Amira Fetouab,
Nicole J. Fenton,
Nelson Thiffault  ,
Martin Barrette
,
Martin Barrette
Planting density and mechanical site preparation effects on understory composition, functional diversity and planted black spruce growth in boreal forests
Fetouab A., Fenton N. J., Thiffault N., Barrette M. (2024). Planting density and mechanical site preparation effects on understory composition, functional diversity and planted black spruce growth in boreal forests. Silva Fennica vol. 58 no. 2 article id 23029. https://doi.org/10.14214/sf.23029
Highlights
- Mechanical site preparation and planting density interacted to influence understory diversity and composition in plowed plots
- Careful logging alone resulted in higher density of naturally established conifers
- Planting density and site preparation alone had no effect on tree height and diameter
- Findings provide insights for managing paludified forests and enhancing forest ecosystem functions.
Abstract
Mechanical site preparation (MSP) is used prior to planting to control competing vegetation and enhance soil conditions, particularly in areas prone to paludification. Tree planting density can be adapted to the management context and objectives, as it influences yield and wood quality. However, the combined effects of MSP and planting density on understory vegetation composition, functional traits, and diversity remain uncertain. We thus conducted a study in the Clay Belt region of northwestern Quebec, Canada. After careful logging, the study area was divided into nine sites, each receiving one of three treatments: plowing, disc trenching, or no preparation. Sites were further divided into two, with black spruce (Picea mariana [Mill.] Britton, Sterns & Poggenb.) seedlings planted at either a low planting density of 1100 seedlings ha-1 or a high planting density of 2500 seedlings ha-1. After nine years, we assessed understory composition, diversity, key functional traits, sapling density and growth of planted trees. Careful logging alone led to a higher density of naturally established conifers compared to plowing or disc trenching. The interaction between planting density and MSP significantly influenced understory diversity and composition in plowed plots. Understory composition was affected by the soil C/N ratio, coniferous species, and deciduous species density. The growth of black spruce was notably enhanced with higher planting density in the plow treatment only. Neither planting density nor MSP alone affected tree height and diameter. Our results suggest that combining plowing with high-density planting can enhance stand growth and improve forest productivity. These findings guide future research on paludified forests.
Keywords
silviculture;
paludification;
reforestation;
plantation establishment;
community ecology;
functional ecology
- Fetouab, Institute for Forest Research and Centre for Forest Research, Université du Québec en Abitibi-Témiscamingue, 445 boul. de l’Université, Rouyn-Noranda, QC J9X 5E4, Canada E-mail amira.fetouab@uqat.ca
-
Fenton,
Institute for Forest Research and Centre for Forest Research, Université du Québec en Abitibi-Témiscamingue, 445 boul. de l’Université, Rouyn-Noranda, QC J9X 5E4, Canada
 https://orcid.org/0000-0002-3782-2361
E-mail
nicole.fenton@uqat.ca
https://orcid.org/0000-0002-3782-2361
E-mail
nicole.fenton@uqat.ca
-
Thiffault,
Institute for Forest Research and Centre for Forest Research, Université du Québec en Abitibi-Témiscamingue, 445 boul. de l’Université, Rouyn-Noranda, QC J9X 5E4, Canada; Canadian Wood Fibre Centre, Natural Resources Canada, 1055 du P.E.P.S, P.O. Box 10380, Sainte-Foy Stn, Québec, QC G1V 4C7, Canada
https://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@canada.ca
-
Barrette,
Institute for Forest Research and Centre for Forest Research, Université du Québec en Abitibi-Témiscamingue, 445 boul. de l’Université, Rouyn-Noranda, QC J9X 5E4, Canada; Direction de la recherche forestière, Ministère des Ressources naturelles et des Forêts, 2700 rue Einstein, Québec, QC G1P 3W8, Canada
https://orcid.org/0000-0001-5937-382X
E-mail
martin.barrette@mffp.gouv.qc.ca
Received 28 June 2023 Accepted 16 March 2024 Published 21 March 2024
Views 97545
Available at https://doi.org/10.14214/sf.23029 | Download PDF

Supplementary Files
1 Introduction
Global wood production strategies are gradually shifting from natural forests to plantation forestry. Plantation forestry has the potential to increase forest productivity when compared with natural forests. The high yields of planted forests are indeed an opportunity for achieving wood fibre production and allowing the provision of other ecosystem services (Paquette and Messier 2010). Plantation success depends on interdependent decisions and actions (Thiffault et al. 2023). For example, planting density is an important factor affecting forest production through its effects on tree survival and growth, wood yield, and biodiversity (Benomar et al. 2012). Planting density has the potential to influence understory species composition and richness, notably by affecting understory and soil temperatures (Hedwall et al. 2015). This can affect the presence of species that compete with forest regeneration. Indeed, rapid canopy closure could limit the growth of competing plants and thus reduce their negative effect on forest productivity (Moroni et al. 2009).
Mechanical site preparation (MSP) before planting is one of the most common practices in Canada’s forests to ensure successful regeneration of harvested sites (Sutherland and Foreman 1995). The main benefits of MSP are to reduce competition from other species colonizing regeneration areas (Johansson et al. 2013; Mallik and Kravchenko 2016) and to increase the availability of resources, which enhances seedling survival (Heiskanen et al. 2013; Luoranen et al. 2017). In boreal forests, MSP has been shown to increase the growth of planted spruce (Picea spp.) by increasing nutrient availability (Sutherland and Foreman 2000) and N mineralization and uptake due to warmer soils and improved soil moisture (Johansson 1994; Nordborg et al. 2003). Therefore, MSP is a critical tool for creating suitable planting microsites on boreal harvested sites (Wallertz et al. 2018). Moreover, MSP can influence understory functional composition and diversity. For example, in a field study conducted in boreal Quebec (Canada), disc scarification led to the emergence of species that were not present in the control plots: the treatment promoted the growth of intolerant, early-successional, seed bank species and noticeably decreased the prevalence of ericaceous shrubs (Thiffault and Jobidon 2006). Eighteen years after treatment, ericaceous shrubs in scarified plots had lower specific rhizome mass, lower N use efficiency, lower tannin and higher N concentrations in leaves and litter than in non-scarified plots (Reicis et al. 2023).
In understanding the multifaceted role of planted forests, it’s important to recognize their contribution beyond wood production and economic gains, including their significance in conserving biodiversity (Bremer and Farley 2010). This aspect becomes particularly evident in conifer plantations, where the environmental manipulation – altering light availability and litter layer characteristics – affects understory plant habitats in temperate and boreal forests (Saetre et al. 1997; Barbier et al. 2008). Furthermore, the implementation of MSP treatments plays a crucial role. Some MSP practices, by managing organic layers and fallen woody debris, not only help in controlling competing species and establishing planted seedling but also enhance site productivity (Bock and Van Rees 2002; Ramovs and Roberts 2003; Powers 2006). MSP practices must be used in a way to maintain ecosystem values including understory plants, whose importance extends to maintaining structural and functional integrity. Their roles in soil nutrient cycling, carbon storage, and as indicators of site quality have been highlighted (Nilsson and Wardle 2005; Gilliam 2007; Zhou et al. 2018; Wei et al. 2021). Hence, we need to understand the interactions between planted trees, silviculture treatments, and the understory in managed forests so that ecosystem dynamics and functions are maintained (Gilliam and Roberts 2003). Few studies have investigated stand properties (i.e., growth, understory composition and function) under different planting densities and after different site preparation techniques.
We aimed to evaluate the main and interacting effects of planting density and MSP on understory composition and functional traits, and the growth of planted black spruce (Picea mariana [Mill.] Britton, Sterns & Poggenb.) in forest sites prone to paludification. Functional traits are measurable characteristics of plants that influence their fitness and growth, and are an effective way to examine the relationship between understory vegetation and ecosystem function (Violle et al. 2007; Wei et al. 2021). In our study, we have specifically focused on distinct functional traits for bryophytes and vascular plants, reflecting their roles in ecosystem functioning. For bryophytes, we measured traits related to colony characteristics that influence ecosystem processes, including colony density, water content at field capacity, water absorption capacity, and water percolation (Lavorel and Garnier 2002; Cornelissen et al. 2007). These traits are important in understanding how bryophyte colonies affect water dynamics and nutrient cycling within forest ecosystems. For vascular plants, our attention was on three functional traits: morphology type, maximum height, and specific leaf area. These traits provide insights into the adaptive strategies of vascular plants, influencing aspects such as light interception, growth patterns, and competitive interactions (Garnier et al. 2004). Morphology type helps in understanding plant structure and its ecological role, while maximum height is indicative of competitive ability for light. Specific leaf area, a key trait related to photosynthetic capacity and resource use efficiency, offers insights into how vascular plants respond to environmental conditions and management practices. By examining these specific traits in both bryophytes and vascular plants, we aimed to understand how changes in understory composition, driven by varying planting densities and MSP treatments, can impact ecosystem properties such as water retention and nutrient cycling. This approach allows us to assess the broader implications of forestry practices on ecosystem health and functionality.
