Mostafa Farhadi,
Mulualem Tigabu  ,
Per Christer Odén
,
Per Christer Odén
Near Infrared Spectroscopy as non-destructive method for sorting viable, petrified and empty seeds of Larix sibirica
Farhadi M., Tigabu M., Odén P. C. (2015). Near Infrared Spectroscopy as non-destructive method for sorting viable, petrified and empty seeds of Larix sibirica. Silva Fennica vol. 49 no. 5 article id 1340. https://doi.org/10.14214/sf.1340
Highlights
- Near Infrared spectroscopy discriminates filled-viable, empty and petrified seeds of Larix sibirica with 98%, 82% and 87% accuracy, respectively based on spectral differences attributed to moisture and storage reserves
- The classification accuracy reached 100% when sorting seeds into viable and non-viable class
- The results demonstrate that NIR spectroscopy has great potential as non-destructive sorting technique to upgrade seed lot quality.
Abstract
Larix sibirica Ledeb. is one of the promising timber species for planting in the boreal ecosystem; but poor seed lot quality is the major hurdle for production of sufficient quantity of planting stocks. Here, we evaluated the potential of Near Infrared (NIR) Spectroscopy for sorting viable and non-viable seeds, as the conventional sorting technique is inefficient. NIR reflectance spectra were collected from single seeds, and discriminant models were developed with Orthogonal Projections to Latent Structure – Discriminant Analysis (OPLS-DA). The computed model predicted the class membership of filled-viable, empty and petrified seeds in the test set with 98%, 82% and 87% accuracy, respectively. When two-class OPLS-DA model was fitted to discriminate viable and non-viable (empty and petrified seeds combined), the predicted class membership of test set samples was 100% for both classes. The origins of spectral differences between non-viable (petrified and empty) and viable seeds were attributed to differences in seed moisture content and storage reserves. In conclusion, the result provides evidence that NIR spectroscopy is a powerful non-destructive method for sorting non-viable seeds of Larix sibirica; thus efforts should be made to develop on-line sorting system for large-scale seed handling.
Keywords
larch;
NIRS;
OPLS;
precision sowing;
seed sorting;
seed quality
- Farhadi, Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail mostafa.farhadi@slu.se
-
Tigabu,
Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, P.O. Box 49, SE-230 53 Alnarp, Sweden
E-mail
mulualem.tigabu@slu.se
- Odén, Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail per.oden@slu.se
Received 23 March 2015 Accepted 25 August 2015 Published 8 September 2015
Views 106013
Available at https://doi.org/10.14214/sf.1340 | Download PDF

1 Introduction
Interests in growing Larix species (commonly known as larch) in the Northern hemisphere, particularly Fenno-Scandinavia, have grown over the past few decades owing to their better juvenile growth, high timber quality, adaptation to the harsh climate and relatively strong resistance to wind throw and root- and butt rot (Polubojarinov et al. 2000; Pâques 2004; Karlman et al. 2011). Sustainable establishment of larch forests, among other things, demands a continuous supply of high quality seeds for the production of the desired quantity of seedlings in nurseries or for direct sowing out in the field. However, poor seed production is the major afforestation barrier for many Larix species due to insufficient female flowering, lack of pollination and fertilization, degeneration of ovule or early embryo, as well as abnormal development of the female gametophyte and premature abortion of female strobili (Said et al. 1991; Owens 1995; Philipson 1996; Slobodník and Guttenberger 2000). Efforts have been made to increase seed yield using different ways (Philipson 1995; Philippe and Baldet 1997; Bonnet-Masimbert et al. 1998), but a large quantity of empty and petrified seeds up to 50% and 40%, depending on the seed lots, is often encountered in seed bulks during collection (pers. obs). While empty seeds are totally devoid of megagametophyte (storage organ) and embryo, petrified seeds are filled seeds but without embryo – a miniature plant that germinates and produces a normal seedling when sown.
To upgrade the quality of larch seed lots, unproductive seeds, such as empty and petrified seeds, should be removed from the seed bulk during seed cleaning stage. For this purpose, several seed handling methods have been introduced; such as pneumatic and hydraulic separators with 40% separation efficiency (Kaliniewicz et al. 2012), and Incubation, Drying and Separation in water (IDS) with limited success in sorting petrified seeds (Lycksell 1993). The main problem, making use of IDS impossible for large-scale sorting of larch seed lots, is that both viable and petrified seeds lose their moisture at equal rate during the drying process, thus resulting in similar specific density. In addition, using these sorting techniques on large-scale is time consuming, and relatively expensive. Hence, rapid, cost-effective and efficient seed sorting technology is needed to upgrade the quality of larch seed lots, which in turn ensures precision sowing (single seed sowing) in the nursery. Precision sowing is highly desirable in nurseries to maintain lower cost of containerized seedling production.
