Māra Kitenberga,
Guntars Šņepsts,
Jānis Vuguls,
Didzis Elferts,
Ieva Jaunslaviete,
Āris Jansons 
Tree- and stand-scale factors shape the probability of wind damage to birch in hemiboreal forests
Kitenberga M., Šņepsts G., Vuguls J., Elferts D., Jaunslaviete I., Jansons Ā. (2021). Tree- and stand-scale factors shape the probability of wind damage to birch in hemiboreal forests. Silva Fennica vol. 55 no. 2 article id 10483. https://doi.org/10.14214/sf.10483
Highlights
- Prior stem and root damage significantly increased the probability of birch being wind-damaged
- Trees with a high slenderness ratio had a lower relative snapping height
- The snapping height was significantly negatively affected by the prior stem and root damage.
Abstract
Strong wind is the major natural disturbance in European forests, that periodically causes tremendous damages to forestry. Yet, factors that affect the probability of wind damage for birch (Betula pendula Roth and B. pubescens Ehrh.), the most common deciduous tree species in hemiboreal forests, are studied scarcely. This study aimed to assess the effects of several tree- and stand-scale variables on the probability of wind damage to birch using data from the Latvian National Forest Inventory (2004–2018), and determine individual tree characteristics that affect the height of the stem breakage. The data analysis was done using the Bayesian binary logistic generalized linear mixed-effects model and a linear mixed-effects model. The probability of wind damage significantly increased by stand age, basal area, and slenderness ratio. Trees with prior damage had a significantly higher probability (odds ratio 4.32) for wind damage. For wind-damaged trees, the snapping height was significantly decreased by an increase in the slenderness ratio (p = 0.03) and prior damage (p = 0.003). Previously damaged trees were more frequently (73%) snapped in the lowest 40% of tree height than trees without prior damage (54%). The probability of wind damage is largely set by factors related to the selection of site, species composition, and rotation. The damage probability could be decreased by management measures that lower competition within the stand with particular regard to preserving intact remaining trees during these manipulations. Factors that reduce the probability of the damage simultaneously increase the snapping height, emphasizing their relevance for mitigation of the wind damages.
Keywords
stem breakage;
Betula pendula;
snapping height;
stem damage;
stem fracture;
tree stability
-
Kitenberga,
Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia
 https://orcid.org/0000-0002-6192-988X
E-mail
mara.kitenberga@silava.lv
https://orcid.org/0000-0002-6192-988X
E-mail
mara.kitenberga@silava.lv
- Šņepsts, Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia E-mail guntars.snepsts@silava.lv
- Vuguls, Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia E-mail janis.vuguls@silava.lv
-
Elferts,
Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia; Faculty of Biology, University of Latvia, Jelgavas Street 1, LV–1004, Rīga, Latvia
https://orcid.org/0000-0002-9401-1231
E-mail
didzis.elferts@lu.lv
- Jaunslaviete, Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia E-mail ieva.jaunslaviete@silava.lv
-
Jansons,
Latvian State Forest Research Institute ‘Silava’, Rigas Street 111, LV–2169, Salaspils, Latvia
https://orcid.org/0000-0001-7981-4346
E-mail
aris.jansons@silava.lv
Received 22 November 2020 Accepted 30 April 2021 Published 11 May 2021
Views 109599
Available at https://doi.org/10.14214/sf.10483 | Download PDF

Supplementary Files
1 Introduction
In Europe, the expected gain in forest productivity driven by climate change might be adversely affected by an increase in the frequency and severity of natural disturbances (Reyer et al. 2017; Seidl et al. 2017). In Northern Europe, most strong windstorms occur during the cold seasons of autumn and winter (Laapas et al. 2019). Studies have revealed that, due to climate change, the amount of wind damage will be amplified by the decreasing period of frozen soil conditions in the cold season, when trees have better anchorage and a higher critical wind speed is needed to cause damage (Peltola et al. 1999, 2000; Kellomäki et al. 2008). The extent of wind damage is also expected to be shaped by climate-driven changes in vegetation structure and composition (Seidl et al. 2017).
In the future, the growth conditions of tree species in boreal and hemiboreal forests will be determined by species-specific responses to the altered climatic conditions. Studies based on global climate model projections have revealed that, in the southern boreal regions in Europe, the silver birch (Betula pendula Roth) will benefit most from climate change in terms of volume growth, yield, and carbon stock (Peltola et al. 2010; Alrahahleh et al. 2018). In hemiboreal forests, based on a tree-ring analysis, the silver birch had the highest tolerance to historic weather fluctuations compared to other tree species, such as the Scots pine (Pinus sylvestris L.), Norway spruce (Picea abies (L.) Karst.), and European beech (Fagus sylvatica L.) (Jansons et al. 2016). Studies have demonstrated high morphological plasticity of silver birch to acclimate growth to site and climatic conditions (Nicoll and Ray 1996; Rosenvald et al. 2011; Tenkanen et al. 2020). Although the predicted increase in air humidity under climate change might compromise the annual increment of silver birch (Sellin et al. 2017), in certain regions in Northern Europe, silver birch could be one of the optimal tree species to maintain forest productivity.
The probability of wind damage for deciduous tree species varies seasonally. The increase in deciduous stands in the future will consequently increase the amount of wind damage in the spring and summer when trees have leaves. However, in the autumn and winter when trees are leafless, susceptibility decreases due to reduced crown area (Peltola et al. 2010; Ikonen et al. 2020). The susceptibility to wind damage of an individual tree or stand changes over time and is related to forest development, management, and climatic and site conditions (Zeng et al. 2004; Heinonen et al. 2011; Snepsts et al. 2020). Based on tree-pulling tests, it is estimated that silver birch has a higher resistive bending moment for stem breakage than pine and spruce (Peltola et al. 2000). Hence, the admixture of silver birch can improve the overall wind stability of stands dominated by less wind-resistant tree species, such as the Norway spruce (Dhôte 2005; Valinger and Fridman 2011). Yet, a detailed analysis of the influence of tree- and stand-scale factors on the probability of wind damage for silver birch is lacking in hemiboreal forests. Such information could be used in wind-damage risk assessment, long-term carbon sequestration calculations, and strategic forest management planning.