Our specific objectives were to evaluate the sole and combined effects of planting density and MSP on (1) environmental conditions, specifically soil chemistry, canopy openness, the density of coniferous and deciduous vegetation and vegetation cover, (2) understory composition (2a) and diversity (2b), (3) bryophyte and vascular plant functional traits, and (4) the growth of planted saplings. We expected significant interactions between the two treatments to influence sapling growth, understory composition, and functional traits. Specifically, we hypothesized that increased planting density and more intensive MSP would significantly alter soil chemistry, mediated by changes in canopy openness and the densities of coniferous and deciduous species. These alterations were expected to influence understory composition and functional traits, subsequently positively affecting the growth of saplings compared to control conditions.
2 Materials and methods
2.1 Study area
The study was conducted in the black spruce–feather moss bioclimatic domain (Saucier et al. 2009) located in the Clay Belt region of north-western Quebec, Canada, which is covered by a fine-textured clay deposit left by the proglacial lakes Barlow and Ojibway (Vincent and Hardy 2011). The soil of the region is dominated by compacted clay and is subdivided by sand and gravel deposits of the Harricana moraine. The study site is located approximately 200 km north-east of Rouyn-Noranda (49°27´N, 78°31´W) (Fig. 1). The site is prone to paludification with imperfect drainage, low topographic relief, and a cold, wet climate (Fenton et al. 2005). The average annual temperature approximates 0°C and the average annual precipitation is 909 mm (Joutel weather station; Environment Canada 2021).
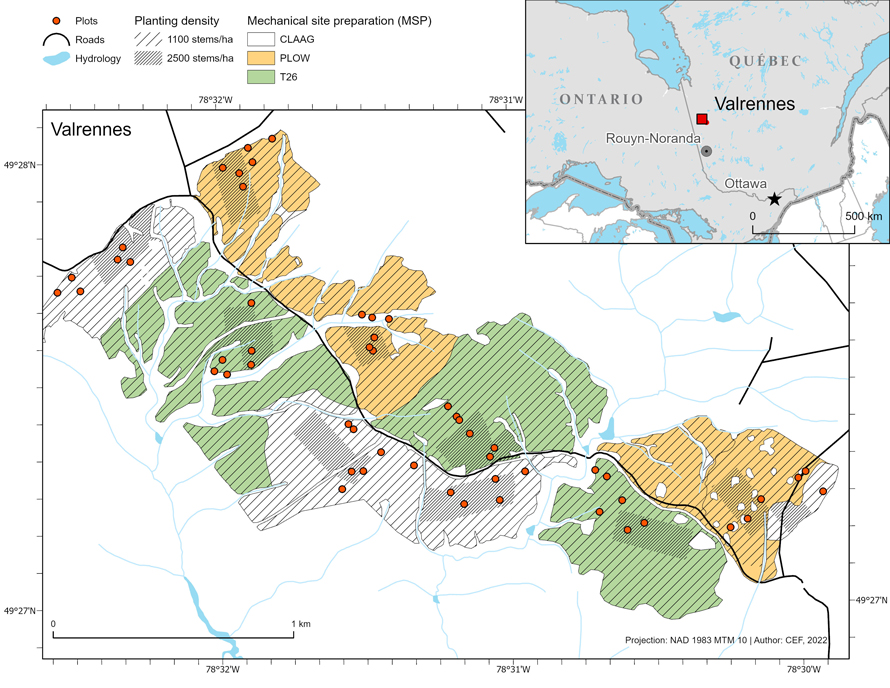
Fig. 1. Location of the study region and experimental site in Quebec (Canada), showing the distribution of the silvicultural treatments (careful logging around advanced growth (CLAAG), plowing and T26 scarification) with location of the low (1100 seedlings ha–1) and high (2500 seedlings ha–1) planting density of black spruce (Picea mariana) areas within each site. View larger in new window/tab.
Before harvest, the site was dominated by mature black spruce, in mixture with a lower proportion of trembling aspen (Populus tremuloides Michx), tamarack (Larix laricina [Du Roi] K. Koch), balsam fir (Abies balsamea [L.] Mill.), and white birch (Betula papyrifera Marshall). The shrub cover was dominated by Labrador tea (Rhododendron groenlandicum [Oeder] Kron & Judd) and sheep laurel (Kalmia angustifolia L.). The forest floor was covered by Sphagnum mosses (including Sphagnum magellanicum Brid.; S. russowii Wamst.; and S. capillifolium [Ehrh.]) and feather mosses (Pleurozium schreberi [Brid.] Mitt).
2.2 Study design and treatments
The field experiment was established on nine sites averaging 32 ha each (Henneb et al. 2019). In the fall of 2010, all sites were harvested using careful logging around advanced growth (CLAAG). In 2011, six sites out of the nine were randomly selected to be subjected to one of two MSP treatments: three were treated using a forest plow (Plow), and three were treated by disc trenching using a T26 scarifier (Bracke Forest AB, Bräcke, Sweden) (Fig. 1). The three-remaining sites were kept as controls (harvested without MSP). In early summer 2012, each site was divided into two sub-sites that were planted with black spruce seedlings at a low (1100 seedlings ha–1) or high (2500 seedlings ha–1) planting density (Fig. 1). Seedlings were produced from a local seed source in containers of 45 cells of 110 cm3 each; seedlings averaged 20 cm in height at the time of planting. After planting in 2012, 120 circular sampling plots (each with a radius of 11.28 m) were randomly distributed across the entire cut-block (Henneb et al. 2019). Within each plot, five 1 m2 quadrats were established at the north, east, south, west, and center positions. We selected and tagged one planted seedling in each quadrat, or the nearest seedling to the quadrat if none were inside.
2.3 Data collection
Tagged seedlings were measured for initial dimensions after planting, and again in 2016. In 2020 (the ninth growing season after planting), we revisited three sampling plots (n = 54) randomly selected in each planting density subsite, as it was not possible to visit all of the 120 plots due to time constraints. During the summer and autumn, we collected data related to understory vegetation diversity and function in two randomly selected 1 m2 quadrats (n = 108) in each selected sampling plot. All vascular plants and macro bryophyte species were identified in the field, except for Sphagnum spp, which were identified in the lab (Faubert 2013).
To determine the functional traits of vascular plants and bryophytes, percent cover was used as a proxy for biomass, and we used the community weighted mean approach, i.e., we summed the cover of the most abundant species in the quadrats until they represented at least 80% of the biomass for each of these layers (Garnier et al. 2004). For bryophytes, we measured the functional traits related to how bryophyte colony characteristics influence ecosystem functioning (Lavorel and Garnier 2002; Cornelissen et al. 2007). We selected four effect traits: colony density (mg cm–3), water content at field capacity (%), water absorption capacity (ml cm–3), and water percolation (% cm–3). Traits were measured on 10 cm diameter colonies collected from each quadrat. In every sampled colony, the 10 cm sample was carefully removed to keep the colony structure intact. In the lab, colony samples were re-humidified before measurements by watering, and then left for 12 h in a container with water for full rehydration and then weighed. Colony density was calculated using water-saturated bryophyte mass and volume, determined by measuring the colony. After 5 days of drying, the bryophyte colony samples were placed in pots that contained 300 ml of water. To measure water content at field capacity, water that was not absorbed by the sample after 12 h of soaking was collected in a 500 ml graduated container. The colony samples were then turned 10 times in a salad spinner to remove water that was not retained internally by the moss. Mass was recorded and the procedure was repeated until a stable weight (+/– 0.1 g) was reached. Samples were then weighed to obtain mass at field capacity, and the field capacity (% weight) was determined. For percolation, water that percolated through the colony was collected and measured. The percentage of water percolated was calculated and standardized by the volume of the colonies to obtain water percolation through the bryophyte colony by volume. Finally, the water was then added to the 500 ml graduated container. By subtracting this new volume from the initial 300 ml, we obtained the water absorption capacity of the samples.
For vascular plants, we used the Traits of Plants in Canada (TOPIC) database (Aubin et al. 2020) to obtain values for three functional traits: morphology type (MOR) (abundance of herbaceous and small shrubs), maximum height (HT) (cm), and specific leaf area (SLA) (m2 kg–1). These traits can be used to assess the impact of community changes on ecosystem properties (Garnier et al. 2004).
To measure environmental conditions and planted tree growth, the north, east, south, and west quadrats were selected for measurement in each of the same 54 plots, thus totaling 216 quadrats in total. Quadrats with dead tagged trees were removed from the analysis; a total of 180 quadrats remained, distributed as follows: 25 in CLAAG – High planting density, 32 in CLAAG – Low planting density, 29 in Plow – High planting density, 30 in Plow – Low planting density, 30 in T26 – High planting density, and 34 in T26 – Low planting density. We collected three soil samples at a 20 cm depth and combined them into one composite soil sample per quadrat for chemical analyses. Soil samples were transported to the lab, air-dried, sieved, and analyzed for determination of total carbon and nitrogen concentrations by dry combustion (LECO CNS-2000, LECO Corporation, TruMac CNS). We measured canopy openness at a 0.3 m height in the center of each quadrat in the four cardinal points with a spherical crown densiometer (Forestry Suppliers, Convex Model A). We also inventoried coniferous, deciduous, and shrub species (stems ha–1) taller than 1 m in height in a 40 m2 circular sampling plot centered on the tagged planted black spruce sapling of each quadrat.