Here, we demonstrated a rapid and non-destructive technique to distinguish filled-viable, empty and petrified seeds of Larix sibirica Ledeb. (Siberian larch) based on near infrared (NIR) spectroscopy. NIR spectroscopy in combination with multivariate analysis methods can measure moisture and chemical composition of biological materials based on absorption of near infrared radiation by bonds between light atoms, such as C – H, O – H and N – H in biological samples that result in overtones and combination bands detectable in the 780–2500 nm wavelength region (Workman and Weyer 2012). The origin, historical development, theory and principles as well as several applications of NIR spectroscopy can be found in Williams Burns and Ciurczak (2001). Several studies have shown the possibility of classifying seed lots of agricultural crops according to their viability using NIR technique (Agelet and Hurburgh 2014), such as gourd seeds (Min and Kang 2003), spinach seeds (Olesen et al. 2011) and soybean seeds (Wang et al. 2002). Similarly, NIR spectroscopy has been proven useful for separation of viable and non-viable tree seeds (Lestander and Odén 2002; Soltani et al. 2003; Tigabu and Odén 2004; Tigabu et al. 2004; 2007). To our knowledge, there is no study that attempted to apply NIR technology for simultaneously sorting filled-viable, empty, petrified seeds of Siberian larch, except the study by Tigabu and Odén (2004) that reported discrimination of filled-viable, empty and insect-attacked seeds of three Larix species with 90% – 100% accuracy using NIR transmittance spectroscopy. The present study is unique in that NIR spectroscopic technique was applied for the first time to discriminate petrified and empty seeds from filled viable seeds.
2 Materials and methods
2.1 Seed samples
For this study, four seed lots of L. sibirica were obtained from the Forest Research Institute, Sävar, Sweden, and sorted into filled, empty and petrified seeds by digital X-ray analysis (MX-20 Cabinet X-ray System; Faxitron X-ray LLC, Lincolnshire, IL 600069). Seeds with visible embryonic cavity and megagametophyte (storage organ) were considered as viable; seeds without any content (megagametophyte and embryo) were considered as empty; and seeds without embryonic axis and purely white hardened content were considered as petrified. Accordingly, a total of 675 seed samples from four different seed lots was sorted into 225 filled-viable, empty and petrified seeds each and employed for NIR analysis.
2.2 Acquisition of NIR spectra
NIR reflectance spectra of single seeds, expressed in the form of log (1/Reflectance), were collected with XDS Rapid Content Analyzer (FOSS NIRSystems, Inc.) from 400 to 2498 nm at an interval of 0.5 nm. Individual seeds were directly placed at the center of the scanning glass window of the instrument with 9 mm aperture at stationary module and then covered with the instrument’s lid that had black background. Prior to collecting the NIR spectra from every batch of seed lot, reference reflectance measurement was taken on standard built-in reference of the instrument. In addition, reference measurements were taken after every 20 scans to reduce the effects of possible instrumental drift. For every seed, 32 scans were made and the average value recorded. Finally, the scanned seeds were arranged individually for germination tests to further verify the viability status. All empty and petrified seeds did not germinate while all filled-viable seeds germinated.
2.3 Multivariate discriminant analysis
The spectral data were divided into calibration and test sets. The calibration set was composed of 540 spectra (180 spectra for filled-viable, empty and petrified seeds each) drawn from two seed lots. The test set samples were drawn from the other two seed lots composed of 135 spectra (45 spectra per seed lot fraction). The visible region was excluded during model development because it appeared to carry very little useful information for discriminating the seed lot fractions. Initially, Orthogonal Projections to Latent Structures-Discriminant Analysis (OPLS-DA) modeling was performed to discriminate filled-viable, empty and petrified seeds simultaneously using the digitized NIR spectra as regressor and a y-matrix of dummy variables (1 for member of a given class, 0.0 otherwise) as regressand. The OPLS-DA modelling approach integrates both data pre-processing to remove unwanted systematic noise in the spectra and subsequent modelling. NIR spectroscopic data are not usually amenable for direct analysis due to light scattering, base line shift, instrumental drift, and path length differences (Tigabu and Odén 2004; Tigabu et al. 2004). This unsystematic noise in the spectra should be remove to enhance signal to noise ratio. The OPLS-DA modelling approach removes the spectral noise by using information in the categorical response matrix Y (a matrix of dummy variables in our case) to decompose the X matrix (the NIR spectral data) into three distinct parts: (1) the predictive score matrix and its loading matrix for X, (2) the corresponding Y-orthogonal score matrix and loading matrix of Y-orthogonal components, and (3) the residual matrix of X (Trygg and Wold 2002). The final discriminant model was then computed using the predictive spectral variations. As there was some overlap between empty and petrified seeds, two-class class OPLS-DA model was also developed by merging empty and petrified seeds to form a non-viable class and filled-viable seeds as a viable class.