Our previous research program focused explicitly on the most wind-susceptible species – Norway spruce – and aimed to assess the stability of the species using static pulling tests and Latvian National Forest Inventory (NFI) data. The main results of this project showed that non-lethal stem damage (bark-stripping) and root rot significantly reduced the stability of Norway spruce (Krisans et al. 2020; Snepsts et al. 2020). The reduction of windstorm damage in Norway spruce stands could be achieved by implementing adaptive silviculture measures, that is, by using improved planting material, decreased planting density, and adjusted timing of pre-commercial thinning (Samariks et al. 2020). Our current research program focuses on birch (B. pendula and B. pubescens Ehrh., not distinguished between the species), which, according to NFI, is the most widespread tree species occupying 27% of the total forest area. In recent years, forest regeneration by birch has been favored due to the low browsing damage, the possibility to regenerate stands naturally with minimal costs, and European Union subsidies that encourage creating mixed deciduous-coniferous stands. The increasing share of birch forests in Baltic countries and Fennoscandia (Kellomäki et al. 2008; Alrahahleh et al. 2018) highlights the importance of further knowledge to sustain forest productivity and resistance to wind damage.
This study aimed to assess the effects of a tree- and stand-scale factors on the probability of wind damage and the snapping height of birch. First, we hypothesized that the probability of wind damage is determined by stand age, growing stock, density, basal area, dominant species, recent management, and site type, as well as by prior damage and stem slenderness of individual trees. Further, we hypothesized that stem slenderness, prior damage, and height of the living crown affect the height of the stem breakage.
2 Materials and methods
The study was carried out in hemiboreal forests (Ahti et al. 1968) across Latvia, in North-Eastern Europe. We used the data of 14 749 repeatedly measured birch trees, collected in 3360 NFI plots between 2004 and 2018. Each tree has a unique identification number (ID) that allows researchers to follow the status of each tree throughout each re-measurement period of five years: 2004–2008, 2009–2013, and 2014–2018. In this study, we accounted for prior non-lethal damage and subsequent damage by wind. The prior damage was visually observed as non-lethal injuries of stem and roots that affect a cambium function and are caused by various damaging agents, e.g., wind, fire, animals, harvesting operations. The subsequent wind damage regards snapped and uprooted stems, whereas broken branches and leaning stems were not included in this study.
We followed tree status by combining two succeeding measurement periods: information of prior damage in the period of 2004–2008 was added to data of wind damage from the period of 2009–2013, and information of prior damage in the period of 2009–2013 was added to data of wind damage from the period of 2014–2018. Similarly, the proportion of wind-damaged trees in each respective period was calculated from the data collected during re-measurements in each combined period. The study design was based on the work of Snepsts et al. (2020) that assessed tree- and stand-scale effects on the probability of wind damage for Norway spruce.
The NFI design is based on a quadratic cluster of four permanent circular sampling plots (each 500 m2), established in a grid of 4 × 4 km. For a detailed description of NFI methodology, see Jansons and Līcīte (2010) and Silava (2013). Within each sample plot, diameter at breast height (DBH; ≥14 cm), height, and crown height is measured for each living tree, and a snapping height is measured for each wind-broken tree. For each measured tree, the occurrence of prior damage is recorded. Additionally, we calculated relative snapping height for each wind-damaged birch as a ratio between snapping height at the repeated measurement period and the height of the tree at the previous measurement period.
We also calculated two slenderness ratios for each tree to test their effect on the probability of birch being wind-damaged. First, we tested the commonly used height to DBH ratio (HD–1). Considering that under conditions where competition is not a limiting factor trees at a young age naturally tend to have higher slenderness than trees at later development stages (Rust 2014; Grabosky and Bassuk 2016) we also tested the effect of a quadratic-height to DBH ratio (H2D–1) to put a larger emphasis on tree height. The results revealed only HD–1 to be significant (Table 1), therefore the model using H2D–1 was rejected from the analysis. However, we have added the results of this model in a supplementary file (Supplementary file S1) as this slenderness measure showed a significant effect on spruce in our previous study (Snepsts et al. 2020).
| Table 1. Bayesian binary logistic generalized linear mixed-effect model estimates of tree- and stand-scale variables on the probability of wind damage for birch. For the variables with several levels, the estimate is the difference from the reference level. Data obtained from National Forest Inventory in Latvia. | ||||
| Variable | Estimate | Est.error | CI | |
| Min | Max | |||
| Intercept | −5.03 | 0.27 | −5.68 | −4.62 |
| Stand age (years)* | 0.39 | 0.07 | 0.26 | 0.53 |
| Dominant tree species (reference level: birch)*: | ||||
| Pine | −0.45 | 0.17 | −0.78 | −0.13 |
| Spruce | 0.08 | 0.16 | −0.25 | 0.39 |
| Black alder | 0.22 | 0.23 | −0.25 | 0.69 |
| Aspen | 0.17 | 0.21 | −0.24 | 0.57 |
| Grey alder | −0.13 | 0.45 | −1.08 | 0.69 |
| Oak | 0.08 | 0.77 | −1.54 | 1.47 |
| Ash | 0.06 | 0.70 | −1.42 | 1.31 |
| Goat willow | −0.39 | 1.31 | −3.48 | 1.71 |
| Stand density (trees ha–1)* | −0.70 | 0.11 | −0.92 | −0.50 |
| Overstory basal area (m2 ha–1)* | 0.21 | 0.07 | 0.08 | 0.34 |
| Management (reference level: not thinned in the last 5 years): | ||||
| Thinned in the last 5 years | 0.17 | 0.17 | −0.16 | 0.49 |
| Site type (reference level: dry mineral soil)*: | ||||
| Wet mineral soil | 0.00 | 0.20 | −0.41 | 0.40 |
| Peat soil | 0.75 | 0.20 | 0.36 | 1.14 |
| Drained wet mineral soil | 0.32 | 0.15 | 0.03 | 0.60 |
| Drained peat soil | 0.94 | 0.15 | 0.65 | 1.24 |
| Slenderness ratio* | 0.39 | 0.04 | 0.30 | 0.47 |
| Prior damage (reference level: not damaged)*: | ||||
| Damaged | 1.46 | 0.15 | 1.18 | 1.79 |
| Est.error – standard deviation of posterior distribution; CI – 95% credible interval; * – significant effect according to 95% credible interval; Birch – Betula pendula and B. pubescens; Pine – Pinus sylvestris; Spruce – Picea abies; Black alder – Alnus glutinosa; Aspen – Populus tremula; Grey alder – Alnus incana; Oak – Quercus robur; Ash – Fraxinus excelsior; Goat willow – Salix caprea | ||||
For each tree, the sample plot data were used to calculate respective stand parameters: standing volume (m3 ha–1), stand density (trees ha–1), basal area (m2 ha–1), and age (years). We grouped nationally classified (Bušs 1976) site types based on soil conditions: mesic and wet mineral soil (freely draining and periodically waterlogged mineral soil, respectively), peat soil (decomposed organic matter layer in ≥30 cm depth), and drained wet mineral- and peat- soil. We included sample plots with dominant common hemiboreal species: birch, Scots pine, Norway spruce, black alder (Alnus glutinosa (L.) Gaertn.), European aspen (Populus tremula L.), grey alder (A. incana (L.) Moench), pedunculate oak (Quercus robur L.), European ash (Fraxinus excelsior L.), and goat willow (Salix caprea L.).