We measured the ground-level diameter and height of the tagged trees and calculated 4-year height growth using 2016 and 2020 data. In fall 2020, needles were harvested from the top third of the planted trees; six current year twigs were collected from each tree. The needles from collected twigs were oven-dried at 50 °C for 24 h. Samples from each quadrat were combined into one composite sample and analyzed for foliar N and C concentrations.
2.4 Statistical analysis
All analyses were conducted in the R statistical environment v.4.1.0 (R Core Team 2020). First, we used linear mixed models (lmer function from the lme4 package; Bates et al. 2015), the sampling plots (nested in sites) were included as random factors, to evaluate the effects of MSP, planting density, and their interaction on conifer density (natural and planted), deciduous species density, shrub density, vegetation cover and soil C/N ratio (Objective 1). The statistical model is as follow:
![]()
where:
Y = response variable,
T = MSP treatment,
D = planting density treatment,
P = sampling plot (random effect) nested in the site factor S.
We calculated estimated marginal means and conducted Tukey’s honestly significant difference (HSD) pairwise comparisons with the emmeans package (Lenth et al. 2022). We used α ≤ 0.05 as a threshold for significance when comparing treatments. Using the same approach, we evaluated treatment effects on bryophytes and vascular plant functional trait values (Objective 3), and on sapling height growth, diameter, and foliar N and C concentrations (Objective 4). Linear correlation was used to examine the relationship between functional traits and growth (Objective 3).
Second, permutational multivariate analysis of variance with 999 permutations (Permanova, “adonis” function from the vegan package, Oksanen et al. 2020) were run to test the effects of planting density, MSP, and their interaction on understory composition vascular plants and bryophytes (Objective 2a). The plot term nested in site was used in the “Strata” function to take into account the spatial structure in the data in the Permanova analysis and non-significant interaction terms were removed in final analysis. Results were visualized using principal coordinates analysis (PCoA, “PCOA” function in the vegan package) with Bray–Curtis dissimilarity distance and the Cailliez correction (package ape; Paradis and Schliep 2019). Convex hulls were used to highlight the plots from different treatments in each planting density type. The “envfit” function in vegan was then used to fit vectors of environmental conditions (conifer density, deciduous density, shrub density, vegetation cover and soil C/N ratio) onto the ordinations to illustrate how environmental conditions in the understory relate to community composition. In each plot, we calculated % cover (for bryophytes), Simpson’s dominance index (for vascular plants) and Shannon-Wiener diversity index using the vegan package (Oksanen et al. 2020). We used linear mixed models followed by multiple means comparisons using emmeans (Objective 2b).
3 Results
3.1 Environmental conditions
Planting density and MSP affected all variables we measured to describe the understory environment, independently or in interaction (Table 1). Coniferous and deciduous density, and vegetation cover were only affected by MSP. There was a higher density of naturally regenerated conifers in CLAAG than Plow, but there was no difference with the T26 treatment (Fig. 2A). The density of deciduous trees was higher also in CLAAG than T26 but there was no difference with the treatment Plow (Fig. 2B). Vegetation cover was higher in CLAAG than in the other treatments (Fig. 2C). There was no significant effect of planting density on natural conifer density, deciduous density, and vegetation cover.
| Table 1. Results from linear mixed model for the main and interacting effects of planting density of black spruce (Picea mariana) and mechanical site preparation on environmental conditions and understory diversity. Df = degrees of freedom. Bold indicates significance at α ≤ 0.05. | ||||||
| Response variable | Treatment effects | |||||
| Planting density | Mechanical site preparation (MSP) | Planting density × MSP | ||||
| Df | p-value | Df | p-value | Df | p-value | |
| Environmental conditions | ||||||
| Conifer density (stems ha–1) | 1 | 0.28 | 2 | 0.02 | 2 | 0.87 |
| Deciduous species density (stems ha–1) | 1 | 0.56 | 2 | 0.036 | 2 | 0.3 |
| Shrub density (stems ha–1) | 1 | 0.55 | 2 | 0.65 | 2 | 0.05 |
| Vegetation cover (%) | 1 | 0.5 | 2 | 0.01 | 2 | 0.96 |
| C/N ratio | 1 | 0.003 | 2 | <0.001 | 2 | 0.3 |
| Understory diversity | ||||||
| Vascular plant dominance (Simpson index) | 1 | 0.3 | 2 | 0.9 | 2 | 0.65 |
| Vascular plant diversity (Shannon index) | 1 | 0.07 | 2 | 0.91 | 2 | 0.09 |
| Bryophyte cover (%) | 1 | 0.002 | 2 | 0.11 | 2 | <0.001 |
| Bryophyte diversity (Shannon-Wiener index) | 1 | 0.15 | 2 | 0.2 | 2 | 0.46 |
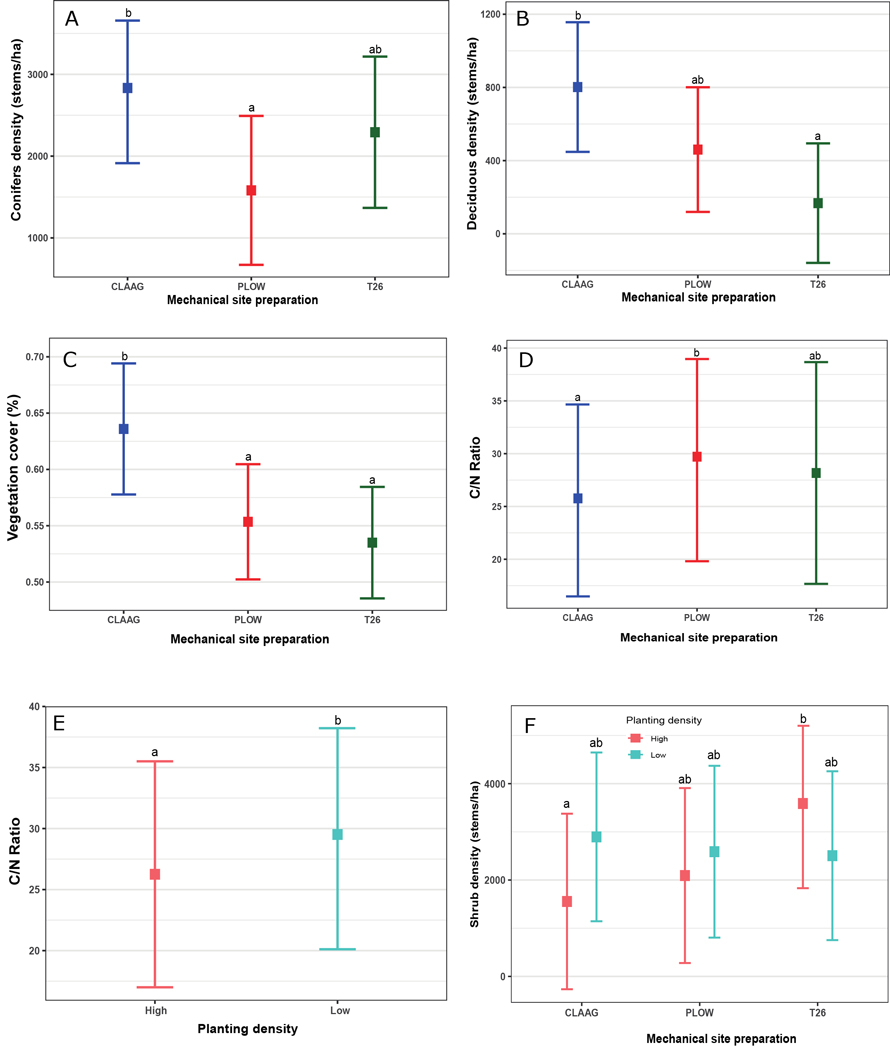
Fig. 2. Effects of planting density of black spruce (Picea mariana) (Low = 1100 seedlings ha–1; High = 2500 seedlings ha–1) and mechanical site preparation (CLAAG = careful logging around advanced growth; Plow = plowing; T26 = disk trenching scarification) on naturally regenerated conifer density (A), deciduous species density (B); vegetation cover (C); and soil C/N ratio (D, E). Panel F illustrates the interacting effects of mechanical site preparation and planting density on shrub density. Data are presented as estimated marginal means with 95% confidence intervals. Means followed by the same letter are not significatively different at α ≤ 0.05 based on linear mixed models with Tukey HSD pairwise comparisons. (Plots CLAAG low, n = 32; plots CLAAG high, n = 25; plots Plow low, n = 30; plots Plow high, n = 29; plots T26 low, n = 34; plots T26 high, n = 30). View larger in new window/tab.
