after the second thinning, the mean volume of the 600 thickest stems per hectare was higher by 46 dm3 (23%) for PW than in plots without second thinning. In the EW height class, these two influences were not significant. On average, live crowns at the end of the study period were 1.7 m longer on twice-thinned plots for both EW and PW.
| Table 4. The influence of second thinning on different growth and yield characteristics according to Figs. 2–7 and Tables 2–3. | ||||||
| Characteristic | Unit | EW stage | p- value | PW stage | p- value | Reference |
| Mean volume of 600 thickest trees ha–1 | dm3 | +14 | 0.16 | +46 | 0.00 | Fig. 3 |
| Mean crown length at the end of study | m | +1.7 | 0.00 | +1.7 | 0.00 | Fig. 2 |
| Annual diameter increment | mm a–1 | +0.89 | 0.00 | +0.71 | 0.00 | Tab. 2 |
| Annual stem volume increment | m3 ha–1 a–1 | −0.24 | 0.67 | −0.06 | 0.85 | Tab. 3 |
| Net stem volume increment | m3 ha–1 a–1 | +0.55 | 0.35 | +0.85 | 0.04 | Fig. 4 |
| Total AWB-production | Mg ha–1 | −3.8 | 0.41 | −4.1 | 0.16 | Fig. 5 |
| Thinning outturn of AWB | Mg ha–1 | +19.2 | 0.00 | +21.6 | 0.00 | Fig. 5 |
| Live AWB-production | Mg ha–1 | +3.8 | 0.44 | +6.5 | 0.03 | Fig. 5 |
| Mortality of AWB | Mg ha–1 | −6.7 | 0.00 | −7.5 | 0.00 | Fig. 5 |
| Production of branches | Mg ha–1 | +2.6 | 0.00 | +2.1 | 0.00 | Fig. 5 |
| Thinning outturn of stem volume | m3 ha–1 | +33.4 | 0.00 | +38.6 | 0.00 | Fig. 6 |
| Mortality of stem volume | m3 ha–1 | −11.3 | 0.00 | −12.8 | 0.00 | Fig. 6 |
| Total stem wood production | m3 ha–1 | −3.3 | 0.69 | −14.0 | 0.02 | Fig. 6 |
| Thinning outturn of pulpwood | m3 ha–1 | +18.4 | 0.00 | +32.5 | 0.00 | Fig. 7 |
| Mortality of pulpwood-sized stems | m3 ha–1 | −6.5 | 0.00 | −10.5 | 0.00 | Fig. 7 |
| Production of commercial wood | m3 ha–1 | +1.7 | 0.83 | −2.2 | 0.67 | Fig. 7 |
| Production of small-sized roundwood | m3 ha–1 | +3.0 | 0.00 | +3.0 | 0.00 | Fig. 7 |
In the second thinning, the thinning outturn for above-ground woody biomass (AWB) and stem volume was nearly the same in the EW and PW height classes, but for pulpwood in the PW class it was almost double the amount seen in EW. In the final measurement, the influence of the second thinning in reduction of standing AWB was still 16 Mg ha–1 for the EW height class and 20 Mg ha–1 for PW. This shows that the second thinning had no significant influence on total production of AWB, or even on production of commercial wood, although mortality was decreased. An exception was found in the total production of stem volume, which in the PW class was 14 m3 ha–1 lower because of second thinning though not significantly different within EW. Instead, the production of small-sized stem wood was increased, because a large amount of this was cut in the second thinning and would have ended up dead if this thinning had not occurred.
4 Discussion
4.1 Stand density and self-thinning
The stem number for growing stock varied over a wide range in this study as compared to most earlier studies of thinning. The benefit of high density was evident especially from the mortality and biomass-production angle. The decrease in stem number in unthinned stands as a function of dominant height was observed to be rectilinear on a logarithmic scale, from 16 000 ha–1 to 3000 ha–1 when the dominant height increased from 10 to 18 m (see Fig. 1). Similar results were obtained by Björklund and Ferm (1982) for downy birch on peatland and Johansson (1999) for silver and downy birch on formerly arable land. The number of live trees in naturally regenerated downy birch thickets in a fertilised peat cut-away area followed the same trend (Hytönen et al. 2011), but in young stands at 7 m dominant height it was 70% above the value seen in the present study though equal beyond a dominant height of 12 m. A difference of this nature was observed by Ferm and Kaunisto (1983), too, for a peat cut-away area, and by Rytter and Werner (2007) for birch-dominated mineral soils in southern Sweden.
