Aleksey Fedorkov  ,
Ludmila Gutiy
,
Ludmila Gutiy
Performance of lodgepole pine and Scots pine in field trials located in north-west Russia
Fedorkov A., Gutiy L. (2017). Performance of lodgepole pine and Scots pine in field trials located in north-west Russia. Silva Fennica vol. 51 no. 1 article id 1692. https://doi.org/10.14214/sf.1692
Highlights
- Stem volume was bigger for lodgepole pine in comparison to local Scots pine except for the southernmost origin
- The proportion of stems with no defects was lower for all lodgepole pine seed sources than for local Scots pine
- Lodgepole pine stem growth traits were significantly related to latitude of seed origin.
Abstract
Mortality, stem growth and quality of lodgepole pine (Pinus contorta var. latifolia) originating from the six Swedish seed orchards and local Scots pine (Pinus sylvestris L.) were estimated in four field trials established in the Komi Republic (north-west Russia). A randomized row-plot design with 6–12 replicates of each entry was used. The tree mortality was slightly higher for Scots pine than that for lodgepole pine, except for the lodgepole pine seed sources of the southern origins with lower survival. Scots pine stem quality was better than that of lodgepole pine, but the lodgepole pine stem growth was faster except the seed source of the southernmost origin. The lodgepole pine seed sources of northern origins had better stem growth (height, diameter at breast height and volume), while the effect of latitude on the quality traits was insignificant.
Keywords
Pinus sylvestris;
stem defects;
mortality;
Pinus contorta;
stem growth
-
Fedorkov,
Institute of Biology, Komi Science Center, Russian Academy of Sciences, 28 Kommunisticheskaya st., Syktyvkar 167982, Russia
 http://orcid.org/0000-0001-7800-7534
E-mail
fedorkov@ib.komisc.ru
http://orcid.org/0000-0001-7800-7534
E-mail
fedorkov@ib.komisc.ru
- Gutiy, Syktyvkar Forest Institute (branch), Saint-Petersburg State Forest Technical University, 39 Lenin st., Syktyvkar 167000, Russia E-mail lguti@mail.ru
Received 31 August 2016 Accepted 26 January 2017 Published 1 February 2017
Views 67920
Available at https://doi.org/10.14214/sf.1692 | Download PDF

1 Introduction
The North American tree species lodgepole pine (Pinus contorta var. latifolia) was introduced in many European countries (Lindgren 1993). Lodgepole pine is one of the most familiar, widespread and important pine species in Pacific North America. In British Columbia, it is the most widely distributed, harvested and planted species and one of the most commercially valuable (Owens 2006).
In Nordic countries, lodgepole pine might be a suitable species for producing pulp and construction wood, wooden biomass, fuels and chemicals (Elfving et al. 2001; Backlund and Bergsten 2012). Numerous studies revealed that lodgepole pine in northern Europe is superior to Scots pine in both survival and stem growth (Elfving and Norgren 1993; Ruotsalainen and Velling 1993; Raevsky 2010). However, in Sweden lodgepole pine is inferior to Scots pine in stem quality (Hansson and Karlman 1997; Nilsson and Cory 2010). In northern Russia, accordingly to Raevsky and Pekkoev (2013) the stem straightness was slightly better and occurrence of forks was lower for Scots pine than for lodgepole pine at the age of 25 years in field trial located in southern Karelia. Similar results were also obtained by Feklistov et al. (2008) in four plantations aged 7–21 years located in Archangelsk region where frequency of stem defects was higher for lodgepole pine than that for Scots pine.
The large-scale introduction of lodgepole pine was initiated in the 1970s into Swedish forests and plantations of this species covers about 475 000 ha of pure stands and 200 000 ha of mixed stands (Nilsson and Cory 2010). The base breeding material was collected in western Canada and major series of progeny trials as well as six seedling seed orchards were established (Ericsson 1994).
The climate in the Komi Republic is continental and differs from the more maritime climate in Fenno-Scandinavia, but however it is more similar to that in the area of lodgepole pine distribution in inferior British Columbia. The aim of this study was to examine mortality, stem growth and quality of the Swedish lodgepole pine seed orchards’ crops in this new environment of the Komi Republic (north-west Russia) and to compare them with the performance of local Scots pine. These results would be important for a proper selection of seed sources for commercial planting.