The soil C/N ratio was influenced by planting density and MSP independently, with lower values measured in low planting density and CLAAG plots (Figs. 2D &E). Plowed plots had significantly higher values than the CLAAG treatment, and T26 was intermediate (Table 1). The interaction between planting density and MSP was significant for shrub density (Table 1), with higher shrub density in plots that received the T26 treatment and high planting density than in CLAAG with high planting density, and no difference with the other treatments (Fig. 2F).
3.2 Understory composition
Understory vascular plant composition was affected by MSP and the interaction with planting density (Table 2). The results of the pairwise Permanova showed that vascular plant communities were significantly different in high planting density plots in the three treatments, but not in low planting density plots (Table 3). The principal coordinates analysis (PCoA) also supported the results of the Permanova (Fig. 3). Overall, the plots on the upper side of PCoA were dominated by Plow in both high and low planting density plots. The lower side was dominated by T26 and CLAAG. However, in low planting density plots, the three treatments overlapped considerably, while in high planting density plots, the shape and position of the ellipses differed among treatments (Fig. 3). Soil C/N ratio and deciduous species density had the strongest relationship with the community pattern for vascular plants on PCoA 1, while conifer density was strongly associated with PCoA 2 (Fig. 3). There were 3 species (Chamaedaphne calyculata [L.] Moench, Scirpus cyperinus [Linnaeus] Kunth and Equisetum arvense Linnaeus) associated positively with C/N ratio on the left of PCoA 1, and species associated with deciduous density on the right (e.g., Ribes triste Pallas and Rubus pubescens Rafinesque) (Fig. 3).
| Table 2. Influence of planting density of black spruce (Picea mariana) and mechanical site preparation and their interaction on vascular plant and bryophytes dissimilarity using permutational multivariate analysis of variance (Permanova) based on the Bray–Curtis dissimilarity index. Df = degrees of freedom. Bold indicates significance at α ≤ 0.05. SS = sum of squares. | ||||||
| Assemblage | Treatment | Df | SS | F | R2 | p-value |
| Vascular plant | Planting density | 1 | 0.33 | 1.24 | 0.011 | 0.316 |
| Mechanical site preparation (MSP) | 2 | 2.13 | 4.02 | 0.07 | 0.001 | |
| Planting × MSP | 2 | 0.84 | 1.58 | 0.02 | 0.048 | |
| Residual | 98 | 25.9 | 0.89 | |||
| Bryophytes | Planting density | 1 | 0.63 | 2.83 | 0.026 | 0.036 |
| Mechanical site preparation (MSP) | 2 | 0.81 | 1.81 | 0.034 | 0.008 | |
| Planting × MSP | 2 | 1.14 | 2.56 | 0.048 | 0.016 | |
| Residual 1 | 94 | 21.10 | 0.89 | |||
| 1 four plots without bryophytes | ||||||
| Table 3. Results from post-hoc pairwise tests for vascular plant and bryophytes between mechanical site preparation treatments after planting of black spruce (Picea mariana) (CLAAG = careful logging around advanced growth; Plow = plowing; T26 = disk trenching scarification; High = planting density of 2500 seedlings ha–1; Low = planting density of 1100 seedlings ha–1). Df = degrees of freedom; SS = sum of squares. Bold indicates significance at α = 0.05. | |||||||||
| Pairs | Df | SS | F | R2 | p-value | adjusted p | |||
| Vascular plants | |||||||||
| CLAAG | vs | PLOW | High | 1 | 1.01 | 3.97 | 0.12 | 0.001 | 0.005 |
| CLAAG | vs | T26 | High | 1 | 0.49 | 2.15 | 0.07 | 0.019 | 0.038 |
| PLOW | vs | T26 | High | 1 | 0.51 | 2.05 | 0.06 | 0.030 | 0.005 |
| CLAAG | vs | PLOW | Low | 1 | 0.64 | 2.16 | 0.05 | 0.012 | 0.035 |
| CLAAG | vs | T26 | Low | 1 | 0.47 | 1.74 | 0.04 | 0.083 | 0.13 |
| PLOW | vs | T26 | Low | 1 | 1.27 | 4.74 | 0.11 | 0.001 | 0.006 |
| Bryophytes | |||||||||
| CLAAG | vs | PLOW | High | 1 | 0.76 | 3.12 | 0.10 | 0.028 | 0.1 |
| CLAAG | vs | T26 | High | 1 | 1.29 | 6.33 | 0.18 | 0.002 | 0.02 |
| PLOW | vs | T26 | High | 1 | 0.20 | 0.71 | 0.02 | 0.605 | 0.68 |
| CLAAG | vs | PLOW | Low | 1 | 0.18 | 0.75 | 0.02 | 0.564 | 0.64 |
| CLAAG | vs | T26 | Low | 1 | 0.08 | 0.47 | 0.01 | 0.658 | 0.7 |
| PLOW | vs | T26 | Low | 1 | 0.42 | 2.08 | 0.05 | 0.090 | 0.26 |
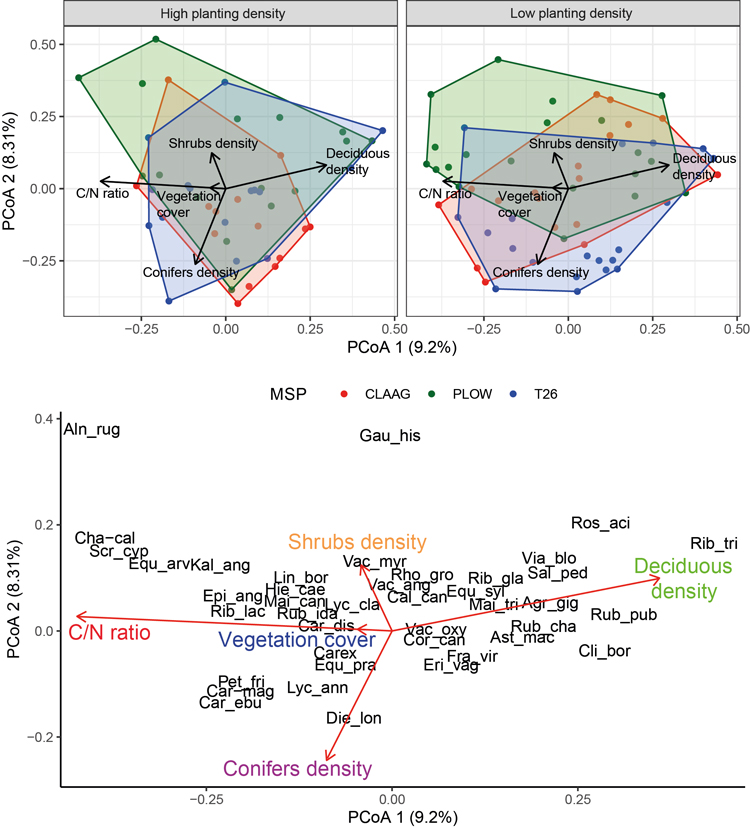
Fig. 3. Principal coordinates analysis (PCoA) for the vascular plant composition using the Bray-Curtis dissimilarity index and a Cailliez correction. Species names and environmental conditions were added by correlation to the axes. Colors in convex hull indicate different mechanical site preparation (MSP) treatments. See Supplementary file S1 for full species names.
When analyzing bryophyte community composition, the results of the Permanova showed significant differences in bryophyte composition associated with the interaction between planting density and MSP (Table 2). The results of pairwise Permanova showed that bryophyte communities differed only in high planting density with CLAAG and T26 (Table 3). The principal coordinates analysis (PCoA) also supported the results of Permanova. For high planting density conditions, plots on the right side at the top and the bottom of the PCoA were dominated by Plow and T26, while in the middle-left side the CLAAG plots were dominant. In the low planting density conditions, the convex hulls of the plots of the three treatments largely overlap each other (Fig. 4). Shrub and deciduous density had the strongest relationship with community patterns for bryophytes on PCoA 1, whereas vegetation cover, soil C/N ratio and conifer density appeared related with community patterns for bryophytes on PCoA 2 (Fig. 4). Ptilidium ciliare (L.) Hampe and Dicranum scoparium Hedw. were positively associated to PCoA 1, while Polytrichum commune Hedw. and a group of Sphagnum spp. were associated positively with shrub density to PCoA 1 (Fig. 4).