Lower densities than found in this study at corresponding dominant heights were found in unmanaged naturally regenerated birch stands in Finland (Koivisto 1959) and in Ireland (Nieuwenhuis and Barret 2002) and for coppicing stands of downy birch on drained peatlands and on wet mineral soils (Ferm 1990a). The mortality of birch in two peat cut-away areas was higher than in our results for a plantation over 19 years from planting (Hytönen and Saarsalmi 2009) and in a birch-dominated coppicing stand with Salix spp. between five- and eight-metre dominant height, especially for a silver birch mixture (Hytönen and Aro 2010 and 2012). On the other hand, fertilisation treatments (NPK, PK, or wood ash) did not significantly affect stand density on these two sites, although the increase in height was greater.
In this study, the basal area of unthinned young and mid-aged downy birch stands rose to 40 m2 ha–1 or even more, increasing the more northern the stand. In connection with this result, the self-thinning was slower than reported earlier for downy birch (Jutras et al. 2003) or other broadleaf species (Hynynen 1993; Eid and Tuhus 2001; Eid and Øyen 2003). The maximum basal area for silver birch has been considered to be roughly 30 m2 ha–1 (Hynynen 1993) or below (Koivisto 1959). The equivalent figure for downy birch has been taken to be 32–33 m2 ha–1 or a little higher (Saramäki 1977; Nieuwenhuis and Barret 2002).
According to Jutras et al. (2003), increasing temperature sum is linked to higher mortality for downy birch stands. This result was surmised to be connected with harsher competition in more southerly locations. The similar result obtained in this study runs counter to findings consistent with general opinion that higher stand density can be maintained on better site types (Vanclay 1994; Mäkinen and Isomäki 2004a and 2004b). Our result may be associated with some shade-tree characteristics of downy birch (Ferm 1990b) and relatively low height increment in northern locations in spite of good site types. However, downy birch may just be better adapted in Northern conditions. In addition to that of the temperature sum, the effect of latitude with different light conditions on self-thinning should be studied.
4.2 Crown development
The base of the living crown rose rapidly in unthinned and lightly thinned downy birch stands, but the crown ratio fell more slowly and seldom to below 40%, because the trees with the smallest crowns were first to die. In fact, the average crown length of living downy birches increased even in unthinned stands as age increased, but this occurred more slowly than in thinned stands (see Kauppi et al. 1988). Mortality and reduction in crown ratio have been found to show a sharper increase in dense silver birch stands (Niemistö 1995; Hynynen 1993) than was shown for the similar downy birch stands in this study.
According to Rytter and Werner (2007) and Zalitis and Zalitis (2007), the early thinning of broadleaf stands is important from the standpoint of crown development and future growth’s influence on the production of valuable timber. So did we start the thinning too late in our experiments, and was this the reason for the low thinning response? The live crowns of birch should be over 50% of tree height (Niemistö 1991). At the outset of the sapling-stand thinning (SS), the crown ratio of all trees was more than 50% and the figure was above 60% for growing stock after thinning. At the end of the study period, the crown ratio was 70% and 65%, respectively, for heavy and moderate thinning, indicating that thinning was not begun too late. In the trials that began in the energy-wood and pulpwood thinning stage, respectively, the initial crown ratio for growing stock was 55% and 50%. That is why it appears that the first thinning may have taken place a little too late. However, the crown ratio on those plots thinned heavily or moderately saw a 5–10-percentage-point increase in the course of the study period.
4.3 Growth response
In the course of the first 15 years after first thinning, the mean diameter growth increased with thinning intensity in all three height classes (see Table 2). The influence on total increase in volume (m3 ha–1) was the opposite over the first 15 years, but no influence was observed later (see Fig. 4). Later (after 15 years), the positive effect of thinning on mean diameter growth was seen only for the youngest stands thinned in the sapling stand stage SS or for heavy thinning only in the energy-wood thinning stage EW.