All calibrations were developed on mean-centered data sets and the numbers of significant model components were determined by a seven-segment cross validation. A component was considered significant if the ratio of the prediction error sum of squares (PRESS) to the residual sum of squares of the previous dimension (SS) was statistically smaller than 1.0 (Eriksson et al. 2006). Finally, the predictive ability of the fitted models was evaluated using test sets, and seeds were considered as members of a given class if predicted values were greater than an acceptance threshold (≥ 0.5) and all others were considered as non-members.
2.4 Absorption bands accounted for class discrimination
Spectral regions accounted for the discrimination of viable and non-viable seeds were determined by a parameter called Variable Influence on Projection (VIP), which was computed as follows.
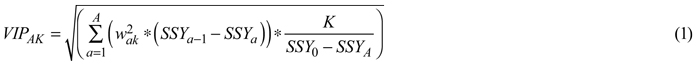
VIP is a weighted sum of squares of the OPLS weights (w), taking into account the amount of explained Y-variance (SSY) of a dimension of K variables. Its major advantage is that there will be only one VIP-vector, summarizing all components and Y-variables; thereby enabling us to identify absorption bands influencing the discriminant models. As a rule, predictors with VIP value more than 1 have a strong influence on the model; but a cut-off around 0.7–0.8 has been suggested to discriminate between relevant and irrelevant predictors (Eriksson et al. 2006). All calculations were performed using Simca-P + software (Version 13.0.0.0, Umetrics AB, Sweden).
3 Results
3.1 Overview of spectral profile
The average absorbance values for filled-viable, empty and petrified seeds of L. sibirica showed a similar spectral profile with major peaks at 1450 nm and 1920 nm with small peaks around 1200 nm, 1670 nm, 2100 nm and 2270 nm (Fig. 1A). The absorbance values for petrified seeds were higher than empty and filled-viable seeds across the whole NIR spectral region while filled-viable seeds had slightly higher absorbance value than empty seeds. The difference spectrum shows that the non-viable seeds (empty and petrified seeds combined) had higher absorbance values in the 780–1100 nm with broad peaks centered at 970 nm (Fig. 1B). As a whole, the NIR spectra contained adequate information to discriminate filled-viable, empty and petrified seeds of L. sibirica.
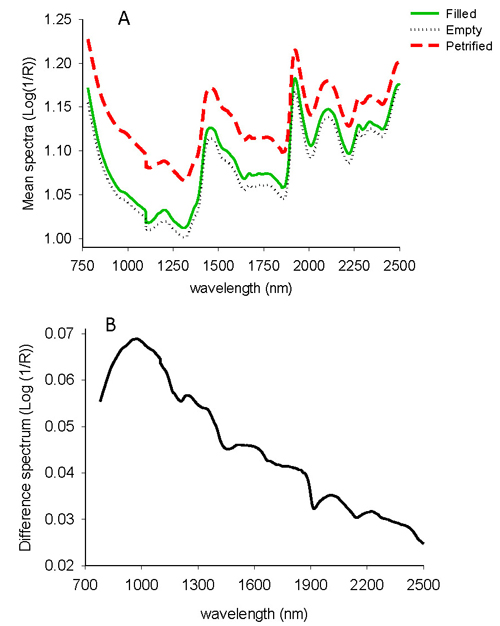
Fig. 1. Average uncorrected NIR spectra of filled-viable, empty and petrified seeds of L. sibirica (A), and difference spectrum of non-viable and viable seeds (B).