To answer our first hypothesis, we assessed the effect of tree- and stand- scale variables (Table 2) on the probability of wind damage on birch by performing a Bayesian binary logistic generalized linear mixed effect model (GLMM). Tree ID and sampling plot ID were used as a nested random effect to consider the structure of the experiment. All continuous variables were scaled by dividing the values with the standard deviations to decrease data variation. Each set of four chains comprised 3500 iterations. R-hat values were used to assess the model performance, considering that values below 1.1 show converged model algorithm (Bürkner 2018). The effect of a variable was interpreted as being significant if its 95% credible interval did not include a value of zero. Then, non-linear hypothesis testing was used to compare the level of significant factors. The odds ratios were calculated to quantify associations (increased or decreased as compared to reference level) of the probability of wind damage between the levels of qualitative factors. The reference levels were: birch for the dominant species, dry mineral soils for the site type, absence of prior damage, and absence of management during the last five years.
| Table 2. The tested tree- and stand-scale variables used to determine the probability of wind damage on birch by Bayesian binary logistic generalized linear mixed. The number of observations is noted for categorical variables and mean ± standard deviation (SD) is noted for continuous variables. Data obtained from National Forest Inventory in Latvia. | ||||
| Variable | Description | Classes/ Range | Number of observations/ Mean ± SD | |
| 2004–2008 | 2009–2013 | |||
| Age | Stand age, years | 11–216 | 56.6 ± 20.9 | 58.1 ± 21.8 |
| Growing stock | Mean standing volume, m3 ha–1 | 16–850 | 270 ± 127 | 281 ± 136 |
| Species | Dominant tree species | Birch | 6639 | 7161 |
| Pine | 1825 | 1869 | ||
| Spruce | 1333 | 1334 | ||
| Black alder | 627 | 657 | ||
| Aspen | 793 | 787 | ||
| Grey alder | 398 | 341 | ||
| Oak | 47 | 47 | ||
| Ash | 63 | 25 | ||
| Goat willow | 36 | 52 | ||
| Stand density | Overstory number of trees, trees ha⁻¹) | 100–11 520 | 1145 ± 875 | 1112 ± 906 |
| Basal area | Overstory basal area, m2 ha–1 | 3–67 | 27.2 ± 9.9 | 27.8 ± 10.2 |
| Management | Harvesting operations in the last five years | Thinned | 1800 | 721 |
| Not thinned | 9974 | 11 571 | ||
| Site type | Site types grouped according to soil conditions | Dry mineral soil | 4718 | 4804 |
| Wet mineral soil | 1374 | 1370 | ||
| Peat soil | 1058 | 1345 | ||
| Drained mineral soil | 2462 | 2434 | ||
| Drained peat soil | 2162 | 2339 | ||
| Slenderness ratio | Height (m) to diameter at breast height (cm) ratio | 0.3–1.9 | 1.0 ± 0.2 | 1.0 ± 0.2 |
| Prior damage | Stem or root-stump damage in the previous measuring period | Not damaged | 11 303 | 11 572 |
| Damaged | 471 | 720 | ||
| Birch – Betula pendula and B. pubescens; Pine – Pinus sylvestris; Spruce – Picea abies; Black alder – Alnus glutinosa; Aspen – Populus tremula; Grey alder – Alnus incana; Oak – Quercus robur; Ash – Fraxinus excelsior; Goat willow – Salix caprea | ||||
A Bayesian GLMM was implemented to characterize the effect of incidence of prior damage on the wind-damage probability in the next period. In this model, only a sampling plot ID was used as a random effect.
To answer our second hypothesis on the effects of tree variables on the relative snapping height of wind-damaged trees, we used a linear mixed-effects model (LMER), applying a sampling plot ID as a random factor. The Variance Inflation Factor was used to assess the multicollinearity between independent variables, and the post-hoc test to assess significant differences between factor levels.
The data analysis was implemented using R 4.0.0 (R Core Team 2020) library ‘brms’ (Bürkner 2018) for Bayesian GLMM, and libraries ‘lme4’ (Bates et al. 2015), ‘lmerTest’ (Kuznetsova et al. 2017), and ‘emmeans’ (Lenth 2020) for LMER.
3 Results
During the combined periods of 2004–2008 to 2009–2013 and 2009–2013 to 2014–2018, 2.9% and 3.4% of all birch trees were wind-damaged, respectively. Among these, a significant portion (69% and 65%, respectively) were snapped, and about one-third (31% and 35%, respectively) were uprooted. The prior damage was a significant factor increasing the probability of wind-damage during the following measurement period (odds ratio 2.86; credible interval (CI)[1.74, 4.53]), as indicated by the combined tree status data. In the first combined period, 4.0% of birch trees initially had non-lethal damage; among these trees, 10.2% were wind-damaged at the time of re-measuring. Among birch trees without prior damage, only 2.6% of trees were wind-damaged. Similarly, in the second combined period, 5.9% of birch trees initially had non-lethal damage; among these trees, 9.3% were wind-damaged at the time of re-measuring. Among birch trees without prior damage, only 3.0% of trees were wind-damaged.
According to the GLMM analysis used to test our first hypothesis, the probability of wind damage to birch was significantly higher by increased slenderness ratio (Fig. 1), stand age (Fig. 2), and basal area as well as by decreased stand density. The presence of prior damage, site type, and dominant tree species were also significant factors with different effect strength among their categories (Table 1). The test of the nonlinear hypothesis indicated that the probability of wind damage was significantly higher for trees with prior damage (odds ratio 4.32; CI[3.261, 5.977]) than for undamaged trees (reference level). Trees growing on peat (odds ratio 2.12; CI[1.43, 3.12]), drained peat (odds ratio 2.56; CI[1.91, 3.47]), and drained wet mineral soils (odds ratio 1.37; CI[1.02, 1.82]) had a higher probability of damage than trees growing on mineral soils (reference level). Results of the same model indicated that birch has a significantly lower probability of wind damage in stands dominated by pine (odds ratio 0.63; CI[0.45, 0.88]) than in stands dominated by all other tested tree species: birch (reference level), spruce (odds ratio 1.07; CI[0.77, 1.47]), aspen (odds ratio 1.18; CI[0.78, 1.77]), black alder (odds ratio 1.24; CI[0.78, 1.98]), grey alder (odds ratio 0.87; CI[0.34, 1.99]), oak (odds ratio 1.08; CI[0.21, 4.35]), and goat willow (odds ratio 0.67; CI[0.03, 5.53]).