2 Materials and methods
2.1 Field experiments
The study was performed in four field trials located in sites with poor sand soil in the Komi Republic (north-west Russia) established in spring 2004 and 2006. Thinning was not carried out in all trials. All trials included identical lodgepole pine material from the six Swedish seed orchards (seed sources) (Fig. 1; Table 1 and 2). Four Scots pine bulk seed lots (provenances) collected in unimproved mature stands after final felling (25–30 felled trees) located at a distance of 15–20 km from each trial site were used as a standard. Seeds of all entries were sown in containers (7×7 cells/container) with a cell size of 128 cm3 in May 2003 and 2004. Seedlings were grown in a greenhouse without heating and additional light; the plastic cover was removed ten weeks after sowing. For three trials (Ukhta, Storojevsk and Koygorodok), 1-year-old potted seedlings were planted. For Syktyvkar field test, 2-year-old potted seedlings were planted. A randomized row-plot design with 6–12 replicates of each entry was employed for all trials.
Fig. 1. Location of the lodgepole pine seed orchards and field trials.
| Table 1. Description of the field trials included in the study. | ||||||||
| Trial | Year of planting | Latitude, Longitude | Altitude (m a.s.l.) | Temperature suma | Spacing (m) | Total no. seedlings | No. blocks | Site type |
| Ukhta | 2004 | 63°40´N, 53°37´E | 140 | 863 | 2.5×2.0 | 3507 | 12 | Sand quarry |
| Storojevsk | 2004 | 61°53´N, 52°45´E | 104 | 1037 | 2.0×2.0 | 3865 | 11 | Farm sandy land |
| Koygorodok | 2004 | 60°22´N, 51°17´E | 167 | 1155 | 2.0×1.5 | 2363 | 10 | Sand quarry |
| Syktyvkar | 2006 | 61°40´N, 51°03´E | 132 | 1075 | 2.0×1.0 | 2374 | 6 | Forest sandy land |
| a Expected mean temperature sum in degree-days, threshold temperature +5 °C (Galenko 1999) | ||||||||
| Table 2. Identification of the lodgepole pine seed sourcesa (seedlings seed orchards) studied (after Ericsson 1994). | |||||||
| No. | Name | Site latitude | Year of establishment | Area, ha | Number of mother trees | Range of source tree origins in Canada | Range of proposed use in Sweden |
| 711 | Närlinge | 60°03´N | 1987 | 18.3 | 100 | 60°44’–63°40´N | 67° and north |
| 712 | Oppala | 60°46´N | 1983,–85 | 12.7 | 211 | 59°17’–62°40´N | 66°30’–67°00´N |
| 713 | Skörserum | 58°00´N | 1984,–85 | 18.2 | 210 | 57°36’–60°38´N | 66°00–66°30´N |
| 714 | Larslund | 58°46´N | 1982 | 15.1 | 178 | 55°38’–58°40´N | 64°30’–66°00´N |
| 715 | Rumhult | 57°41´N | 1981 | 21.7 | 218 | 54°17’–56°08´N | 62°30’–64°30´N |
| 716 | Österby | 58°08´N | 1981 | 14.3 | 300 | 50°51’–53°50´N | 60°30’–62°40´N |
| a In this study seed sources Närlinge, Oppala and Skörserum are considered as northern origins and Larslund, Rumhult and Österby as southern ones | |||||||
2.2 Trait measurements and assessments
All trees in each field experiment were assessed after 12 (Ukhta, Storojevsk and Koygorodok) and 10 (Syktyvkar) growing seasons in the field in autumn 2015. The trees were categorized in three classes: tree with no stem defects, tree with stem defects (crookedness, forking, ramicorn branches and spike knots) and dead tree. Stem crookedness was visually graded in two classes: class 1 = stems with a weakly crooked bole and class 2 = stems with a severely crooked bole. Forked trees are those with two stems having about equal stem diameters. Ramicorn branches are large, steep-angled branches that occur when terminal shoot temporarily loses apical dominance to a lateral branch (Magalska and Howe 2014). Spike knot is a leader shoot change. Type of defects was recorded. Small living trees (height < 1.5 m) were not evaluated, but were also recorded to be used for calculation of proportions.