The increase in diameter of the 600 thickest stems per hectare was accelerated by thinning of young birch stands (SS and EW) but not of mature stands (PW) (see Fig. 3). On the other hand, the second thinning accelerated diameter growth in mature stands but had no influence on the total increase in volume (m3 ha–1); meanwhile, the net volume increment was higher after the second thinning, because of reduced mortality. The net volume increment was highest for very light thinning of younger stands (SS and EW; see Fig. 4).
The average stem volume increment for unthinned stands in 15 years from the start of the experiments was 5.7 to 5.9 m3 ha–1 a–1 in all three height classes (see Table 3). Light, moderate, and heavy thinning decreased volume growth by 10–26%, 12–41%, and 29–50%, respectively, with the low end of the range indicating the mean for the mature stands (PW) and the upper bound representing the youngest stands (SS). The volume increment in this study was 70–80% of that found with mixtures of Norway spruce and downy birch described in the model of Repola et al. (2006). In these mixtures, the decrease in volume growth caused by the first thinning was approximately half of that measured in the present study. When the proportion of birch was set to 90–100% in the model, the thinning response and even the volume increment were almost equal to those seen in this study. Increased diameter growth due to the thinning was found only for spruce, not for birch. The mortality of downy birch was higher in mixed stands than the pure birch stands considered in this study.
According to Kojola et al. (2004), the thinning response of Scots pine on drained peatlands was as low as in this study with similar first thinning stages (PW) and the same climate conditions. In contrast for downy birch in the present study, the thinning response in the pine stands with a high level of initial stocking was greater. On upland sites in southern Finland, the volume increment for Scots pine after heavy thinning was 75% of that seen for unthinned stands (Mäkinen and Isomäki 2004a). On fertile upland sites in more southerly locations, clearly smaller reductions in the volume increment caused by heavy thinning have been reported for spruce (Mäkinen and Isomäki 2004b; Eriksson and Karlsson 1997) and for planted silver birch (Niemistö 2008) in the 15 years after first thinning. The effect of the number of stems retained appeared to be more important in downy birch stands on peatlands (in the present study) than in earlier studies on upland sites for silver birch and Norway spruce, and even for spruce or mixed stands on peatlands. Scots pine lies somewhere between downy birch and spruce in this respect.
The thinning response for downy birch was greater in mature stands (PW and after second thinning) than in younger stands after first thinning (SS and EW). This agrees with the findings of Niemistö (1991) and of Niemistö and Poutiainen (2004), while the result runs counter to those of earlier studies, which showed greater thinning response at young ages (for broadleafs, see Juodvalkis et al. 2005; for mixed stands, see Repola et al. 2006; for spruce stands, see Mäkinen and Isomäki 2004b; for spruce shelter-wood cuttings, see Holgén 2003). In this respect, birch and spruce seem to be quite different species because of their shadow-tolerance and different growth rhythm through the rotation.
4.4 Yield potential
In spite of the northerly location of the experiments in this study, the yield of stem volume with bark (MAI: 4 m3 ha–1 a–1) in dense birch stands on peatlands was rather high in comparison to naturally normal birch stands on upland sites in the southern half of Finland (Koivisto 1959): 4.1 and 4.9 m3 ha–1 a–1 for Myrtillus and Oxalis-Myrtillus site types, respectively. However, the stem number was lower in those stands on upland sites. The highest mean annual increment for merchantable wood (MAImerch: live trees, d > 6.5 cm) in this study was 3.2 m3 ha–1 a–1 in very lightly thinned downy birch stands. For dense Scots pine stands with the same climatic conditions but with longer rotation on slightly lower-production peatland sites (Kojola et al. 2004), both MAItot (2.3–3.9 m3 ha–1 a–1) and MAImerch (1.7–2.5 m3 ha–1 a–1) were lower than for the downy birch in this study.