3.2 Discrimination of filled-viable, empty and petrified seeds
The OPLS-DA model developed to simultaneously discriminate the three seed lot fractions had two predictive and 13 Y-orthogonal components (A = 2 + 13). The total spectral variation described by the model was 100%; of which the predictive variation (R2XP) accounted for 26.7% and the Y-orthogonal spectral variation (R2Xo) constituted 73.3%. The predictive spectral variation, in turn, modelled 84.2% of the class variation (R2Y) in the calibration set with 82.0% prediction accuracy (Q2cv) according to cross validation (Table 1). The score plot for the predictive components (Fig. 2A) showed clear separation of petrified seeds from filled-viable and empty seeds along the first component (tp[1]) and filled-viable seeds from the other two seed lot fractions along the second component (tp[2]). The corresponding predictive loading plot revealed that the absorption band in 780–1100 nm with a broad peak centered at 970 nm was attributed to separating petrified seeds from filled-viable and empty seeds (Fig. 2B). Whereas absorption bands in 1140–1256 nm, 1268–1418 nm, 1590–2035 nm with major peaks at 1196 nm, 1390 nm, 1706 nm, 1859 nm, 1878 nm and 1986 nm were attributed to discriminating filled-viable seeds from petrified and empty seeds (Fig. 2C). The score plot for the first two orthogonal components (to[1] versus to[2]) showed that few samples from each class were dispersed slightly away from the swarm of the majority of the samples, and the corresponding loading plot revealed that absorption band in 780–1100 nm with a peak at 970 nm appeared to contribute to this unwanted spectral variation (data not shown).
| Table 1. A statistical summary of discriminant models computed to classify viable, empty and petrified seeds of L. sibirica into three and two classes. | |||||
| Class | A | R2Xp | R2X0 | R2Y | Q2 cv |
| Three | 2 + 13 | 0.267 | 0.733 | 0.842 | 0.820 |
| Two | 1 + 12 | 0.041 | 0.959 | 0.937 | 0.931 |
| A = number of significant components to build the model (the first and second values represent the number of components for predictive and orthogonal variations, respectively); R2Xp = the predictive spectral variation; R2Xo = Y-orthogonal variation (spectral variation uncorrelated to class discrimination) for the OPLS-DA models; R2Y = the modelled class variation; and Q2cv = the predictive power of a model according to cross validation. | |||||
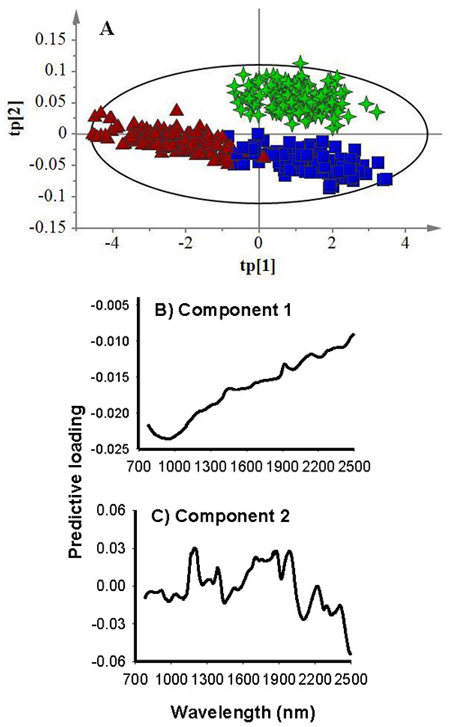
Fig. 2. Score plot for the first (tp[1]) and second (tp[2]) predictive components (A) showing clear clustering patterns of filled-viable (green star), empty (blue box) and petrified (brown triangle) seeds, and loading plots for the first (B) and second (C) predictive components showing absorption bands accounted for class discrimination.
For test set samples, the computed three-class OPLS-DA model correctly assigned filled-viable, empty and petrified seeds with 98%, 82% and 87% classification accuracy (Table 2). None of the filled-viable seeds were misclassified as member of other class, but one sample appeared to have no class. Similarly, neither empty nor petrified seeds were misclassified as filled-viable seeds, but nearly 11% of empty seeds in the test set was misclassified as petrified seed and 4% as both empty and petrified seeds while 4% of petrified seeds were misclassified as empty and as both empty and petrified. Nearly 9% of petrified seeds and 2% of empty seeds had no class.
| Table 2. Predicted class membership of viable, empty and petrified seeds in the test sets by three-class OPLS-DA modelling of single seed NIR spectra. | ||||||||||
| Seed lot fractions | No. of test | 1 | 2 | 3 | No class | 1 & 2 | 1 & 3 | 2 & 3 | 1 & 2 & 3 | Classification accuracy (%) |
| Viable (1) | 45 | 44 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 98 |
| Empty (2) | 45 | 0 | 37 | 5 | 1 | 0 | 0 | 2 | 0 | 82 |
| Petrified (3) | 45 | 0 | 1 | 39 | 4 | 0 | 0 | 1 | 0 | 87 |
3.3 Discrimination of viable and non-viable seeds
Two-class OPLS-DA model, fitted to discriminate seed lot fractions into viable and non-viable classes, had one predictive and 12 Y-orthogonal components, and the predictive variation of the spectra (R2XP) was to 4.1%, but this small fraction of the predictive variation still explained 93.7% of the between-class variation with 93.1% predictive ability of the model according to cross validation (Table 1). The largest proportion of the spectral variation was uncorrelated to between-class variation (R2Xo = 0.959; Table 1). The score plot showed a symmetrical separation of viable and non-viable seeds along the predictive component and within-class variation along the Y-orthogonal component (Fig. 3A). Although some seeds from each viability class fell outside of the 95% confidence ellipse according to Hotelling’s T2 test (a multivariate generalization of Student’s t-test), they were not strong outliers. The corresponding predictive loading for the first component revealed that absorption peaks centered around 970 nm, 1250 nm and 1352 nm were mainly accounted for discriminating non-viable seeds from viable seeds (Fig. 3B); while the Y-orthogonal loading plot showed a broad absorption band in 1300–1900 nm that were uncorrelated to between-class variation (Fig. 3C). For test set samples, the computed two-class model assigned viable and non-viable classes with 100% accuracy (Fig. 4A).