Trees without defects were callipered at breast height which was defined as 1.3 m (DBH, 1 mm accuracy) and tree height was recorded in meter (0.1 m accuracy) using a measure stick. Since there is no volume function for lodgepole pine for Russian conditions we used Scots pine volume functions for the Komi Republic condition for both species. The stem volume (on bark) was calculated using Tjurin’s (1972) equations. For trees with height < 5.0 m:
![]()
For trees with height > 5.0 m:

where V is the volume (m3), h is the height (m) and d is DBH (cm). The volumes were then converted to units of dm3.
2.3 Statistical analysis
Statistical analysis of growth traits (height, DBH and stem volume) was performed on a plot mean basis as independent units. These characteristics exhibited normal distribution accordingly to the Kolmogorov-Smirnov test. For quality traits (proportion of trees with or no defects and mortality) the frequencies of tree categories for each plot were also calculated. Tests of normal distribution of quality parameters were also done with the Kolmogorov-Smirnov criterion. However, proportion of stems with no defects and mortality were only approximately normally distributed. Since arcsin transformation changed the results only slightly, these traits were left untransformed. The statistical significance of the effects of trial site, seed source (provenance) and block on the parameters studied was estimated using the analysis of variance. The linear model equation was defined as:
![]()
where yijk = the trait value for plot means in the ith trial of the jth seed source (provenance) in the kth block; μ = overall mean; Ti = the effect of trial site, i = 1…4; Pj = the effect of seed source (provenance), j = 1…7; Bk = the effect of block, k = 1…12; eijk = the experimental error.
Taking into account that lodgepole pine was considered as alternative to Scots pine the differences between seed sources/provenances (Scots pine vs. lodgepole pine) were analyzed by the Scheffe’s test. The analysis was performed using the ANOVA and post hoc comparison procedure. Simple regressions were employed to explain the relationships between the traits studied and average latitude of lodgepole pine material (parent seed trees in Canada) included in the Swedish seed orchards. The analysis was performed using the SAS statistical package (SAS/STAT User’s Guide 1999).
3 Results
The test site and seed source (provenance) had a highly significant (p < 0.05) effect on all growth and quality traits studied. Block was also a significant factor for all growth traits and stems with defects, but insignificant for stems with no defects and mortality (Table 3).
| Table 3. Analysis of variance of growth and quality traits of the all material studied. | ||||||||||
| Source | Growth traits | |||||||||
| Height, m | DBH, cm | Volume, dm3 | ||||||||
| df | MS | F-value | p-value | MS | F-value | p-value | MS | F-value | p-value | |
| Trial | 3 | 16.76 | 53.18 | <0.001 | 99.51 | 75.07 | <0.001 | 644.88 | 68.61 | <0.001 |
| Seed source (provenance) | 6 | 1.04 | 3.30 | 0.004 | 3.26 | 2.46 | 0.026 | 28.34 | 3.02 | 0.008 |
| Block | 10 | 1.20 | 3.82 | <0.001 | 5.65 | 4.27 | <0.001 | 29.92 | 3.18 | <0.001 |
| Error | 210 | 0.32 | - | - | 1.33 | 9.40 | ||||
| Quality traits | ||||||||||
| Stems with no defects | Stems with defects | Mortality | ||||||||
| df | MS | F-value | p-value | MS | F-value | p-value | MS | F-value | p-value | |
| Trial | 3 | 0.82 | 54.40 | <0.001 | 0.98 | 29.18 | <0.001 | 0.18 | 5.74 | <0.001 |
| Seed source (provenance) | 6 | 0.14 | 9.30 | <0.001 | 0.21 | 6.15 | <0.001 | 0.08 | 2.70 | 0.015 |
| Block | 10 | 0.02 | 1.15 | 0.327 | 0.10 | 3.11 | <0.001 | 0.02 | 0.56 | 0.842 |
| Error | 213 | 0.02 | 0.03 | 0.03 | ||||||
Lodgepole pine trees were taller (4–13%) than Scots pine trees except for the seed source of the southernmost origin Österby). The same results were also obtained for diameter growth (superiority was 4–13%) but the southernmost lodgepole pine seed source (Österby) had DBH under the standard (Table 4). The most important growth trait, stem volume was bigger (12–31%) for all lodgepole pine seed sources compared to local Scots pine except for the southernmost seed source (Österby) with stem volume under the standard. For all stem growth traits (height, DBH, and volume) the differences between lodgepole pine and local Scots pine were insignificant (Table 4).