The mean annual AWB increment was also highest, at 2.5 Mg ha–1 a–1, in unthinned and very lightly thinned dense birch thickets. A similar or higher yield inclusive of dead trees (2.5–5.3 Mg ha–1 a–1) has been reported in naturally regenerated, young (7–23 years) birch thickets in Finland (Björklund and Ferm 1982; Ferm 1990b). Renou-Wilson et al. (2010) estimated yields of 3.1–5.8 Mg ha–1 a–1 for naturally regenerated birch stands in Ireland, and Johansson (1999) found a corresponding 0.5–4.4 Mg ha–1 a–1 on previously arable land in Sweden. In the peat cut-away areas, MAIAWB inclusive of dead trees has been found to be higher than in this study: 2.9–3.2 Mg ha–1 a–1 over 15–26 years (Hytönen et al. 2011) for naturally regenerated downy birch stands and 3.0 Mg ha–1 a–1 over 21 years for unfertilised coppice stands (Hytönen and Aro 2012), even reaching 4.1–4.6 Mg ha–1 a–1 for fertilised ones. In silver-birch-dominated peat cut-away areas, biomass production has reached still higher figures, 4.4 Mg ha–1 a–1 over 16 years (Ferm and Kaunisto 1983), 5.7 Mg ha–1 a–1 over 37 years (Hytönen and Aro 2012), and 5.7–5.9 Mg ha–1 a–1 over 19 years on planted (with 20 000 seedlings per hectare) and intensively fertilised plots (Hytönen and Saarsalmi 2009). All of the peat cut-away areas had locations equivalent to those of the southernmost stands in this study.
Heavy thinning in young stands (here SS and EW) reduced AWB yield by 20% (see Fig. 6), but second thinning of denser stands did not affect it. For the mature stands (PW), the first thinning had no influence on live-biomass production but the second thinning increased it by 6.5 Mg ha–1 during the study period. The variation in the biomass of growing tree stands caused by thinning in this study diminished in the course of time, but significant differences were still present at the end of the study period. The relative increase in biomass after heavy thinning was greatest for branches, whose volume approached the level seen in denser stands after 15 years. Stem wood was the largest biomass component, at 65–78% of AWB, with the proportion continuously increasing, while the share of stem bark (11–13%) and of stumps (6–10%) decreased slightly with increasing age and thinning grade. The percentage of branches (10–23%) decreased with age, but it was the higher the heavier thinning was.
4.5 Thinning practices
Throughout the history of thinning research, there has been discussion of thinning’s positive and negative influences on the total or merchantable yield of a tree stand (Zeide 2001). Generally, thinning, mostly from below, is thought to increase the merchantable but not the total volume increment of trees per unit area. Within a wide range of density, total production is thought to be independent of thinning intensity. In the present study, similar results were found for total production in the pulpwood-thinning stage only, not prior to that, and not for merchantable volume at all. One reason for this result might be, in addition to tree species itself, the rising of the groundwater level after thinning, followed by decreased canopy interception and transpiration (Sarkkola et al. 2010; Koivusalo et al. 2011), which, in turn, can result in a reduced volume increment on peatland sites (Päivänen and Sarkkola 2000).
At what density the merchantable increment reaches its maximum for a given species, site, and age is at the heart of the question. In this study, the attention is focused in downy birch stands on fertile peatlands. Because of the extraordinarily wide stand-density range and the long study period for our permanent plots, it should be possible to determine the culmination density for the yield of pulpwood or merchantable biomass. In the trials established with sapling stands, maximum biomass production was reached with very light thinning (5000–7000 stems ha–1). The difference from the unthinned control was not significant for total biomass inclusive of mortality, but it was very significant for the yield of living above-ground biomass. On the contrary, the total production of stem volume or pulpwood was highest on unthinned plots in the course of 20–30 years after thinning of sapling stands. This result does not cover the whole rotation, but considerable changes in stand development should take place if the result for stem volume is to be changed.
In the trials established at the energy-wood thinning stage, the maximum biomass or stem volume production, including or excluding mortality, was reached with very light thinning (2500–3000 stems ha–1). The same result was obtained for pulpwood yield. However, differences from the unthinned control (4000–10 000 stems ha–1) or light thinning (1700–2200 stems ha–1) were not significant. Also, the second thinning did not show a significant influence on the total or merchantable yield of biomass or stem volume in these young stands. The density associated with the highest merchantable yield depends also on the minimum size of the commercial bole, which was quite small for birch pulpwood. Production of merchantable biomass is not limited by tree size, but it can be diminished by mortality in very dense stands.