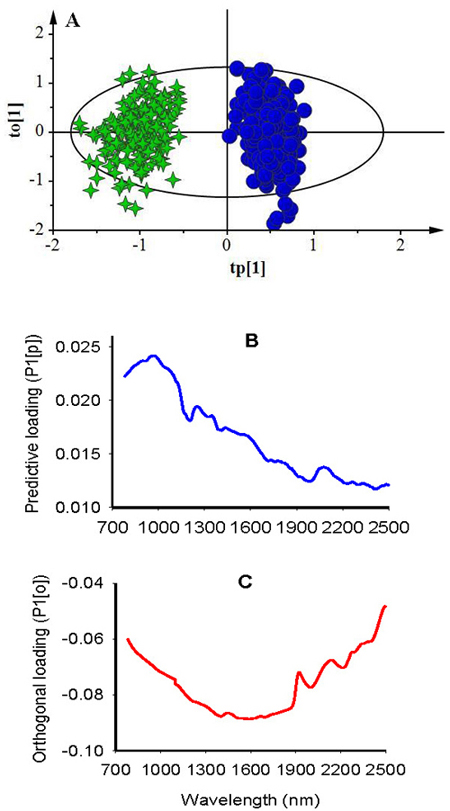
Fig. 3. Score plot for the first predictive (tp[1]) versus orthogonal (to[1]) components showing symmetrical separation of viable (green stars) and non-viable (blue dots) seeds (A); loading plot for the first predictive component (P1[p]) showing absorption bands correlating to seed classes (B); and loading plot for the first orthogonal component (P1[o]) showing absorption bands that are not correlated to class discrimination (C).
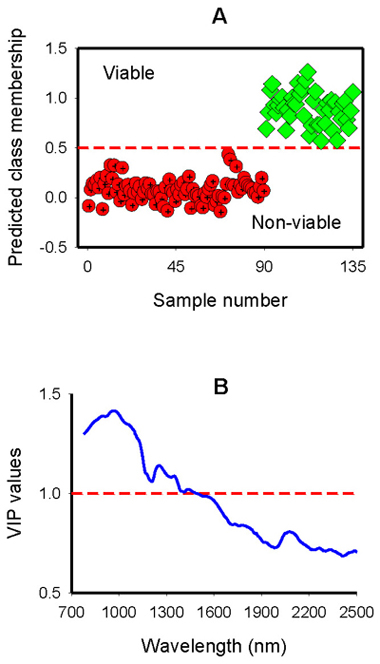
Fig. 4. Predicted class membership of non-viable (empty and petrified) and viable seeds in the test set by two-class OPLS-DA modelling of NIR spectral region (A); and Plot of Variable Influence on Projection, VIP, (B) showing spectral regions that influenced the discrimination of viable from non-viable seeds. The horizontal dotted line in panel A is the class limit (Ypredicted > 0.5) for assigning the test sets into viable class otherwise non-viable, and that in panel B is the cut-off limit (1.0) for discriminating relevant and irrelevant predictors.
3.4 Absorption bands accounted for discrimination of viable and non-viable seeds
The VIP plot shows that absorbance in 780–1300 nm with a major peak centered around 970 nm, and a smaller peak at 1256 nm as well as a small bump at 1350 nm had a strong influence on the discrimination of filled-viable and non-viable seeds (VIP > 1; Fig. 4B). The spectral region between 1414 nm and 1644 nm with a broad absorption band also accounted for class discrimination. Other regions of interest in the longer wavelength range appeared at 2080 nm that contributed well for class discrimination (VIP = 0.81).