| Table 4. The mean values and standard errors of growth s and quality traits of all material studied. | |||||||||||
| Seed source (provenance) | Growth traits | ||||||||||
| Height, m | DBH, cm | Volume, dm3 | |||||||||
| Mean | SE | p-value | Mean | SE | p-value | Mean | SE | p-value | |||
| Närlinge | 3.38 | 0.15 | 0.250 | 5.2 | 0.33 | 0.500 | 7.5 | 0.88 | 0.453 | ||
| Oppala | 3.36 | 0.14 | 0.320 | 5.0 | 0.26 | 0.868 | 6.8 | 0.66 | 0.932 | ||
| Skörserum | 3.33 | 0.13 | 0.437 | 5.1 | 0.32 | 0.671 | 7.6 | 0.92 | 0.375 | ||
| Larslund | 3.12 | 0.15 | 0.988 | 4.9 | 0.33 | 0.973 | 6.8 | 0.82 | 0.910 | ||
| Rumhult | 3.15 | 0.13 | 0.979 | 4.8 | 0.28 | 0.997 | 6.5 | 0.72 | 0.984 | ||
| Österby | 2.96 | 0.13 | 0.999 | 4.4 | 0.30 | 0.998 | 5.1 | 0.71 | 0.994 | ||
| Standard | 3.00 | 0.11 | - | 4.6 | 0.26 | - | 5.8 | 0.60 | - | ||
| Quality traits | |||||||||||
| Stems with no defects | p-value | Stems with defects | p-value | Mortality | p-value | ||||||
| Närlinge | 0.22 | 0.03 | <0.001 | 0.43 | 0.04 | 0.226 | 0.30 | 0.03 | 0.999 | ||
| Oppala | 0.22 | 0.03 | <0.001 | 0.47 | 0.04 | 0.035 | 0.25 | 0.03 | 0.898 | ||
| Skörserum | 0.19 | 0.03 | <0.001 | 0.47 | 0.04 | 0.034 | 0.28 | 0.03 | 0.992 | ||
| Larslund | 0.23 | 0.04 | 0.003 | 0.55 | 0.04 | <0.001 | 0.23 | 0.02 | 0.688 | ||
| Rumhult | 0.17 | 0.04 | <0.001 | 0.54 | 0.04 | <0.001 | 0.32 | 0.03 | 0.999 | ||
| Österby | 0.20 | 0.04 | 0.002 | 0.48 | 0.05 | 0.026 | 0.38 | 0.04 | 0.883 | ||
| Standard | 0.38 | 0.04 | - | 0.31 | 0.03 | - | 0.31 | 0.03 | - | ||
The share of stems without defects was significantly (p < 0.05) larger for Scots pine compared to that for lodgepole pine. Thereafter the proportion of stems with defects was lower for Scots pine compared to that for lodgepole pine, except for the seed source of the northernmost origin Närlinge (Table 4). The most common defects for both species were weak crookedness (proportion of severe crooked stems was less 1.0%.) and ramicorns (Table 5). The mortality was slightly higher for Scots pine than that for lodgepole pine, but differences were statistically insignificant (Table 4).