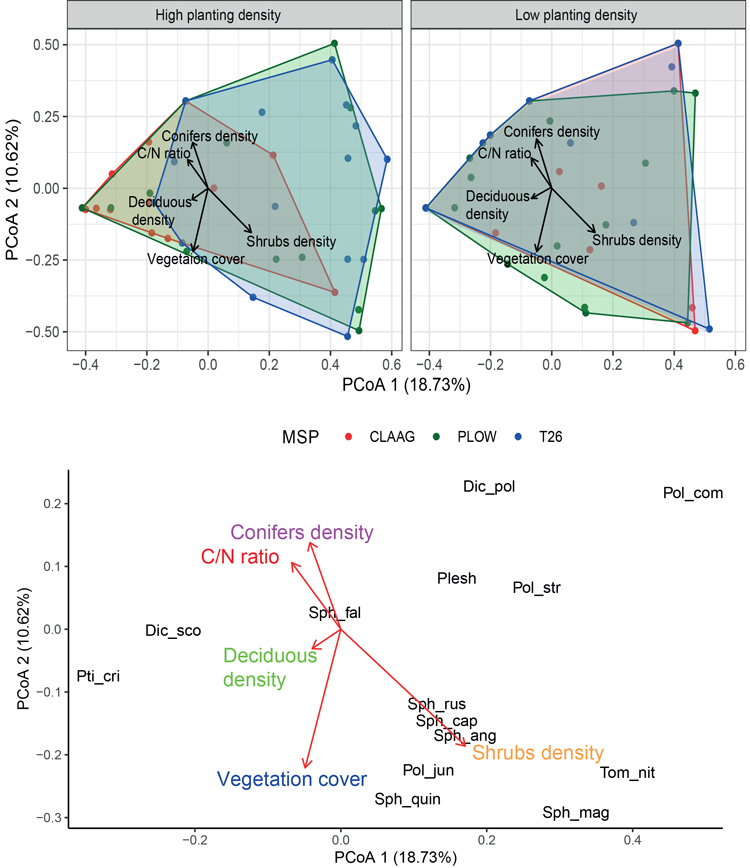
Fig. 4. Principal coordinates analysis (PCoA) for the bryophyte composition using the Bray-Curtis dissimilarity index and a Cailliez correction. Species names and environmental conditions were added by correlation to the axes. Colors in convex hull indicate different mechanical site preparation (MSP) treatments. See Suppl. file S1 for full species names.
3.3 Understory diversity
For vascular plant communities, there was no significant effect of planting density, MSP, and their interaction on the dominance and the diversity-based Simpson and Shannon indices, respectively (Table 1). In bryophyte communities, total cover was significantly influenced by planting density in interaction with MSP, with greater bryophyte cover in low planting density plots compared to high planting density plots in the plow treatment (Table 1).
3.4 Functional traits of the understory
One of the bryophytes and two of the vascular plant functional traits varied with planting density and MSP (Table 4). For bryophyte traits, water content at field capacity was significantly affected by MSP (Table 4), as it was higher in the plowing treatment compared to the other treatments (Fig. 5A). For vascular plant communities, the abundance of small shrubs was influenced by the interaction between planting density and MSP, while herbaceous plants were only affected by planting density (Table 4). Small shrub abundance was higher in Plow – low planting density plots than in other treatment combinations, while herbaceous abundance was higher in high planting density plots than in low planting density plots (Figs. 5B, C).
| Table 4. Results from linear mixed model for the main and interacting effects of planting density of black spruce (Picea mariana) and mechanical site preparation on functional trait of vascular plant and bryophytes. Df = degrees of freedom. Bold indicates significance at α ≤ 0.05. | ||||||
| Response variable | Treatment effect | |||||
| Planting density | Mechanical site preparation (MSP) | Planting density × MSP | ||||
| Df | p-value | Df | p-value | Df | p-value | |
| Functional trait of bryophytes | ||||||
| Water content at field capacity (%) | 1 | 0.77 | 2 | 0.02 | 2 | 0.5 |
| Colony density (mg cm–3) | 1 | 0.93 | 2 | 0.51 | 2 | 0.09 |
| Absorption capacity (ml cm–3) | 1 | 0.13 | 2 | 0.86 | 2 | 0.16 |
| Percolation (% cm –3) | 1 | 0.74 | 2 | 0.95 | 2 | 0.58 |
| Functional trait of vascular plants | ||||||
| Maximum height (cm) | 1 | 0.86 | 2 | 0.91 | 2 | 0.09 |
| Specific leaf area (cm2 g–1) | 1 | 0.93 | 2 | 0.43 | 2 | 0.14 |
| Small shrubs abundance | 1 | 0.55 | 2 | 0.05 | 2 | 0.055 |
| Herbaceous abundance | 1 | 0.01 | 2 | 0.7 | 2 | 0.84 |
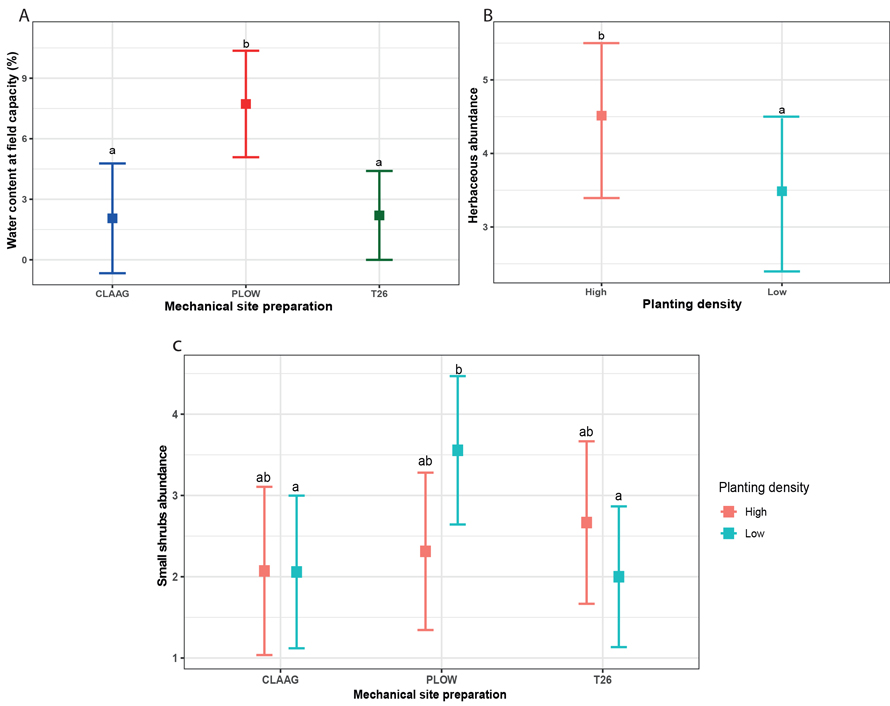
Fig. 5. Effects of planting density of black spruce (Picea mariana) (Low = 1100 seedlings ha–1; High = 2500 seedlings ha–1) and mechanical site preparation (CLAAG = careful logging around advance growth; Plow = plowing; T26 = disk trenching scarification) on water content at field capacity and the herbaceous abundance (A, B). Panel C illustrates the interacting effects of mechanical site preparation and planting density on the small shrubs abundance. Data are presented as estimated marginal means with 95% confidence intervals. Means followed by the same letter are not significatively different at α ≤ 0.05 based on linear mixed models with Tukey HSD pairwise comparisons. (Plots CLAAG low, n = 18; plots CLAAG high, n = 14; plots Plow low, n = 20; plots Plow high, n = 16; plots T26 low, n = 20; plots T26 high, n = 16). View larger in new window/tab.
3.5 Planted black spruce
Neither MSP, planting density or their interaction had a significant effect on planted black spruce height and ground-level diameter in 2020 (Table 5). Foliar N concentration was affected by MSP; it was higher in plots submitted to the plowing treatment than in the CLAAG plots, but not different than in the T26 plots (Table 5). Foliar C concentration and sapling growth between 2016 and 2020 were affected by the interacting effects of planting density and MSP (Table 5). Foliar C concentration was higher in low planting density conditions in CLAAG and plow treatment plots, and lower in T26 – low planting density plots (Fig. 6A). Sapling growth was higher in the high planting density, plow treatment plots, and lower in CLAAG and T26 – high planting density plots (Fig. 6B).