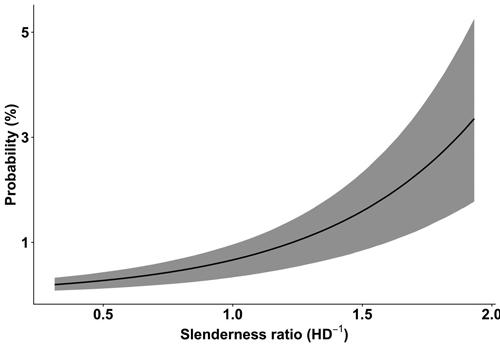
Fig. 1. The probability of wind damage to birch (Betula pendula and B. pubescens) in relation to the slenderness ratio (height (m) to the diameter at breast height (cm) ratio, HD–1), based on Latvian National Forest Inventory data. The 95% credible interval is represented by the grey area.
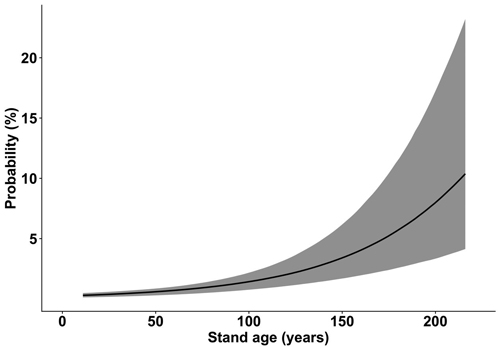
Fig. 2. The probability of wind damage to birch (Betula pendula and B. pubescens) in relation to the age of dominant tree species in a stand, based on Latvian National Forest Inventory data. The 95% credible interval is represented by the grey area.
The relative snapping height was significantly affected by two factors: the slenderness ratio HD–1 (F1423.3 = 4.74; p-value = 0.03) and prior damage (F1418.2 = 8.92; p-value = 0.003), as indicated by the LMER analysis used to test our second hypothesis. The snapping height gradually decreased with an increasing slenderness ratio (Fig. 3). Prior damage had a significant negative influence on the snapping height (Table 3). Trees with prior stem damage were considerably more frequently (73%) snapped in the first 40% of their height than undamaged trees (54%) (Fig. 4).
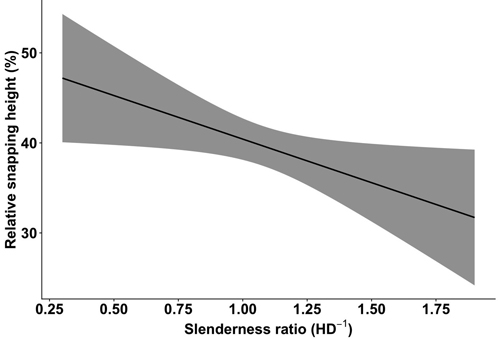
Fig. 3. The relative snapping height of birch (Betula pendula and B. pubescens) in relation to the slenderness ratio (height (m) to the diameter at breast height (cm) ratio, HD–1), based on Latvian National Forest Inventory data. The 95% confidence interval is represented by the grey area.
| Table 3. Generalized linear mixed-effects model estimates of slenderness ratio (HD–1), prior damage, and crown height (h-crown) effects on relative snapping height for birch. Data obtained from National Forest Inventory in Latvia. | ||||
| Variable | Estimate | SE | t-value | p-value |
| Intercept | 0.532 | 0.048 | 10.98 | <0.001 |
| HD–1 | −0.097 | 0.044 | −2.18 | 0.03 |
| h-crown (m) | 0.001 | 0.003 | −0.49 | 0.06 |
| Prior damage (Damaged vs Not damaged) | −0.090 | 0.030 | −2.98 | 0.003 |
| SE = standard error | ||||
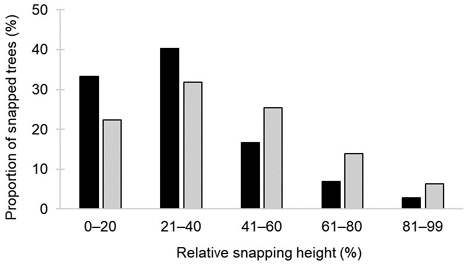
Fig. 4. The proportion of snapped birch trees (Betula pendula and B. pubescens) according to relative snapping height groups for previously damaged (black bars) and previously undamaged (light grey bars) trees, based on Latvian National Forest Inventory data.
4 Discussion
In this work, we summarized the effect of a tree- and stand-scale factors on the probability of wind damage to birch based on NFI sample plots in hemiboreal forests. In a previous study with a similar design, we assessed the probability of wind damage to Norway spruce in Latvia (Snepsts et al. 2020). Although there are some differences between the results of our current and former studies, several tree- and stand-scale factors appear to have a significant influence on both spruce and birch.
For birch, the probability of wind damage increased with increasing stand age and basal area (Table 1 and Fig. 2). The risk of wind damage is enhanced with stand development when increasing stand age is associated with an increase in height, basal area, and standing volume, and greater wind loading is received (Valinger and Fridman 2011; Donis et al. 2018). Suvanto et al. (2016) also found that increasing stand age and basal area are significant factors that increase the probability of wind damage following autumn storms and thunderstorms. For spruce, the only significant factor was stand age, not basal area (Snepsts et al. 2020). We assume that this difference could be due to statistical rather than ecological factors, although there are well-known ecological differences between broadleaves and conifers regarding wind damage probability.
For broadleaves, vulnerability to wind damage varies greatly between seasons (Ikonen et al. 2020). In our study, the structure of the NFI data precludes the attribution of the season when the damage occurred. In Northern Europe, the majority of strong winds occur during the autumn and winter seasons (Laapas et al. 2019) when the vulnerability of broadleaves is significantly lower, hence, they generally experience less damage than conifers (Peltola et al. 2010). The rooting systems also have substantial differences between species (Kalliokoski et al. 2008), and shallow rooting systems are one of the primary reasons why Norway spruce has a high susceptibility to wind damage. The modification of rotation length is one of the adaptive silviculture management options to minimize wind damage risks associated with increasing stand age. By shortening the rotation length, the period when trees are most vulnerable to wind damage can be reduced (Donis et al. 2020).