4 Discussion
The results demonstrate the potential of NIR spectroscopy as a non-destructive method for successfully sorting filled-viable, empty and petrified seeds of L. siberica. By extracting irrelevant spectral variations that are not useful for class discrimination, the OPLS-DA modelling results in parsimonious models (Table 1). Dimensional complexity is an important factor in the interpretation of multivariate analysis and parsimonious models with few dimensions (components) are often highly preferred (Trygg and Wold 2002; Pinto et al. 2012). In the present study, the proportion of spectral variations that is uncorrelated with class discrimination is larger than the predictive variation (Table 1). This might be attributed to spectral redundancy, as the absorbance values were measured at 0.5 nm wavelength interval; hence it is legitimate to expect a high degree of redundancy in the absorbance values at this scale of resolution. In addition, variations in size and moisture content among individual seeds induce path length difference and light scattering, which in turn are uncorrelated to class discrimination (Tigabu and Odén 2004). This is further evidenced from the Y-orthogonal score plot (data not shown) where few samples from each class positioned far away from the bulk of the samples while the corresponding orthogonal loading plot showed a major absorption peak at 970 nm, which is attributed to water (Lestander and Odén 2002). Thus, variation in moisture content among individual seeds could be a source of unwanted spectral variation that had no correlation with class discrimination. Nevertheless, NIR spectroscopy is highly sensitive in detecting subtle differences as low as 0.1% of the total concentration of the analyte (Osborne et al. 1993) while multivariate analysis is powerful in extracting such information from the spectra (Eriksson et al. 2006).
The absorption band in 780–1100 nm with a broad peak centered at 970 nm was positively correlated with petrified seeds (Fig. 2A). This region is characterized by O – H stretching second overtone where absorption spectra of aliphatic and aromatic hydroxyl groups as well as starch and water overlap (Osborne et al. 1993; Workman and Weyer 2012). Lestander and Odén (2002) found the absorption peak at 970 nm useful to detect moisture difference between filled-viable and dead-filled seeds of Scots pine. As petrified seeds dry more slowly and maintain fairly high moisture content longer than empty seeds during drying, the origin of spectral differences between petrified and empty seeds could be attributed to divergence in moisture content between these seed lot fractions.
For discriminating filled-viable seeds from empty and petrified seeds, the model utilized spectral information in the longer wavelength regions with major peaks at 1200 nm, 1390 nm, 1706 nm, 1859 nm, 1878 nm and 1986 nm (Fig. 2B). The 1100–1300 nm region is characteristic of the second overtone of C – H stretching vibration and functional group responsible for absorption are methyl and methylene (Shenk et al. 2001; Workman and Weyer, 2012). It has been shown that the major absorption band in fat or oil is due to a long chain fatty acid moiety that gives rise to CH2 second overtone at 1200 nm; and the band near 1180 nm has been assigned as the second overtone of the fundamental C – H absorption of pure fatty acids containing cis double bonds, e.g. oleic acid (Sato et al. 1991; Osborne et al. 1993). The 1300–1600 nm regions presents two peaks at 1320 nm and 1390 nm, which corresponds to C – H combination and first overtone of N – H stretching vibration due to absorption by CH2 and protein moieties (Shenk et al. 2001). Protein moieties could be the possible source of variation for discriminating filled-viable seeds from empty and petrified seeds in this region, as the absorption band in this region has been shown to play minor role for oil and fat classification (Hourant et al. 2000). The 1600–1900 nm shows several bumps and peaks in the vicinity of 1706 nm, 1760 nm, 1856 nm and 1876 nm. The region is characteristic of the first overtone of the C – H stretching vibration of methyl and methylene groups (Shenk et al. 2001).) The absorption peaks at 1710 nm and 1725 nm correlates to linoleic and oleic acids respectively as well as triolein in the vicinity of 1725 nm, trilinolein near 1717 nm, and trilinolenin near 1712 nm (Sato et al. 1991). The absorption bands observed in our study could, therefore, be correlated to the dominant fatty acids in L. siberica seeds: linoleic, Δ5-olefinic, pinolenic and oleic acids, which account 42.66%, 30.8%, 30.57% and 16.67% of the total seed fatty acids, respectively (Wolff et al. 1997).
The 1850–2050 nm region shows one absorption band, centered near 1986 nm that arises from C = O stretch second overtone, combination of O – H stretch and HOH deformation, as well as O – H bend second overtone. Several compounds, notably protein, starch and water, show characteristic absorption in this region (Shenk et al. 2001). We believe that the absorption band in this region correlates more to water than to other compounds because viable seeds often retain more bound water than empty seeds. As a whole, the discriminant models utilized spectral differences attributed to moisture and storage reserves as a basis to discriminate filled-viable, empty and petrified seeds.