| Table 5. Results from linear mixed model for the main and interacting effects of planting density of black spruce (Picea mariana) and mechanical site preparation on height, growth, diameter at ground level, N and C foliar concentration of planted trees. Df = degrees of freedom. Bold indicates significance at α ≤ 0.05. | ||||||
| Response variable | Treatment effect | |||||
| Planting density | Mechanical site preparation (MSP) | Planting density × MSP | ||||
| Df | p-value | Df | p-value | Df | p-value | |
| Height trees measured in 2020 (cm) | 1 | 0.32 | 2 | 0.75 | 2 | 0.47 |
| Diameter at ground-level (mm) | 1 | 0.6 | 2 | 0.35 | 2 | 0.42 |
| Sapling growth measured between 2016–2020 (cm) | 1 | 0.89 | 2 | 0.88 | 1 | 0.03 |
| N foliar % | 1 | 0.88 | 2 | 0.02 | 2 | 0.29 |
| C foliar % | 1 | 0.2 | 2 | 0.001 | 2 | <0.001 |
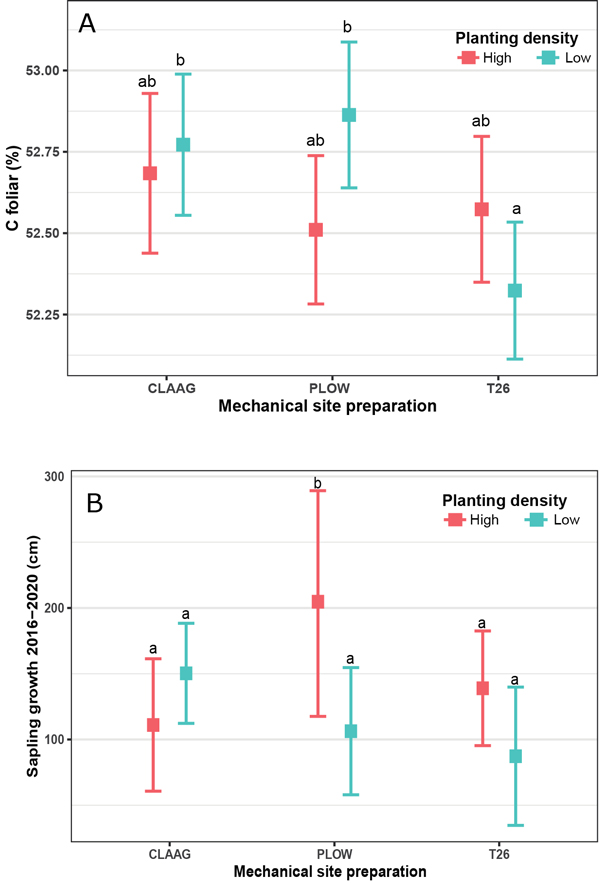
Fig. 6. Effects of planting density of black spruce (Picea mariana) (Low = 1100 seedlings ha–1; High = 2500 seedlings ha–1) and mechanical site preparation (CLAAG = careful logging around advanced growth; Plow = plowing; T26 = disk trenching scarification) on foliar C concentration (A) and on sapling growth (B). Data are presented as estimated marginal means with 95% confidence intervals. Means followed by the same letter are not significatively different at α ≤ 0.05 based on linear mixed models with Tukey HSD pairwise comparisons. (Plots CLAAG low, n = 32; plots CLAAG high, n = 25; plots Plow low, n = 30; plots Plow high, n = 29; plots T26 low, n = 34; plots T26 high, n = 30).
4 Discussion
As observed elsewhere (Akers et al. 2013; Henneb et al. 2019), planting density and MSP both affected environmental conditions and plant growth after nine years, though their effects seem limited in our study. We found that MSP had a greater effect than planting density on environmental conditions (Objective 1). Bryophyte cover was the only understory variable affected by planting density and MSP (Objective 2), most probably because of the higher sensitivity of bryophytes to environmental changes than other understory groups (Frego 2007). Responses to environmental changes depend on the species surrounding the bryophytes; by changing interspecific interactions, environmental changes modify the effects on bryophytes and further change species composition in communities (Bu et al. 2011). MSP also led to some dissimilarities in understory composition (Objective 2), as has been reported before (Löf et al. 2012). Furthermore, plant functional traits of vascular plants and bryophytes (abundance of small shrubs, herbaceous abundance, and water content at field capacity) were affected by planting density and MSP (Objective 3). Sapling growth depended on the interaction between planting density and MSP (Objective 4); high planting density in plots submitted to plowing resulted in greater growth than in the other treatments.
4.1 Effect of planting density and MSP on environmental conditions
We found a higher density of naturally established conifers after CLAAG than after the other treatments, which differs from the results of previous studies that reported greater conifer density after CLAAG + MSP. These studies were conducted decades after treatments, while our study was carried out after just nine years, which encompasses the normal black spruce establishment time after disturbance (Greene et al. 1999). Time after treatment and the time to canopy closure could explain this difference in our results, as a closed canopy normally occurs after more than 30 years in this portion of the boreal forest (Taylor et al. 1988; Kane et al. 2010).
We found no effect of planting density on regenerated conifer density, which was a surprising result. We posit that stands with low planting density provided more space and opportunities for spontaneous colonization by natural trees than high density planted stands, as there would be a greater availability of resources such as light, water and nutrients (Shaoxiong et al. 2012). In contrast, higher plantation density can lead to greater intraspecific competition (Boyden et al. 2008), which could induce a decrease in survival compared to lower planting density conditions (Akers et al. 2013). Therefore, based on our results only 9 years after planting, it is not clear which planting density is best for forest management (with an objective of producing wood fibre), as some advantages of both high and low planting densities were detectable. Furthermore, the interplay between planting density and forest productivity is dynamic and changes over time. While stand volume generally increases with higher planting density, this trend only persists up to a certain threshold (Baldwin Jr. et al. 2000). Beyond this point, denser planting may lead to reduced growth per tree and an increase in mortality (Smith et al. 1997). Conversely, lower planting densities typically promote the growth in diameter of individual trees, albeit with an increase in stem taper over time. This can have long-term effects on certain wood characteristics (Mvolo et al. 2022). Additionally, the influence of planting density on the natural regeneration of preferred species is an important factor to consider.
4.2 Effect on the understory composition and the functional traits
Understory composition was affected by the interaction between planting density and MSP, while these differences detected by Permanova were not visually obvious on the PCoA. The composition of vascular plant communities differed among MSP at both lower and higher planting densities. Only plots submitted to CLAAG and T26 did not present a difference in vascular plant composition at the low planting density. The composition of the bryophyte community differed among treatments at the higher planting density, with a clear difference between CLAAG vs the plow and T26 treatments. This difference might have been caused by the environmental conditions found under high planting density regime (lower light availability, higher nutrient availability and higher moisture), which would be affected by the higher deciduous sapling density in this treatment (Rodríguez-Calcerrada et al. 2008; Tinya et al. 2009). Few studies are available to explore potential reasons for these findings; further studies are needed to clarify the mechanisms behind this response.
Moreover, understory composition was affected by the soil C/N ratio, coniferous species, and deciduous species density. Deciduous species density was the main driver of many vascular plant community composition patterns. This is consistent with previous findings that show how deciduous stands tend to have an understory with higher species richness than coniferous stands, as they are characterized by higher understory light and soil nutrient availability (Légaré et al. ٢٠٠٢; Roberts 2004; Tinya et al. 2009). Furthermore, the topsoil of coniferous stands usually has higher C/N ratios, lower pH values and lower nutrient availability compared to that found in deciduous stands (Légaré et al. 2001; Augusto et al. 2003), which might be a limiting factor for vascular species establishment and growth (Barbier et al. 2008). In contrast to the vascular plants, the environmental conditions had little effect on the first axis of bryophyte composition. However, as observed for vascular plants, conifer density, C/N ratio, and shrub density drove the pattern on the second axis of bryophyte composition. The first axis may have been affected by other variables influencing the bryophyte community that we did not measure, such as the thickness of the organic layer or the position of the water table (Fenton and Bergeron 2006).
To our knowledge, our study is the first to evaluate the potential interacting effects of planting density and MSP on functional traits of vascular plants and bryophytes. Our findings for some vascular plant traits were similar to those reported by Wei et al. (2019), who found that MSP affected vascular plants traits after soil disturbance and as a function of light availability. Hence, MSP played an important role in understory functional composition and diversity. The positive effect of higher planting density in interaction with MSP on small shrubs and herbaceous species abundance suggests that their abundance could be responsive to environmental changes (Haeussler et al. 2002). The effects that we observed for small shrubs and herbaceous species support that trait–growth rate relationships are much stronger for small plants than for large trees. Interspecific variation in functional traits (demographic attributes) is indeed much larger for small plants than for large trees, which makes it statistically easier to detect environmental changes–functional trait relationships (Poorter et al. 2008).
Functional trait values have important consequences for ecosystem processes, properties, and services (Garnier et al. 2004; Fortunel et al. 2009; Conti and Díaz 2013). The relationship between vascular plants traits and sapling growth was not significant (results not shown), contrary to the results of Wei et al. (2021), who reported a significant relationship between vascular plant traits and sapling growth in CLAAG sites after 25 years. This indicates that time since treatment, age, and tree size play an important role on this trait–growth relationship. Further studies about functional traits and sapling growth are needed to further understand these results.
Among the traits we measured, only the water content of bryophytes was affected by MSP, being higher in the plowing treatment compared to the other treatments. Previous studies also support the role of water content as an indicator of environmental changes (Zotz et al. 2000; Rice and Schneider 2004); water content in bryophytes is controlled by forest canopy structural properties as it mediates the amount of light reaching the understory (Michel et al. 2013). Consequently, our results suggest that MSP could affect bryophyte water content through its effects on stand structure. MSP could ultimately affect ecological services provided by bryophytes, as leaf water content is an important limiting factor for bryophyte photosynthesis (Newmaster et al. 2007). The water dynamics of nonvascular plants are not as well understood or quantified as those of vascular plants and are rarely modelled (Sonnentag et al. 2008; Bond-Lamberty et al. 2011). Therefore, further studies are needed to better understand this relationship.