The trials begun in the pulpwood thinning stage were an exception, because the light thinning took place at the sapling stage. The outturn of this early thinning could not be included in the plots’ total yield but could safely be assumed to be the same with all treatments. The total production of both biomass and stem volume was highest on unthinned plots. The yield of living biomass and commercially exploitable pulpwood did not differ between thinning treatments. The second thinning decreased total production of biomass and stem volume slightly, but a significant difference was found only for stem volume. On the other hand, the second thinning had a positive influence on the yield of living above-ground woody biomass but not on that of pulpwood.
From the results, it seems advantageous to produce energy wood alone or together with pulpwood on either unthinned or very lightly thinned downy birch stands. The growth in very dense downy birch stands remained at an acceptable level to the age of 40–50 years. On the other hand, the mortality of birch on peatlands in northern and western Finland was not too high until this age. Rotation shorter than 40 years was not promising, as the MAI was still increasing (see also Hytönen and Issakainen 2001 and Hytönen and Aro 2012). Careful research should be carried out from an economic angle that considers not just the yield of woody biomass. The influence of stand density and stem size on the harvesting cost and revenue at the time of both commercial thinning and final cutting must be thoroughly examined, as should the cost of all tending or pre-commercial-thinning operations. On the other hand, thinning of young sapling stands can result in the new coppice being even denser, with sprouts almost equal in size to the trees in unthinned sapling stands.
The only financial benefit of tending or thinning of downy birch stands seems to be earlier incomes and lower harvesting cost, resulting from the larger stems removed in thinning and final cutting. There is not a higher increment or greater value of wood produced. Also speaking in favour of thinnings for downy birch stands are the benefits of management practices such as creation of two-storey stands and regeneration via undergrowth (Mård 1996; Valkonen and Valsta 2001; Niemistö and Poutiainen 2004; Niemistö et al. 2012). The yield of pulpwood and, especially, energy wood was fairly high on fertile peatlands, where the regeneration and growing of more valuable tree species is often problematic. The productivity of dense downy birch stands was higher than expected in the northernmost locations included in the study material.
Acknowledgements
The author wishes to thank Eero Poutiainen for his tenacious fieldwork and proficient data processing throughout the course of the study. In addition, I thank Kari Alatalo, Jorma Issakainen, Olavi Kohal, Tapani Orttenvuori, and all of the field staff for taking care of the experiments and fieldwork; Ari Ferm, Matti Oikarinen, and Jussi Saramäki for planning the experiments; and Jaakko Repola for help with the biomass calculations.
References
Assman E. (1970). The principles of forest yield study. Pergamon Press, Oxford. 506 p.
Atkinson C.J. (1984). Quantum flux density as a factor controlling the rate of growth, carbohydrate partitioning and wood structure of Betula pubescens seedlings. Annals of Botany 54: 397–411.
Björklund T., Ferm A. (1982). Pienikokoisen koivun ja harmaalepän biomassa ja tekniset ominaisuudet. Summary: Biomass and technical properties of small-sized birch and grey alder. Folia Forestalia 500. 37 p.
Braastad H. (1967). Produksjonstabeller for bjørk. Meddelelser fra det Norske Skogforsøksvesen 22: 265–365. [In Norwegian].
Cameron A.D., Dunham R.A., Petty J.A. (1995). The effects of heavy thinning on stem quality and timber properties of silver birch (Betula pendula Roth). Forestry 68(3): 275–285. http://dx.doi.org/10.1093/forestry/68.3.275.
Eid T., Tuhus E. (2001). Models for individual tree mortality in Norway. Forest Ecology and Management 154(1–2): 69–84. http://dx.doi.org/10.1016/S0378-1127(00)00634-4.
Eid T., Øyen B.H. (2003). Models for prediction of mortality in even-aged forest. Scandinavian Journal of Forest Research 18(1): 64–77. http://dx.doi.org/10.1080/02827581.2003.10383139.