Our attempt to discriminate larch seed lots into viable and non-viable class was very successful; i.e. resulting in complete separation of filled-viable seeds from both empty and petrified seeds. This is more appealing from practical seed handling point of view, as it ensures greater handling efficiency. The VIP plot (Fig. 4B), which is analogous to the difference spectrum (Fig. 1B), shows that non-viable seeds had higher absorbance values in 780–1200 nm and 1210–1389 nm regions than viable seeds. The absorption bands in these regions are characterized by O – H stretching second overtone and second overtone of C – H stretch, respectively due to absorption by aliphatic and aromatic hydroxyl groups as well as starch and water overlap (Osborne et al. 1993; Workman and Weyer, 2012). Tigabu and Odén also found higher absorption in 780–1100 nm by empty seeds of three Larix species. Thus, the absorbance values from both petrified and empty seeds in these regions dominate the spectral signature than viable seeds, and hence accounted for discrimination of non-viable seed class from that of viable seed class.
5 Conclusion
The findings demonstrate the prospect of NIR spectroscopy as a non-destructive method for upgrading the quality of larch seeds. The technique is so accurate and time-saving and can be applied for sorting larch seeds according to their viability without any pretreatment. Moreover, the OPLS-DA modelling approach not only results in parsimonious models but also provides additional information to examine within-class variation and its cause to better understand the modelling process. Therefore, efforts should be made to scale-up the technique to on-line sorting system for large-scale seed handling operations.
Acknowledgement
Financial support for the first author from Karl Erik Önnesjös Foundation is gratefully acknowledged. We acknowledge the Forest Research Institute of Sweden in Sävar for providing us L. sibirica seeds, and particularly Mr. Erik Walfridsson is acknowledged for valuable discussions on the seed-related problem of this species, and for sorting seed lots according to their viability class by X-ray analysis.
References
Agelet L.E., Hurburgh C.R. (2014). Limitations and current applications of Near Infrared Spectroscopy for single seed analysis. Talanta 121: 288–299. http://dx.doi.org/10.1016/j.talanta.2013.12.038.
Bonnet-Masimbert M., Baldet P., Pâques L.E., Philippe G. (1998). From flowering to artificial pollination in larch for breeding and seed orchard production. Forestry Chronicle 74: 195–202.
Burns D.A., Ciurczak E.W. (2001). Handbook of Near-Infrared Analysis. 2nd edition. Marcel Dekker Inc., New York. 803 p.
Eriksson L., Johansson E., Kettaneh-Wold N., Trygg J., Wikström C., Wold S. (2006). Multi- and megavariate data analysis. Basic principles and applications. Second revised and enlarged edition. Umetrics Academy, Umeå, Sweden. 425 p.
Hourant P., Baeten V., Morales M.T., Meurens M., Aparicio R. (2000). Oil and fat classification by selected bands of Near-Infrared Spectroscopy. Applied spectroscopy 54: 1168–1174. http://dx.doi.org/10.1366/0003702001950733.
Kaliniewicz Z., Markowski P., Anders A., Rawa T., Liszewski A., Fura S. (2012). Correlations between the germination capacity and selected attributes of European larch seeds (Larix Decidua Mill.). Technical Sciences 15: 229–242.
Karlman L., Fries A., Martinsson O., Westin J. (2011). Juvenile growth of provenances and open pollinated families of four Russian larch species (Larix Mill.) in Swedish field tests. Silvae Genetica 60: 165–177.
Lestander T.A., Odén P.C. (2002). Separation of viable and non-viable filled Scots pine seeds by differentiating between drying rates using single seed near infrared transmittance spectroscopy. Seed Science and Technology 30: 383–392.
Lycksell S. (1993). Seed conditioning of Larix sukaczewii (Ledeb.) using PREVAC, IDS and a flotation technique. Master thesis. Swedish University of Agricultural Sciences, Umeå, Sweden. 23 p.
Min T.G., Kang W.S. (2003). Nondestructive separation of viable and nonviable gourd (Lagenaria siceraria) seeds using single seed near infrared spectroscopy. Journal of Korean Horticultural Science 44: 545–548.
Olesen M.H., Shetty N., Gislum R., Boelt B. (2011). Classification of viable and non-viable spinach (Spinacia oleracea L.) seeds by single seed NIR spectroscopy and extended canonical variates analysis. Journal of Near Infrared Spectroscopy 19: 171–180. http://dx.doi.org/10.1255/jnirs.928.
Osborne B.G., Fearn T., Hindle P.H. (1993). Practical NIR spectroscopy: with applications in food and beverage analysis. 2nd ed. Longman Scientific and Technical, Harlow, UK. 227 p.