Trees with higher slenderness ratio values had a higher probability of wind damage (Fig. 1) and a lower relative snapping height (Fig. 4). Several studies have agreed that a lower critical wind speed is needed to damage trees with a high slenderness ratio (Päätalo et al. 1999; Peltola et al. 1999, 2000; Gardiner et al. 2000; Pukkala et al. 2016), hence, making stands with such trees more prone to wind damage. For spruce, however, a trend of increased susceptibility for trees with a lower slenderness ratio was found in our previous study (Snepsts et al. 2020). Similarly, spruce trees with a low slenderness ratio were found to have an increased probability of uprooting (Díaz-Yáñez et al. 2017) and pooled damage of uprooting, stem breakage, and leaning (Valinger and Fridman (1997). Other studies have found no difference in slenderness ratio between uprooted, snapped, and undamaged trees for Sitka spruce (Dunham and Cameron 2000) and Norway spruce (Ilisson et al. 2005). Overall, these studies indicate that conifers with conical crown architecture might have a different effect of slenderness ratio on snapping and uprooting than broadleaves.
The lower snapping height for trees with high slenderness ratio values (Fig. 3) suggests that the value of recovered timber would be considerably lower due to the higher probability of a damaged first cut log. A similar relationship between the relative snapping height and slenderness ratio has also been observed for spruce (Snepsts et al. 2020), suggesting that this association is similar between coniferous and broadleaved tree species in hemiboreal forests.
In general, the slenderness ratio is closely linked to the stand development and can be shaped by silvicultural practices (Valinger and Fridman 2011; Bošeľa et al. 2014). One of the adaptive management options to increase stand resistance to wind disturbances is thinning, which, in the long term, enhances the growth of above- and below-ground biomass. Following thinning, especially in young stands, trees acclimate to a higher wind load by enhancing their diameter growth, which decreases the stem taper (Peltola et al. 2007), and by changing the shape and allocation of their root system, which improves anchorage (Ruel et al. 2003).
The prior damage significantly increased the probability of wind damage and reduced the snapping height of wind-damaged trees (Tables 1 and 3); a similar observation was also reported for spruce (Snepsts et al. 2020). Studies have revealed that resistance to wind damage is significantly negatively affected by stem damage or defects (Peltola et al. 2000; Snepsts et al. 2020), which reduces the mechanical stability, wood strength, and/or vitality of the trees (Wood 1995; Peltola et al. 2000). Through the stem wounds, fungi can infect silver birch, often causing discoloration (Vasaitis et al. 2012) and occasionally leading to decay that negatively affects the mechanical properties of the wood (Terho et al. 2007). For spruce, stem wounds can be easily infected by decay-causing fungi (Burneviča et al. 2016), which can deteriorate wood after only a few years and significantly decrease the mechanical stability of the tree (Krisans et al. 2020). Infestation by decay-causing fungi is less likely for birch than for spruce.
Even without infestation stem wounds are likely to decrease the vitality of the tree, therefore increasing the probability of wind damage. Studies show that stem damage reduces the wood strength of the stem section which is close to it (Welch and Scott 2008), likely explaining the lower snapping height of the damaged birch (Fig. 4) and spruce (Snepsts et al. 2020) trees. In our study, most of the prior stem damage was located within the first two meters from the ground, due to either cervids or an anthropogenic origin. A higher proportion of prior-damaged trees that break within the first 20% of the tree height (Fig. 4) significantly decreases the value of the recovered timber because the first log, i.e. the most valuable assortment, is damaged (Welch and Scott 2008; Dubrovskis et al. 2018).
The dominant tree species had a significant effect on the probability of wind damage (Table 1). The probability of wind damage for birch was significantly lower in stands dominated by pine, corresponding with earlier studies showing that a slightly higher critical wind speed is needed to cause damage to pine than birch (Peltola et al. 2010; Donis et al. 2018). The positive effect of a pine admixture on lowering the probability of wind damage is also observed for spruce (Valinger and Fridman 2011; Snepsts et al. 2020). Several studies have shown that the admixture of wind firm tree species has a positive effect on the wind stability of a stand (Schelhaas 2008). The improved stability of birch and spruce in an admixture of pine could be attributed to deeper root systems, which form in response to competition (Schmid and Kazda 2001; Kalliokoski et al. 2008) and could enhance the wind stability of a stand. In addition, mixed stands are more likely to have heterogeneous stand structure that is less susceptible to uprooting damage (Díaz-Yáñez et al. 2017). Studies show that higher stand heterogeneity in birch-dominated stands (birch > 70% of basal area) decreases damage probability in comparison to spruce- and pine-dominated stands (Díaz-Yáñez et al. 2019). This suggests that, in addition to pine, both birch and pine admixtures improve overall stand stability. The NFI sampling design used in our study is not suitable to assess the influence of different proportions of admixture on the overall stand stability against wind disturbances.
The significantly higher probability of wind damage for birch in site types on peat, drained peat, and drained mineral soils could be linked to poorer root anchorage. Many studies have shown that root anchorage is one of the most important aspects determining vulnerability to wind damage (Nicoll et al. 2006; Peltola 2006; Fourcaud et al. 2008). Generally, lower critical wind speed is needed to damage trees on soils where root system development is restricted by a high groundwater level or other factors (Nicoll et al. 2006; Zubizarreta-Gerendiain et al. 2012). A higher probability of wind damage for birch in site types on drained soils could be linked to the condition of the drainage system and soil nutrition. In site types on drained soils in hemiboreal forests, the growth and vitality of trees can be hampered by a lack of soil mineral elements, such as potassium (Bārdule et al. 2009; Klavina et al. 2016). Tree growth on drained soils can be negatively affected by an unstable soil-water table, which can significantly fluctuate due to meteorological conditions, management practices, and the gradual deterioration of properly unmanaged ditch system (Jutras et al. 2006; Hökkä et al. 2008, 2012; Sikström and Hökkä 2016). We assume that trees might need to constantly adapt to changing site conditions, due to fluctuations in the groundwater table that could compromise their resistance to wind damage.