Eriksson H., Karlsson K. (1997). Effects of different thinning and fertilization regimes on the development of Scots pine (Pinus sylvestris (L.)) and Norway spruce (Picea abies (L.) Karst.) stands in long-term silvicultural trials in Sweden. Swedish University of Agricultural Sciences, Department of Forest Yield Research, Report 42. 135 p. [In Swedish with English summary].
Ferm A. 1990a. Nuorten vesasyntyisten hieskoivikoiden kehitys ja lahoisuus turvemaalla. Summary: Development and decay of young Betula pubescens coppice stands on peatland. Folia Forestalia 744. 17 p.
Ferm A. 1990b. Coppicing, aboveground woody biomass production and nutritional aspects of birch with specific reference to Betula pubescens. Finnish Forest Research Institute, Research Reports 348. 35 p.
Ferm A. (1993). Birch production and utilization for energy. Biomass and Bioenergy 4(6): 391–404. http://dx.doi.org/10.1016/0961-9534(93)90061-8.
Ferm A., Kaunisto S. (1983). Luontaisesti syntyneiden koivumetsiköiden maanpäällinen lehdetön biomassa-tuotos entisellä turpeennostoalueella Kihniön Aitonevalla. Summary: Above-ground leafless biomass production of naturally generated birch stands in a peat cut-away area at Aitoneva, Kihniö. Folia Forestalia 558. 32 p.
Ferm A., Kauppi A. (1990). Coppicing as a means for increasing hardwood biomass production. Biomass 22: 107–121. http://dx.doi.org/10.1016/0144-4565(90)90010-H.
Fries J. (1964). Summary: Yield of Betula verrucosa Ehrh. in Middle Sweden and southern North Sweden. Studia Forestalia Suecica 14. 303 p.
Hakkila P. (1982). Wood density survey and dry weight tables for pine, spruce and birch stems in Finland. Communicationes Instituti Forestalis Fenniae 96(3). 59 p.
Heinonen J. (1994). Koealojen puu- ja puustotunnusten laskentaohjelma KPL, käyttöohje. Finnish Forest Research Institute, Research Reports 504. 80 p. [In Finnish].
Holgén P., Söderberg U., Hånell B. (2003). Diameter increment in Picea abies shelterwood stands in northern Sweden. Scandinavian Journal of Forest Research 18(2): 163–167. http://dx.doi.org/10.1080/02827580310003731.
Hultén E., Fries M. (1986). Atlas of North European vascular plants: north of the Tropic of Cancer. I. Introduction, Taxonomic Index to the Maps 1–996, Maps 1–996. Koeltz Scientific Books, Königstein. 498 p.
Hynynen J. (1993). Self-thinning models for even-aged stands of Pinus sylvestris, Picea abies and Betula pendula. Scandinavian Journal of Forest Research 8(3): 326–336. http://dx.doi.org/10.1080/02827589309382781.
Hynynen J., Niemistö P., Viherä-Aarnio A., Brunner A., Hein S., Velling P. (2010). Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83(1): 103–119. http://dx.doi.org/10.1093/forestry/cpp035.
Hytönen J., Aro L. (2010). Biomass production of birch on cut-away peatlands – energy wood with short rotation? In: Parrotta J.A., Carr M.A. (eds.). Forests for the future: sustaining society and the environment. XXIII IUFRO Congress, 23–28 August 2010, Seoul, Republic of Korea. Abstracts. The International Forestry Review 12(5): 173.
Hytönen J., Aro. L. (2012). Biomass and nutrition of naturally regenerated and coppiced birch on cutaway peatland during 37 years. Silva Fennica 46(3): 377–394.
Hytönen J., Issakainen J. (2001). Effect of repeated harvesting on biomass production and sprouting of Betula pubescens. Biomass Bioenergy 20(4): 237–245. http://dx.doi.org/10.1016/S0961-9534(00)00083-0.
Hytönen J., Saarsalmi A. (2009). Long-term biomass production and nutrient uptake of birch, alder and willow plantations on cut-away peatland. Biomass Bioenergy 33(9): 1197–1211. http://dx.doi.org/10.1016/j.biombioe.2009.05.014.