| Table 5. The percentagea of trees with different types of defects for lodgepole pine seed sources and local Scots pine (standard). | |||||
| Seed source (provenance) | Forksb | Ramicornsb | Spike knotsc | Crookednessb | |
| weak | severe | ||||
| Närlinge | 27.7 | 18.7 | 12.0 | 40.8 | 0.8 |
| Oppala | 32.6 | 13.5 | 11.1 | 42.4 | 0.4 |
| Skörserum | 29.4 | 25.7 | 10.5 | 34.1 | 0.3 |
| Larslund | 27.3 | 32.6 | 4.4 | 34.8 | 0.9 |
| Rumhult | 21.7 | 39.4 | 10.4 | 28.5 | - |
| Österby | 23.2 | 41.5 | 7.3 | 27.7 | 0.3 |
| Standard | 13.0 | 30.5 | 22.0 | 33.9 | 0.6 |
| a the percentages were calculated as share of stems with defects b forks, ramicorns and crooked trees may have also spike knots c straight stems with spike knots | |||||
The latitudinal range of the lodgepole pine origins was large enough for them to exhibit a clinal variation in parameters studied. The northern seed sources had better stem growth (height, DBH and volume), while the effect of latitude on the quality traits was insignificant (Fig. 2).
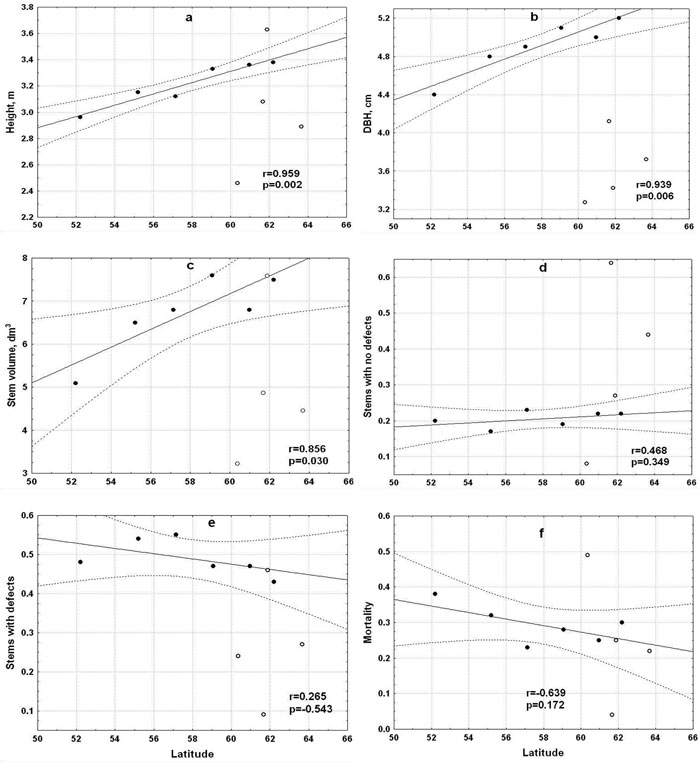
Fig. 2. Relationships between traits values (●) and average latitude of lodgepole pine material: a) height, b) diameter at breast height (DBH), c) volume, d) proportion of stems with no defects, e) proportion of stems with defects and f) mortality. Dotted lines represent 95% confidence intervals. Trait values of four standard Scots pine provenances (○). View larger in new window/tab
4 Discussion and conclusion
The six lodgepole pine seed sources planted together with local Scots pine provenances in four field trials in southern and central parts of the Komi Republic (Tables 1 and 2; Fig. 1) offered a good possibility to study mortality, stem growth and quality. It also gives some hints on the possibilities for using lodgepole pine in artificial reforestation in north-west Russia.
The tree vitality was about the same for lodgepole pine and Scots pine in Ukhta, Storojevsk, and Koygorodok field trials at the age of 4 years (Fedorkov and Turkin 2010), but at the age of 8 years tree vitality of Scots pine was better compared to lodgepole pine owing to frost damage in the end of winter and beginning of spring 2010 (Fedorkov 2012).