Studies that followed the windstorm ‘Gudrun’ in 2005 have revealed higher amounts of wind damage on peat soils than on mineral soils in hemiboreal forests (Donis et al. 2018). Contradicting results were obtained in boreal forests in Finland, where the probability of wind damage following windstorms was lower on organic soils than that on mineral soils (Suvanto et al. 2016). These contradicting results between boreal and hemiboreal forests could be linked to differences in the organic soil structure (the depth of the peat layer and the composition of the peat), groundwater level, and tree-species composition. In addition, the probability of wind damage is shaped by tree species-specific acclimatization to growth on peat soils (Nicoll and Ray 1996): in Finland, the probability of wind damage was assessed for spruce and pine (Suvanto et al. 2016), whereas in our study, it was assessed for birch.
Our data did not allow a separation between downy and silver birch; however, studies have shown that these two species differ in several morphological aspects related to wind resistance, such as wood anatomy. Downy birch prevails on moist and peat soils, whereas silver birch prevails on mineral soils (Hynynen et al. 2010). Thus, species could be one of the factors affecting wind resistance on different soil types. Unfortunately, no current studies on the differences in wind resistance exist between these two species.
Previous management within the last five years was not a significant factor in the model, a finding which resembles the results in a study of spruce (Snepsts et al. 2020). Several studies have shown that previous management is a significant factor, affecting the tree- and the stand-scale probability of wind damage (Zeng et al. 2004, 2006; Valinger and Fridman 2011; Donis et al. 2018). Additionally, information about tree cutting in NFI data is recorded within the sample plots, while information about management activities in neighboring stands is not, precluding a full examination of the potential influence of this factor on the probability of wind damage.
5 Conclusions
Our results reveal that the probability of wind damage for birch is shaped by several individual tree- and stand-scale factors, which are interconnected and change over stand development. Prior damage had a significant effect on both the probability of wind damage and the snapping height of wind-damaged trees. To increase the overall resistance of a stand to wind damage, trees with lower individual wind stability due to stem or root damage and high slenderness ratios should be removed during thinning or selective cuts. Alternatively, a management regime that minimizes the probability of mechanical damage to slender trees should be applied: lower initial spacing and few to no commercial thinning. The site type and tree-species admixture in the stand also had a significant influence; however, further studies are needed to obtain a deeper understanding of the individual and stand-scale resistance to wind damage before any practical recommendations could be proposed.
Acknowledgments
Study was carried out in Latvian Council of Science project “Reduction of wind storm damage risk in private forest management” (No lzp-2018/2-0349).
References
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in northwestern Europe. Ann Bot Fenn 5: 169–211.
Alrahahleh LA, Kilpeläinen A, Ikonen VP, Strandman H, Asikainen A, Venäläinen A, Kaurola J, Kangas J, Peltola H (2018) Effects of using certain tree species in forest regeneration on volume growth, timber yield, and carbon stock of boreal forests in Finland under different CMIP5 projections. Eur J For Res 137: 573–591. https://doi.org/10.1007/s10342-018-1126-z.
Bārdule A, Bāders E, Stola J, Lazdiņš A (2009) Latvijas meža augšņu īpašību raksturojums demonstrācijas projekta BioSoil rezultātu skatījumā. [Forest soil characteristic in Latvia according results of the demonstration project BioSoil]. Mežzinātne 20: 105–124.
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67: 1–48. https://doi.org/10.18637/jss.v067.i01.
Bošeľa M, Konôpka B, Šebeň V, Vladovič J, Tobin B (2014) Modelling height to diameter ratio—an opportunity to increase Norway spruce stand stability in the Western Carpathians. Cent Eur For J 60: 71–80. https://doi.org/10.2478/forj-2014-0007.
Bürkner PC (2018) Advanced Bayesian multilevel modeling with the R package brms. R J 10: 395–411. https://doi.org/10.32614/RJ-2018-017.
Burneviča N, Jansons Ā, Zaļuma A, Kļaviņa D, Jansons J, Gaitnieks T (2016) Fungi inhabiting bark stripping wounds made by large game on stems of Picea abies (L.) Karst. in Latvia. Balt For 22: 2–7.
Bušs K (1976) Latvijas PSR meža tipoloģijas pamati. [Fundamentals of forest classification in Latvia SSR]. LRZTIPI, Riga, Latvia
Dhôte J (2005) Implication of forest diversity in resistance to strong winds. In: Scherer-Lorenzen M, Körner C, Schulze ED (eds) Forest diversity and function. Temperate and Boreal Systems Ecological Studies 176. Springer, Berlin, Heidelberg, pp 291–307.
Díaz-Yáñez O, Mola-Yudego B, González-Olabarria JR, Pukkala T (2017) How does forest composition and structure affect the stability against wind and snow? Forest Ecol Manag 401: 215–222. https://doi.org/10.1016/j.foreco.2017.06.054.
Díaz-Yáñez O, Mola-Yudego B, González-Olabarria JR (2019) Modelling damage occurrence by snow and wind in forest ecosystems. Ecol Model 408, article id 108741. https://doi.org/10.1016/j.ecolmodel.2019.108741.
Donis J, Kitenberga M, Snepsts G, Dubrovskis E, Jansons A (2018) Factors affecting windstorm damage at the stand level in hemiboreal forests in Latvia: case study of 2005 winter storm. Silva Fenn 52, article id 10009. https://doi.org/10.14214/sf.10009.
Donis J, Saleniece R, Krisans O, Dubrovskis E, Kitenberga M, Jansons A (2020) A financial assessment of windstorm risks for Scots pine stands in hemiboreal forests. Forests 11, article id 566. https://doi.org/10.3390/F11050566.
Dubrovskis E, Donis J, Racenis E, Kitenberga M, Jansons A (2018) Wind-induced stem breakage height effect on potentially recovered timber value: case study of the Scots pine (Pinus sylvestris L.) in Latvia. For Stud 69: 24–32. https://doi.org/10.2478/fsmu-2018-0009.
Dunham RA, Cameron AD (2000) Crown, stem and wood properties of wind-damaged and undamaged Sitka spruce. Forest Ecol Manag 135: 73–81. https://doi.org/10.1016/S0378-1127(00)00299-1.
Fourcaud T, Ji JN, Zhang ZQ, Stokes A (2008) Understanding the impact of root morphology on overturning mechanisms: a modelling approach. Ann Bot 101: 1267–1280. https://doi.org/10.1093/aob/mcm245.
Gardiner B, Peltola H, Kellomäki S (2000) Comparison of two models for predicting the critical wind speeds required to damage coniferous trees. Ecol Model 129: 1–23. https://doi.org/10.1016/S0304-3800(00)00220-9.
Grabosky J, Bassuk N (2016) Seventeen years’ growth of street trees in structural soil compared with a tree lawn in New York City. Urban For Urban Gree 16: 103–109. https://doi.org/10.1016/j.ufug.2016.02.002.