Hytönen J., Issakainen J., Vihanta S. (2011). Hieskoivun biomassatuotos suonpohjalla Limingan Hirvinevalla. [Project report in Finnish].
Johansson T. (1999). Biomass equations for determining fractions of pendula and pubescent birches growing on abandoned farmland and some practical implications. Biomass Bioenergy 16(3): 223–238. http://dx.doi.org/10.1016/S0961-9534(98)00075-0.
Juodvalkis A., Kairiukstis L., Vasiliauskas R. (2005). Effects of thinning on growth of six tree species in north-temperate forests of Lithuania. European Journal of Forest Research 124(3): 187–192. http://dx.doi.org/10.1007/s10342-005-0070-x.
Jutras S., Hökkä H., Alenius V., Salminen H. (2003). Modeling mortality of individual trees in drained peatland sites in Finland. Silva Fennica 37(2): 235–251.
Kauppi A., Kiviniitty M., Ferm A. (1988). Growth habits and crown architecture of Betula pubescens Ehrh. of seed and sprout origin. Canadian Journal of Forest Research 18(12): 1603–1613. http://dx.doi.org/10.1139/x88-244.
Keltikangas M., Seppälä K. (1977). Ojitusalueiden koivikoiden kasvatus taloudellisena vaihto-ehtona. Summary: The economics of growing birch stands on drained peatlands. Silva Fennica 11(1): 49–68.
Koivisto P. (1959). Kasvu- ja tuottotaulukoita. Summary: Growth and yield tables. Communicationes Instituti Forestalis Fenniae 51(8). 49 p.
Koivusalo H., Hökkä H., Lauren A. (2011). A small catchment scale approach for modeling effects of forest management on water cycle in boreal landscape. In: Bredemeier M., Cohen S., Godbold D.L., Lode E., Pichler V., Schleppi P. (eds.). Forest management and the water cycle – an ecosystem-based approach. Springer Verlag. p. 419–434.
Kojola S., Penttilä T., Laiho R. (2004). Impacts of different thinning regimes on the yield of uneven-structured Scots pine stands on drained peatland. Silva Fennica 38(4): 393–403.
Laasasenaho J. (1982). Taper curve and volume functions for pine, spruce and birch. Communicationes Instituti Forestalis Fenniae 108. 74 p.
Mäkinen H., Isomäki A. 2004a. Thinning intensity and growth of Scots pine stands in Finland. Forest Ecology and Management 201: 311–325. http://dx.doi.org/10.1016/j.foreco.2004.07.016.
Mäkinen H., Isomäki A. 2004b. Thinning intensity and growth of Norway spruce stands in Finland. Forestry 77(4): 349–364. http://dx.doi.org/10.1093/forestry/77.4.349.
Mård H. (1996). The influence of a birch shelter (Betula spp.) on the growth of young stands of Picea abies. Scandinavian Journal of Forest Research 11(4): 343–350. http://dx.doi.org/10.1080/02827589609382945.
Moilanen M. (1985). Lannoituksen ja harvennuksen vaikutus hieskoivun kasvuun ohutturpeisilla oji-te-tuilla rämeillä. Summary: The effect of fertilization and thinning on the growth of birch Betula pubescens on the drained mires with thin peat layer. Folia Forestalia 629. 29 p.
Näslund M. (1937). Skogsförsökanstaltens gallringförsök i tallskog. [The thinning experiments of the Forest Research Institute in Scots pine stand]. Meddelandet från Statens Skogsförsökanstalt 29(1). 169 p. [In Swedish with German summary].
Niemistö P. (1991). Hieskoivikoiden kasvatustiheys ja harvennusmallit Pohjois-Suomen turvemailla. Summary: Growing density and thinning models for Betula pubescens stands on peatlands in northern Finland. Folia Forestalia 782. 36 s.
Niemistö P. (1995). Influence of initial spacing and row-to-row distance on the crown and branch properties and taper of silver birch (Betula pendula). Scandinavian Journal of Forest Research 10(3): 235–244. http://dx.doi.org/10.1080/02827589509382889.