Survival is a complex character reflecting combined effects of all events causing injuries and die-back in pine populations (Persson et al. 2010). The results of this study are consistent with conclusions presented in Elfving et al. (2001) about higher survival of lodgepole pine during establishment but higher mortality after the first thinning compared to Scots pine (Table 4). One of the reasons is higher resistance of lodgepole pine against some fungi that causes severe problems in Scots pine, for example, snow blight (Phacidium infestans Karst.) (Elfving et al. 2001). However, Varmola et al. (2000) found in a series of field trials established in Finland that lodgepole pine survival at the age of 13–14 years was in average 14% lower in comparison with Scots pine. Opposite results were obtained in northern Finland by Ruotsalainen and Velling (1993) where survival of lodgepole pine was better compare to Scots pine and clearly dependent on provenance at age about 20 years.
High occurrence of stem defects among lodgepole pine trees of southern origins can be partly attributed to later cessation of shoot growth. It is well known that late growth cessation in autumn is strongly related to the risk of frost damage (Aitken and Hannerz 2001). Phenology study based on the same material in the Komi Republic revealed such later cessation of shoot growth of lodgepole pine vs. Scots pine and a latitudinal cline among lodgepole pine origins (Fedorkov 2010). A higher proportion of damaged trees with killed leader shoots for lodgepole pine seed sources of southern origins was recorded in fields trials studied in 2011 (Fedorkov 2012).
The volume superiority of lodgepole pine of northern origins (17–31%) revealed in our study is close to results obtained in Sweden (Elfving et al. 2001) and south Karelia (Raevsky and Pekkoev 2013) where lodgepole pine trees were superior to Scots pine trees in term of stem volume to 36 and 38%, correspondingly. The stem growth superiority of northern lodgepole pine origins in comparison with southern ones correspond well with study of Ruotsalainen and Velling (1993). Statistical insignificance of this superiority (Table 4) may be partly explained by row-plot design of field experiments employed in our study which is less efficient in comparison with single-tree plot design. Reasons for the superior growth of lodgepole pine under boreal condition may be an earlier start of growth in spring and a lower required heat sum to start shoot elongation in comparison to Scots pine (Fedorkov 2010).
In conclusion, Scots pine stem quality was better than that of lodgepole pine but, on the contrary, lodgepole pine stem growth was faster except for the southernmost seed source Österby. Suitability of lodgepole pine for use in forest cultivation in north-west Russia depends on population origin. Undoubtedly, lodgepole pine seed sources of southern origins are less suitable for artificial reforestation in the Komi Republic.
Acknowledgements
We thank Svenska Skogsplantor AB and SkogForsk for the Swedish lodgepole pine seed orchards’ seed lots provided. Many thanks go to the personnel of Forest Committee of the Komi Republic for seedlings production and planting of field trials. We would like to acknowledge anonymous reviewer and editor for valuable comments and suggestions on the manuscript.
References
Aitken S., Hannerz M. (2001). Genecology and gene resource management strategies for conifer cold hardiness. In: Bigras F.I., Colombo S.J. (eds.). Conifer cold hardiness. Kluwer Academic Publishers. p. 23–53.
Backlund I., Bergsten U. (2012). Biomass production of dense direct-seeded lodgepole pine (Pinus contorta) at short rotation periods. Silva Fennica 46(4):609–623. http://dx.doi.org/10.14214/sf.914.
Elfving B., Ericsson T., Rosvall O. (2001). The introduction of lodgepole pine for wood production in Sweden – a review. Forest Ecology and Management 141(1–2):15–20. http://dx.doi.org/10.1016/S0378-1127(00)00485-0.
Elfving B., Norgren O. (1993). Volume yield superiority of lodgepole pine compared to Scots pine in Sweden. In: Lindgren D. (ed.). Proceedings of a meeting IUFRO Working Party: Pinus contorta from untamed forest to domesticated crop. Swedish University of Agricultural Sciences, Umeå. p. 69–80.
Ericsson T. (1994). Lodgepole pine (Pinus contorta var. latifolia) breeding in Sweden – results and prospects based on early evaluations. Swedish University of Agricultural Sciences, Umeå. Dissertation.
Fedorkov A. (2010). Variation in shoot elongation patterns in Pinus contorta and Pinus sylvestris in north-west Russia. Scandinavian Journal of Forest Research 25(3): 208–212. http://dx.doi.org/10.1080/02827581.2010.491229.