Owens J.N. (1995). Reproductive biology of larch. In: Schmidt W.C., McDonald K.J. (eds.). Ecology and management of Larix forests: a look ahead. USDA, Forest Service, Intermountain Research Station. General Technical Report-INT-319. p. 97–109.
Owens J.N., Morris S.J., Catalano G.L. (1994). How the pollination mechanism and prezygotic and postzygotic events affect seed production in Larix occidentalis. Canadian Journal of Forest Research 24: 917–927. http://dx.doi.org/10.1139/x94-121.
Philippe G., Baldet P. (1997). Electrostatic dusting: an efficient technique of pollination in larch. Annals of Forest Science 54: 301–310. http://dx.doi.org/10.1051/forest:19970307.
Philipson J.J. (1995). Effects of cultural treatments and gibberellin A4/7 on flowering of container-grown European and Japanese larch. Canadian Journal of Forest Research 25: 184–192. http://dx.doi.org/10.1139/x95-022.
Philipson J.J. (1996). Effects of girdling and gibberellin A4/7 on flowering of European and Japanese larch grafts in an outdoor clone bank. Canadian Journal of Forest Research 26: 355–359. http://dx.doi.org/ 10.1139/x26-039.
Pinto R.C., Trygg J., Gottfries J. (2012). Advantages of orthogonal inspection in chemometrics. Journal of Chemometrics 26: 231–235. http://dx.doi.org/10.1002/cem.2441.
Polubojarinov O.I., Chubinsky A.N., Martinsson O. (2000). Decay resistance of Siberian larch wood. Ambio 29: 352–353. http://dx.doi.org/10.1579/0044-7447-29.6.352.
Said C., Villar M., Zandonella P. (1991). Ovule receptivity and pollen viability in Japanese larch (Larix leptolepis Gord.). Silvae Genetica 40: 1–6.
Sato T., Kawano S., Iwamoto M. (1991). Near-Infrared Spectral patterns of fatty acid analysis from fats and oils. Journal of American Oil Chemists Society 68: 827–833. http://dx.doi.org/10.1007/BF02660596.
Shenk J.S., Workman J.J., Westerhaus M.O. (2001). Application of NIR spectroscopy to agricultural products. In: Burns D.A., Ciurczak E.W. (eds.). Handbook of near-infrared spectroscopy. Marcel Dekker Inc., New York. p. 419–474.
Slobodník B., Gutternberger H. (2000). Ovule, megaspores and female gametophyte formation in Larix decidua Mill. (Pinaceae). Acta Biologica Cracoviensia 42: 93–100.
Soltani A., Lestander T.A., Tigabu M., Odén P.C. (2003). Prediction of viability of oriental beechnuts, Fagus orientalis using near infrared spectroscopy and partial least squares regression. Journal of Near Infrared Spectroscopy 11: 357–364. http://dx.doi.org/10.1255/jnirs.387.
Tigabu M., Odén P.C. (2004). Simultaneous detection of filled, empty and insect-infested seeds of three Larix species with single seed near infrared transmittance spectroscopy. New Forests 27:39–53. http://dx.doi.org/10.1023/A:1025037408922.
Tigabu M., Odén P.C., Shen T.Y. (2004). Application of near infrared spectroscopy for the detection of internal insect infestation in Picea abies seed lots. Canadian Journal of Forest Research 34: 76–84. http://dx.doi.org/10.1139/x03-189.
Tigabu M., Fjellström J., Odén P.C., Teketay D. (2007). Germination of Juniperus procera seeds in response to stratification and smoke treatments, and detection of insect-damaged seeds with VIS + NIR spectroscopy. New Forests 33: 155–169. http://dx.doi.org/10.1007/s11056-006-9020-9.
Trygg J., Wold S. (2002). Orthogonal projections to latent structures (O-PLS). Journal of Chemometrics 16: 119–128. http://dx.doi.org/10.1002/cem.695.
Wang D., Ram M.S., Dowell F.E. (2002). Classification of damaged soybean seeds using near-infrared spectroscopy. Transactions of the American Society of Agriculture Engineers 45: 1943–1948. http://dx.doi.org/10.13031/2013.11410.
Wollf R.L., Comps B., Marpeau A.M. (1997). Taxonomy of Pinus species based on the seed oil fatty acid compositions. Trees 12: 113–118. http://dx.doi.org/10.1007/PL00009698.
Workman J., Weyer L. (2012). Practical Guide and Spectral Atlas for Interpretive Near-Infrared Spectroscopy. 2nd edition. CRC Press, Taylor & Francis, Boca Raton, Florida. 326 p. http://dx.doi.org/10.1201/b11894.
Total of 31 references.