Niemistö P. (2008). Koivikon kasvatus. In: Niemistö P., Viherä-Aarnio A., Velling P., Heräjärvi H., Verkasalo E. (eds.). Koivun kasvatus ja käyttö. Metla and Metsäkustannus Oy. p. 96–136. [In Finnish].
Niemistö P., Korhonen K.T. (2008). Koivuvarat ja kasvu. In: Niemistö P., Viherä-Aarnio A., Velling P., Heräjärvi H., Verkasalo E. (eds.). Koivun kasvatus ja käyttö. Metla and Metsäkustannus Oy. p. 175–181. [In Finnish].
Niemistö P., Poutiainen E. (2004). Hieskoivikon käsittelyn vaikutus kuusialikasvoksen kehitykseen Keski- ja Pohjois-Pohjanmaan viljavilla ojitusalueilla. Metsätieteen aikakauskirja 4/2004: 441–459. [In Finnish].
Niemistö P., Korpunen H., Lauren, A, Salomäki M., Uusitalo J. (2012). Impacts and productivity of harvesting when retaining young understorey spruces in final cutting of downy birch (Betula pubescens). Silva Fennica 46(1): 81–97.
Nieuwenhuis M., Barrett F. (2002). The growth potential of downy birch (Betula pubescens (Ehrh.)) in Ireland. Forestry 75(1): 75– 87. http://dx.doi.org/10.1093/forestry/75.1.75.
Nygren M., Kellomäki S. (1983). Effect of shading on leaf structure and photosynthesis in young birches, Betula pendula Roth and B. pubescens Ehrh. Forest Ecology and Management 7: 119–132. http://dx.doi.org/10.1016/0378-1127(83)90024-5.
Päivänen J., Sarkkola S. (2000). The effect of thinning and ditch network maintenance on the water table level in Scots pine stand on peat soil. Suo 51(3): 131–138.
Renou-Wilson F., Pöllänen M., Byrne K., Wilson D., Farrell E.P. (2010). The potential of birch afforestation as an after-use option for industrial cutaway peatlands. Suo 61(3–4): 59–76.
Repola J. (2008). Biomass equations for birch in Finland. Silva Fennica 42(4): 605–624.
Repola J. (2009). Biomass equations for Scots pine and Norway spruce in Finland. Silva Fennica 43(4): 625–647.
Repola J., Hökkä H., Penttilä T. (2006). Thinning intensity and growth of mixed spruce-birch stands on drained peatlands in Finland. Silva Fennica 40(1): 83–99.
Rytter L., Werner M. (2007). Influence of early thinning in broadleaved stands on development of remaining stems. Scandinavian Journal of Forest Research 22: 198–210. http://dx.doi.org/10.1080/02827580701233494.
Saramäki, J. 1977. Development of white birch (Betula pubescens Ehrh.) stands on drained peatlands in Northern Central Finland. Communicationes Instituti Forestalis Fenniae 91(2). 59 p.
Sarkkola S., Hökkä H., Koivusalo H., Nieminen M., Ahti E., Päivänen J., Laine J. (2010). Role of tree stand evapotranspiration in maintaining satisfactory drainage conditions in drained peatlands. Canadian Journal of Forest Research 40: 1485–1496. http://dx.doi.org/10.1139/X10-084.
Valkonen S., Valsta L. (2001). Productivity and economics of mixed two-storied spruce and birch stands in Southern Finland simulated with empirical models. Forest Ecology and Management 140(2–3): 133–149. http://dx.doi.org/10.1016/S0378-1127(00)00321-2.
Vanclay J.K. 1994. Modelling forest growth and yield: applications to mixed tropical forests. CAB International, Wallingford, U.K. 312 p.
Verkasalo E. (1997). Hieskoivun laatu vaneripuuna. Summary: Quality of European white birch (Betula pubescens Ehrh.) for veneer and plywood. Finnish Forest Research Institute, Research Reports 632. 483 p. + app. 59 p.
Zalitis T., Zalitis P. (2007). Growth of young stands of silver birch (Betula pendula Roth) depending on pre-commercial thinning intensity. Baltic Forestry 13(1): 61–67.
Zeide B. (2001). Thinning and growth: a full turnaround. Journal of Forestry 99: 20–25.
Total of 59 references