4.3 Black spruce growth
We posit that after the plowing treatment, higher planting density probably led to higher tree litter production and lower ericaceous shrub litter, which resulted in higher nutrient availability for sapling growth (Joanisse et al. 2007) than in the lower planting density (Guo and Sims 1999). Previous studies conducted on the same site demonstrated that plowing favoured early seedling growth, compared to the other treatments (Henneb et al. 2015; Henneb et al. 2019). The higher growth of black spruce in plowed plots (highest severity disturbance) can be explained by the resultant higher soil temperatures and lower interception of water by competing vegetation, which is known to increases soil water availability and favours root growth (Hébert et al. 2006). In support of this interpretation, we found that foliar nutrient concentrations, especially N, were higher in plots submitted to plowing compared to plots treated by CLAAG only or T26. Other studies have found positive effects of plowing, even in the long-term (Mäkitalo et al. 2010; Hjelm et al. 2019).
5 Conclusions
Our study has provided valuable insights into the interacting effects of planting density and MSP on plantation characteristics, yet it is important to acknowledge certain limitations in our sampling approach. The scale and scope of our sampling were primarily designed to capture short-term effects, which may not fully represent long-term ecological dynamics and responses. For instance, the absence of significant effects on tree diameter after nine years suggests that longer-term observations might be necessary to fully understand the impact of planting density on tree growth and forest productivity.
Additionally, while our study highlighted the utility of bryophyte cover as an indicator of treatment effects, the responses of other understory components, particularly vascular plants, were less pronounced. This indicates a need for more detailed investigation into the functional responses of different understory species, which could yield a more nuanced understanding of how silvicultural treatments influence forest ecosystems. Moreover, our study design encompassed only two initial planting densities – 1100 and 2500 seedlings ha–1. This limited range of densities might have restricted our capacity to thoroughly assess treatment effects on both the ecosystem and the growth responses of planted trees in their early developmental stage. The extent of competitive interactions among individuals and their impact on understory vegetation, therefore, may not have been fully established at these densities.
Future research could benefit from extended monitoring to capture long-term trends and responses. Incorporating a wider range of ecological indicators, such as soil microbial activity or wildlife habitat use, could also support our understanding of how plantation management practices impact forest ecosystems. Further studies might also explore the interaction of these silvicultural techniques with varying climatic conditions, particularly in the context of global change. Such investigations would not only increase our knowledge of forest management practices but also contribute to the development of strategies that enhance forest resilience and productivity in a changing global environment (Achim et al. 2022).
Acknowledgements
This manuscript was prepared from the Master’s thesis of the main author. We are grateful to Julie Arseneault, Fanny Dubois, Marie-Anyse Dubuc, Sameera Liyanage, and Xavier St-Amant for their help in field work, and to Daniel Lesieur and Ange-Marie Botroh for support in data management and statistical analyses. We thank Dr. David Paré and Sébastien Dagnault for their contribution to chemical analyses. We thank Dr. Kaysandra Waldron, Dr. Mélanie Jean, two anonymous reviewers and the editors for their constructive comments on earlier versions of the manuscript. This work aligns with the objectives of the IUFRO Task Force on Resilient Planted Forests Serving Society and Bioeconomy, of which Nelson Thiffault is a member.
Funding
This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC), in collaboration with Natural Resources Canada, GreenFirst Forest Products and the UQAT-UQAM Chair in Sustainable Forest Management.
Author contributions
Amira Fetouab: Data curation; Formal analysis; Investigation; Visualization; Writing - original draft. Nicole Fenton: Conceptualization; Funding acquisition; Methodology; Project administration; Resources; Supervision; Validation; Writing - review & editing. Nelson Thiffault: Conceptualization; Funding acquisition; Methodology; Supervision; Validation; Writing - review & editing. Martin Barrette: Funding acquisition; Writing - review & editing.
Data availability
Data will be made available on request.
References
Achim A, Moreau G, Coops N, Axelson J, Barrette J, Bédard S, Byrne K, Caspersen J, Dick A, D’Orangeville L, Drolet G, Eskelson B, Filipescu C, Flamand-Hubert M, Goodbody T, Griess V, Hagerman S, Keys K, Lafleur B, White J (2022) The changing culture of silviculture. Forestry 95: 143–152. https://doi.org/10.1093/forestry/cpab047.
Akers MK, Kane M, Zhao D, Teskey RO, Daniels RF (2013) Effects of planting density and cultural intensity on stand and crown attributes of mid-rotation loblolly pine plantations. For Ecol Manage 310: 468–475. https://doi.org/10.1016/j.foreco.2013.07.062.
Aubin I. Cardou F, Boisvert‐Marsh L, Garnier E, Strukelj M, Munson AD (2020) Managing data locally to answer questions globally: the role of collaborative science in ecology. J Veg Sci 31: 509–517. https://doi.org/10.1111/jvs.12864.
Augusto L, Dupouey JL, Ranger J (2003) Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann For Sci 60: 823–831. https://doi.org/10.1051/forest:2003077.
Baldwin Jr. VC, Peterson KD, Clark III A, Ferguson RB, Strub MR, Bower DR (2000) The effects of spacing and thinning on stand and tree characteristics of 38-year-old loblolly pine. For Ecol Manage 137: 91–102. https://doi.org/10.1016/S0378-1127(99)00340-0.
Barbier S, Gosselin F, Balandier P (2008) Influence of tree species on understory vegetation diversity and mechanisms involved – a critical review for temperate and boreal forests. For Ecol Manage 254: 1–15. https://doi.org/10.1016/j.foreco.2007.09.038.
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. J Stat Soft 67: 1–48. https://doi.org/10.18637/jss.v067.i01.
Benomar L, DesRochers A, Larocque GR (2012) The effects of spacing on growth, morphology and biomass production and allocation in two hybrid poplar clones growing in the boreal region of Canada. Trees 26: 939–949. https://doi.org/10.1007/s00468-011-0671-6.
Bock MD, Van Rees KC (2002) Mechanical site preparation impacts on soil properties and vegetation communities in the Northwest Territories. Can J For Res 32: 1381–1392. https://doi.org/10.1139/x02-067.
Bond-Lamberty B, Gower ST, Amiro B, Ewers BE (2011) Measurement and modelling of bryophyte evaporation in a boreal forest chronosequence. Ecohydrology 4: 26–35. https://doi.org/10.1002/eco.118.
Boyden S, Binkley D, Stape JL (2008) Competition among Eucalyptus trees depends on genetic variation and resource supply. Ecology 89: 2850–2859. https://doi.org/10.1890/07-1733.1.
Bremer LL, Farley KA (2010) Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodiv Conserv 19: 3893–3915. https://doi.org/10.1007/s10531-010-9936-4.
Bu ZJ, Rydin H, Chen S (2011) Direct and interaction-mediated effects of environmental changes on peatland bryophytes. Oecologia 166: 555–563. https://doi.org/10.1007/s00442-010-1880-1.
Conti G, Díaz S (2013) Plant functional diversity and carbon storage – an empirical test in semi-arid forest ecosystems. J Ecol 101: 18–28. https://doi.org/10.1111/1365-2745.12012.
Cornelissen JH, Lang SI, Soudzilovskaia NA, During HJ (2007) Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry. Ann Bot 99: 987–1001. https://doi.org/10.1093/aob/mcm030.
Environment Canada (2021) Canadian climate normals – Climate – Environment and climate change Canada. https://climate.weather.gc.ca/climate_normals/index_e.html.
Faubert J (2013) Flore des bryophytes du Québec-Labrador, Volume 2: Mousses, première partie. [Flora of the Bryophytes of Quebec-Labrador, Volume 2: Mosses, Part 1]. Société québécoise de la bryologie, Saint-Valérien, Québec.
Fenton NJ, Bergeron Y (2006) Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. J Veg Sci 17: 65–76. https://doi.org/10.1111/j.1654-1103.2006.tb02424.x.
Fenton NJ, Lecomte N, Légaré S, Bergeron Y (2005) Paludification in black spruce (Picea mariana) forests of eastern Canada: potential factors and management implications. For Ecol Manage 213: 151–159. https://doi.org/10.1016/j.foreco.2005.03.017.
Fortunel C, Garnier E, Joffre R, Kazakou E, Quested H, Grigulis K, Lavorel S, Ansquer P, Castro H, Cruz P, DoleŽal J, Eriksson O, Freitas, H, Golodets, C, Jouany, C, Kigel J, Kleyer M, Lehsten V, Lepš J, Meier T, Pakeman R, Papadimitriou M, Papanastasis VP, Quétier F, Robson M, Sternberg M, Theau JP, Thébault A, Zavorali M (2009) Leaf traits capture the effects of land use changes and climate on litter decomposability of grasslands across Europe. Ecology 90: 598–611. https://doi.org/10.1890/08-0418.1.
Frego KA (2007) Bryophytes as potential indicators of forest integrity. For Ecol Manage 242: 65–75. https://doi.org/10.1016/j.foreco.2007.01.030.