Fedorkov A. (2012). Swedish lodgepole pine seed orchard crops tested in north-west Russia. Scandinavian Journal of Forest Research 27(7): 675–680. http://dx.doi.org/10.1080/02827581.2012.690042.
Fedorkov A.L, Turkin A.A. (2010). Экспериментальные культуры сосны скрученной в Республике Коми. [Test of shore pine in the Republic of Komi]. Russian Journal of Forest Science 1:70–74. [In Russian with English summary].
Feklistov P.A, Birjkov S.Y, Fedjev A.L. (2008). Сравнительные эколого-биологические особенности сосны скрученной и обыкновенной в северной подзоне европейской тайги. [The biological properties lodgepole pine and Scots pine in northern taiga zone]. Archangelsk State Technical University, Archangelsk, Russian Federation. 118 p. [In Russian].
Galenko E.P. (1999). Климатические условия и фитоклиматический режим. [Climatic conditions and phytoclimatic regime]. Moskow, Russian Federation. 332 p. [In Russian].
Hansson P., Karlman M. (1997). Survival, height and health status of 20-year-old Pinus sylvestris and Pinus contorta after different scarification treatments in a harsh boreal climate. Scandinavian Journal of Forest Research 12(4): 340–350. http://dx.doi.org/10.1080/02827589709355421.
Lindgren D. (1993). Breeding Pinus contorta in different countries. In: Lindgren D. (ed.). Proceedings of a meeting IUFRO Working Party: Pinus contorta from untamed forest to domesticated crop. Swedish University of Agricultural Sciences, Umeå. p. 264–270.
Magalska L., Howe G.T. (2014). Genetic and environmental control of Douglas-fir stem defects. Forest Ecology and Management 318: 228–238. http://dx.doi.org/10.1016/j.foreco.2014.01.002.
Nilsson P., Cory N. (2010). Skogsdata 2010, aktuella uppgifter om de svenska skogarna från Riksskogstaxeringen. [Forestry statistics 2010]. Sveriges Lantbruksuniversitet, Umeå. 119 p. [In Swedish with English summary].
Owens J.N. (2006). The reproductive biology of lodgepole pine. Forest Genetics Council of British Columbia, Canada. 66 p. http://www.fgcouncil.bc.ca/ExtNote7-Final-web.pdf. [Cited 10 June 2016].
Persson T., Andersson B., Ericsson T. (2010). Relationship between autumn cold hardiness and field performance in northern Pinus sylvestris. Silva Fennica 44(2): 255–266. http://dx.doi.org/10.14214/sf.152.
Raevsky B.V. (2010). Ход роста смешанных культур сосны скрученной и сосны обыкновенной в южной Карелии. [Growth and development of lodgepole pine and Scots pine provenances in mixed plantation in south Karelia]. Notes of Komi Science Center 1: 31–38. [In Russian].
Raevsky B.V., Pekkoev A.N. (2013). Some perspectives of lodgepole pine planting in South Karelia. Paper presented at: Innovations and technologies in forestry-2013, 3rd International meeting, St-Petersburg, Russian Federation. p. 182–190. [In Russian with English summary].
Ruotsalainen S., Velling P. (1993). Pinus contorta in northern Finland – first 20 years. In: Lindgren D. (ed.). Proceedings of a meeting IUFRO Working Party: Pinus contorta from untamed forest to domesticated crop. Swedish University of Agricultural Sciences, Umeå. p. 122–136.
SAS Institute (1999). SAS/STAT user’s guide. Cary (NC), SAS Institute.
Tjurin E.G. (1972). Таблицы хода роста смешанных сосново-берёзовых молодняков Коми АССР и объёмы их стволов. [Growth and yield tables and stem volumes of Scots pine and birch in mixed stands]. Северо-Западное книжное издательство, Vologda. 25 p. [In Russian].
Varmola M., Salminen H., Rikala R., Kerkelä M. (2000). Survival and early development of lodgepole pine. Scandinavian Journal of Forest Research 15(4): 410–423. http://dx.doi.org/10.1080/028275800750172619.
Total of 22 references.