Heinonen T, Pukkala T, Ikonen VP, Peltola H, Gregow H, Venäläinen A (2011) Consideration of strong winds, their directional distribution and snow loading in wind risk assessment related to landscape level forest planning. Forest Ecol Manag 261: 710–719. https://doi.org/10.1016/j.foreco.2010.11.030.
Hökkä H, Repola J, Laine J (2008) Quantifying the interrelationship between tree stand growth rate and water table level in drained peatland sites within Central Finland. Can J Forest Research 38: 1775–1783. https://doi.org/10.1139/X08-028.
Hökkä H, Salminen H, Ahti E (2012) Effect of temperature and precipitation on the annual diameter growth of Scots pine on drained peatlands and adjacent mineral soil sites in Finland. Dendrochronologia 30: 157–165. https://doi.org/10.1016/j.dendro.2011.02.004.
Hynynen J, Niemistö P, Viherä-Aarnio A, Brunner A, Hein S, Velling P (2010) Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in Northern Europe. Forestry 83: 103–119. https://doi.org/10.1093/forestry/cpp035.
Ikonen VP, Kilpeläinen A, Strandman H, Asikainen A, Venäläinen A, Peltola H (2020) Effects of using certain tree species in forest regeneration on regional wind damage risks in Finnish boreal forests under different CMIP5 projections. Eur J For Res 139: 685–707. https://doi.org/10.1007/s10342-020-01276-6.
Ilisson T, Metslaid M, Vodde F, Jõgiste K, Kurm M (2005) Storm disturbance in forest ecosystems in Estonia. Scand J Forest Res 20: 88–93. https://doi.org/10.1080/14004080510041020.
Jansons Ā, Matisons R, Šēnhofa S, Katrevičs J, Jansons J (2016). High-frequency variation of tree-ring width of some native and alien tree species in Latvia during the period 1965–2009. Dendrochronologia 40: 151–158. https://doi.org/10.1016/j.dendro.2016.10.003.
Jansons J, Līcīte I (2010) Latvia. In: Tomppo E, Gschwantner T, Lawrence M, McRoberts RE (eds) National forest inventories: pathways for common reporting. Springer, Dordrecht, The Netherlands, pp 341–350.
Jutras S, Plamondon AP, Hökkä H, Bègin J (2006) Water table changes following precommercial thinning on post-harvest drained wetlands. Forest Ecol Manag 235: 252–259. https://doi.org/10.1016/j.foreco.2006.08.335.
Kalliokoski T, Nygren P, Sievänen R (2008) Coarse root architecture of three boreal tree species growing in mixed stands. Silva Fenn 42(2): 189–210. https://doi.org/10.14214/sf.252.
Kellomäki S, Peltola H, Nuutinen T, Korhonen KT, Strandman H (2008) Sensitivity of managed boreal forests in Finland to climate change, with implications for adaptive management. Philos T R Soc B 363: 2341–2351. https://doi.org/10.1098/rstb.2007.2204.
Klavina D, Menkis A, Gaitnieks T, Velmala S, Lazdins A, Rajala T, Pennanen T (2016) Analysis of Norway spruce dieback phenomenon in Latvia – a belowground perspective. Scand J Forest Res 31: 156–165. https://doi.org/10.1080/02827581.2015.1069390.
Krisans O, Matisons R, Rust S, Burnevica N, Bruna L, Elferts D, Kalvane L, Jansons A (2020) Presence of root rot reduces stability of Norway spruce (Picea abies): results of static pulling tests in Latvia. Forests 11, article id 416. https://doi.org/10.3390/f11040416.
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 82: 1–26. https://doi.org/10.18637/jss.v082.i13.
Laapas M, Lehtonen I, Venäläinen A, Peltola H (2019) The 10-year return levels of maximum wind speeds under frozen and unfrozen soil forest conditions. Climate 7: 62. https://doi.org/10.3390/cli7050062.
Lenth R (2020) emmeans: estimated marginal means, aka least- squares means. R package version 1.4.4. https://cran.r-project.org/web/packages/emmeans/index.html. Accessed 18 March 2021.
Nicoll BC, Ray D (1996) Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol 16: 891–898. https://doi.org/10.1093/treephys/16.11-12.891.
Nicoll BC, Gardiner BA, Rayner B, Peace AJ (2006) Anchorage of coniferous trees in relation to species, soil type, and rooting depth. Can J Forest Res 1883: 1871–1883. https://doi.org/10.1139/X06-072.
Päätalo ML, Peltola H, Kellomäki S (1999) Modelling the risk of snow damage to forests under short-term snow loading. Forest Ecol Manag 116: 51–70. https://doi.org/10.1016/S0378-1127(98)00446-0.
Peltola H (2006) Mechanical stability of trees under static loads. Am J Bot 93: 1501–1511. https://doi.org/10.3732/ajb.93.10.1501.
Peltola H, Kellomäki S, Väisänen H, Ikonen V-P (1999) A mechanistic model for assessing the risk of wind and snow damage to single trees and stands of Scots pine, Norway spruce, and birch. Can J Forest Res 29: 647–661. https://doi.org/10.1139/x99-029.
Peltola H, Kellomäki S, Hassinen A, Granander M (2000) Mechanical stability of Scots pine, Norway spruce and birch: an analysis of tree-pulling experiments in Finland. Forest Ecol Manag 135: 143–153. https://doi.org/10.1016/S0378-1127(00)00306-6.
Peltola H, Kilpeläinen A, Sauvala K, Räisänen T, Ikonen VP (2007) Effects of early thinning regime and tree status on the radial growth and wood density of scots pine. Silva Fenn 41: 489–505. https://doi.org/10.14214/sf.285.
Peltola H, Ikonen V, Gregow H, Strandman H, Kilpeläinen A, Venäläinen A, Kellomäki S (2010) Impacts of climate change on timber production and regional risks of wind-induced damage to forests in Finland. Forest Ecol Manag 260: 833–845. https://doi.org/10.1016/j.foreco.2010.06.001.
Pukkala T, Laiho O, Lähde E (2016) Continuous cover management reduces wind damage. Forest Ecol Manag 372: 120–127. https://doi.org/10.1016/j.foreco.2016.04.014.
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 18 March 2021.
Reyer CPO, Bathgate S, Blennow K, Borges JG, Bugmann H, Delzon S, Faias SP, Garcia-Gonzalo J, Gardiner BA, Gonzalez-Olabarria JR, Gracia C, Hernández JG, Kellomäki S, Kramer K, Lexer MJ, Lindner M, van der Maaten E, Maroschek M, Muys B, Nicoll B, Palahi M, Palma JHN, Paulo JA, Peltola H, Pukkala T, Rammer W, Ray D, Sabaté S, Schelhaas M-J, Seidl R, Temperli C, Tomé M, Yousefpour R, Zimmermann NE, Hanewinkel M (2017) Are forest disturbances amplifying or canceling out climate change-induced productivity changes in European forests? Environ Res Lett 12, article id 034027. https://doi.org/10.1088/1748-9326/aa5ef1.
Rosenvald K, Ostonen I, Truu M, Truu J, Uri V, Vares A, Lhmus K (2011) Fine-root rhizosphere and morphological adaptations to site conditions in interaction with tree mineral nutrition in young silver birch (Betula pendula Roth.) stands. Eur J For Res 130: 1055–1066. https://doi.org/10.1007/s10342-011-0492-6.
Ruel JC, Larouche C, Achim A (2003) Changes in root morphology after precommercial thinning in balsam fir stands. Can J Forest Res 33: 2452–2459. https://doi.org/10.1139/x03-178.
Rust S (2014) Analysis of regional variation of height growth and slenderness in populations of six urban tree species using a quantile regression approach. Urban For Urban Gree 13: 336–343. https://doi.org/10.1016/j.ufug.2013.12.003.
Samariks V, Krisans O, Donis J, Silamikele I, Katrevics J, Jansons A (2020) Cost–benefit analysis of measures to reduce windstorm impact in pure Norway spruce (Picea abies L. Karst.) stands in Latvia. Forests 11, article id 576. https://doi.org/10.3390/f11050576.
Schelhaas MJ (2008) The wind stability of different silvicultural systems for Douglas-fir in the Netherlands: a model-based approach. Forestry 81: 399–414. https://doi.org/10.1093/forestry/cpn028.
Schmid I, Kazda M (2001) Vertical distribution and radial growth of coarse roots in pure and mixed stands of Fagus sylvatica and Picea abies. Can J Forest Res 31: 539–548. https://doi.org/10.1139/x00-195.
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, Wild J, Ascoli D, Petr M, Honkaniemi J, Lexer MJ, Trotsiuk V, Mairota P, Svoboda M, Fabrika M, Nagel TA, Reyer CPO (2017) Forest disturbances under climate change. Nat Clim Change 7: 395–402. https://doi.org/10.1038/nclimate3303.
Sellin A, Alber M, Keinänen M, Kupper P, Lihavainen J, Lõhmus K, Oksanen E, Sõber A, Sõber J, Tullus A (2017) Growth of northern deciduous trees under increasing atmospheric humidity: possible mechanisms behind the growth retardation. Reg Environ Change 17: 2135–2148. https://doi.org/10.1007/s10113-016-1042-z.
Sikström U, Hökkä H (2016) Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance. Silva Fenn 50, article id 1416. https://doi.org/10.14214/sf.1416.
Silava (2013) Nacionālā meža monitoringa 1.1. aktivitātes “Meža resursu monitorings” metodika. [Methodology of national forest inventory.]. http://www.silava.lv/userfiles/file/Nacionalais%20meza%20monitorings/Me%C5%BEa%20resursu%20monitoringa%20metodika%2026_04_2013.pdf. Accessed 18 March 2021.
Snepsts G, Kitenberga M, Elferts D, Donis J, Jansons A (2020) Stem damage modifies the impact of wind on Norway spruces. Forests 11, article id 463. https://doi.org/10.3390/f11040463.
Suvanto S, Henttonen HM, Nöjd P, Mäkinen H (2016) Forest susceptibility to storm damage is affected by similar factors regardless of storm type: comparison of thunder storms and autumn extra-tropical cyclones in Finland. Forest Ecol Manag 381: 17–28. https://doi.org/10.1016/j.foreco.2016.09.005.
Tenkanen A, Keski-Saari S, Salojärvi J, Oksanen E, Keinänen M, Kontunen-Soppela S (2020) Differences in growth and gas exchange between southern and northern provenances of silver birch (Betula pendula Roth) in northern Europe. Tree Physiol 40: 198–214. https://doi.org/10.1093/treephys/tpz124.
Terho M, Hantula J, Hallaksela AM (2007) Occurrence and decay patterns of common wood-decay fungi in hazardous trees felled in the Helsinki City. Forest Pathol 37: 420–432. https://doi.org/10.1111/j.1439-0329.2007.00518.x.
Valinger E, Fridman J (1997) Modelling probability of snow and wind damage in Scots pine stands using tree characteristics. Forest Ecol Manag 97: 215–222. https://doi.org/10.1016/S0378-1127(97)00062-5.
Valinger E, Fridman J (2011) Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. Forest Ecol Manag 262: 398–403. https://doi.org/10.1016/j.foreco.2011.04.004.
Vasaitis R, Bakys R, Vasiliauskas A (2012) Discoloration and associated fungi in stems of silver birch (Betula pendula Roth.) following logging damage. Forest Pathol 42: 387–392. https://doi.org/10.1111/j.1439-0329.2012.00771.x.
Welch D, Scott D (2008) An estimate of timber degrade in Sitka spruce due to bark stripping by deer in a Scottish plantation. Forestry 81: 489–497. https://doi.org/10.1093/forestry/cpn016.
Wood CJ (1995) Understanding wind forces on trees. In: Coutts MP, Grace J (eds) Wind and trees. Cambridge University Press, Cambridge, pp. 133–164.
Zeng H, Peltola H, Talkkari A, Venäläinen A, Strandman H, Kellomäki S, Wang K (2004) Influence of clear-cutting on the risk of wind damage at forest edges. Forest Ecol Manag 203: 77–88. https://doi.org/10.1016/j.foreco.2004.07.057.
Zeng H, Peltola H, Talkkari A, Strandman H, Venäläinen A, Wang K, Kellomäki S (2006) Simulations of the influence of clear-cutting on the risk of wind damage on a regional scale over a 20-year period. Can J Forest Res 36: 2247–2258. https://doi.org/10.1139/x06-123.
Zubizarreta-Gerendiain A, Pellikka P, Garcia-Gonzalo J, Ikonen VP, Peltola H (2012) Factors affecting wind and snow damage of individual trees in a small management unit in Finland: assessment based on inventoried damage and mechanistic modelling. Silva Fenn 46: 181–196. https://doi.org/10.14214/sf.441.
Total of 67 references.