Garnier E, Cortez J, Billès G, Navas ML, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint JP (2004) Plant functional markers capture ecosystem properties during secondary succession. Ecology 85: 2630–2637. https://doi.org/10.1890/03-0799.
Gilliam FS (2007) The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57: 845–858. https://doi.org/10.1641/B571007.
Gilliam FS, Roberts MR (2003) Interactions between the herbaceous layer and overstory canopy of eastern forests. In: Gilliam F (ed) The herbaceous layer in forests of eastern North America. Oxford University Press, Oxford, pp 233–254. https://doi.org/10.1093/acprof:osobl/9780199837656.003.0009.
Greene DF, Zasada JC, Sirois L, Kneeshaw D, Morin H, Charron I, Simard MJ (1999) A review of the regeneration dynamics of North American boreal forest tree species. Can J For Res 29: 824–839. https://doi.org/10.1139/x98-112.
Guo LB, Sims REH (1999) Litter production and nutrient return in New Zealand eucalypt short-rotation forests: implications for land management. Agric Ecosyst Environ 73: 93–100. https://doi.org/10.1016/S0167-8809(99)00006-7.
Haeussler S, Bedford L, Leduc A, Bergeron Y, Kranabetter J (2002) Silvicultural disturbance severity and plant communities of the southern Canadian boreal forest. Silva Fenn 36: 307–327. https://doi.org/10.14214/sf.565.
Hébert F, Boucher JF, Bernier PY, Lord D (2006) Growth response and water relations of 3-year-old planted black spruce and jack pine seedlings in site prepared lichen woodlands. For Ecol Manage 223: 226–236. https://doi.org/10.1016/j.foreco.2005.11.005.
Hedwall PO, Skoglund J, Linder S (2015) Interactions with successional stage and nutrient status determines the life-form-specific effects of increased soil temperature on boreal forest floor vegetation. Ecol Evol 5: 948–960. https://doi.org/10.1002/ece3.1412.
Heiskanen J, Saksa T, Luoranen J (2013) Soil preparation method affects outplanting success of Norway spruce container seedlings on till soils susceptible to frost heave. Silva Fenn 47, article id 893. https://doi.org/10.14214/sf.893.
Henneb M, Valeria O, Fenton NJ, Thiffault N, Bergeron Y (2015) Mechanical site preparation: Key to microsite creation success on Clay Belt paludified sites. For Chron 91: 187–196. https://doi.org/10.5558/tfc2015-030.
Henneb M, Valeria O, Thiffault N, Fenton NJ, Bergeron Y (2019) Effects of mechanical site preparation on microsite availability and growth of planted black spruce in canadian paludified forests. Forests 10, article id 670. https://doi.org/10.3390/f10080670.
Hjelm K, Nilsson U, Johansson U, Nordin P (2019) Effects of mechanical site preparation and slash removal on long-term productivity of conifer plantations in Sweden. Can J For Res 49: 1311–1319. https://doi.org/10.1139/cjfr-2019-0081.
Joanisse GD, Bradley RL, Preston CM, Munson AD (2007) Soil enzyme inhibition by condensed litter tannins may drive ecosystem structure and processes: the case of Kalmia angustifolia. New Phytol 175: 535–546. https://doi.org/10.1111/j.1469-8137.2007.02113.x.
Johansson K, Nilsson U, Örlander G (2013) A comparison of long-term effects of scarification methods on the establishment of Norway spruce. Forestry 86: 91–98. https://doi.org/10.1093/forestry/cps062.
Johansson M (1994) The influence of soil scarification on the turn‐over rate of slash needles and nutrient release. Scan J For Res 9: 170–179. https://doi.org/10.1080/02827589409382828.
Kane VR, Bakker JD, McGaughey RJ, Lutz JA, Gersonde RF, Franklin JF (2010) Examining conifer canopy structural complexity across forest ages and elevations with LiDAR data. Can J For Res 40: 774–787. https://doi.org/10.1139/X10-064.
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Func Ecol 16: 545–556. https://doi.org/10.1046/j.1365-2435.2002.00664.x.
Légaré S, Bergeron Y, Leduc A, Paré D (2001) Comparison of the understory vegetation in boreal forest types of southwest Quebec. Can J Bot 79: 1019–1027. https://doi.org/10.1139/b01-076.
Légaré S, Bergeron Y, Paré D (2002) Influence of forest composition on understory cover in boreal mixedwood forests of western Quebec. Silva Fenn 36: 353–366. https://doi.org/10.14214/sf.567.
Lenth RV, Buerkner P, Herve M, Love J, Miguez F, Riebl H, Singmann H (2022) emmeans: estimated marginal means, aka least-squares means (version 1.7.2). https://CRAN.R-project.org/package=emmeans.
Löf M, Dey DC, Navarro RM, Jacobs DF (2012) Mechanical site preparation for forest restoration. New For 43: 825–848. https://doi.org/10.1007/s11056-012-9332-x.
Luoranen J, Viiri H, Sianoja M, Poteri M, Lappi J (2017) Predicting pine weevil risk: effects of site, planting spot and seedling level factors on weevil feeding and mortality of Norway spruce seedlings. For Ecol Manage 389: 260–271. https://doi.org/10.1016/j.foreco.2017.01.006.
Mäkitalo K, Alenius V, Heiskanen J, Mikkola K (2010) Effect of soil physical properties on the long-term performance of planted Scots pine in Finnish Lapland. Can J Soil Sci 90: 451–465. https://doi.org/10.4141/CJSS08070.
Mallik A, Kravchenko D (2016) Black spruce (Picea mariana) restoration in Kalmia heath by scarification and microsite mulching. For Ecol Manage 362: 10–19. https://doi.org/10.1016/j.foreco.2015.10.020.
Michel P, Payton IJ, Lee WG, During HJ (2013) Impact of disturbance on above-ground water storage capacity of bryophytes in New Zealand indigenous tussock grassland ecosystems. New Z J Ecol 37: 114–126.
Moroni MT, Thiffault N, Titus BD, Mante C, Makeschin F (2009) Controlling Kalmia and reestablishing conifer dominance enhances soil fertility indicators in central Newfoundland, Canada. Can J For Res 39: 1270–1279. https://doi.org/10.1139/X09-061.
Mvolo CS, Goudiaby V, Koubaa A, Stewart JD (2022) Influence of four spacings between trees and four samplings heights on selected wood quality attributes of white spruce (Picea glauca (Moench) Voss). Forests 13, article id 1807. https://doi.org/10.3390/f13111807.
Newmaster SG, Parker WC, Bell FW, Paterson JM (2007) Effects of forest floor disturbances by mechanical site preparation on floristic diversity in a central Ontario clearcut. For Ecol Manage 246: 196–207. https://doi.org/10.1016/j.foreco.2007.03.058.
Nilsson MC, Wardle DA (2005) Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front Ecol Environ 3: 421–428. https://doi.org/10.1890/1540-9295(2005)003[0421:UVAAFE]2.0.CO;2.
Nordborg F, Nilsson U, Örlander G (2003) Effects of different soil treatments on growth and net nitrogen uptake of newly planted Picea abies (L.) Karst. seedlings. For Ecol Manage 180: 571–582. https://doi.org/10.1016/S0378-1127(02)00650-3.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, Mc Glinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens HH, Szoecs E, Wagner H (2020) vegan: community ecology package (version 2.5-7). https://CRAN.R-project.org/package=vegan.
Paquette A, Messier C (2010) The role of plantations in managing the world’s forests in the Anthropocene. Front Ecol Environ 8: 27–34. https://doi.org/10.1890/080116.
Paradis E, Schliep K (2019) ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35: 526–528. https://doi.org/10.1093/bioinformatics/bty633.
Poorter L, Wright SJ, Paz H, Ackerly DD, Condit R, Ibarra-Manríquez G, Harms KE, Licona JC, Martínez-Ramos M, Mazer SJ, Muller-Landau HC, Peña-Claros M, Webb CO, Wright IJ (2008) Are functional traits good predictors of demographic rates? Evidence from five neotropical forests. Ecology 89: 1908–1920. https://doi.org/10.1890/07-0207.1.
Powers RF (2006) Long-Term Soil Productivity: genesis of the concept and principles behind the program. Can J For Res 36: 519–528. https://doi.org/10.1139/x05-279.
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. https://www.r-project.org/.
Ramovs BV, Roberts MR (2003) Understory vegetation and environment responses to tillage, forest harvesting, and conifer plantation development. Ecol Applic 13: 1682–1700. https://doi.org/10.1890/02-5237.
Reicis K, Bradley RL, Joanisse G, Thiffault N, Scott D, Parsons WFJ (2023) Mechanical scarification can reduce competitive traits of boreal ericaceous shrubs and improve nutritional site quality. Forestry 96: 293–303. https://doi.org/10.1093/forestry/cpac047.
Rice SK, Schneider N (2004) Cushion size, surface roughness, and the control of water balance and carbon flux in the cushion moss Leucobryum glaucum (Leucobryaceae). Am J Bot 91: 1164–1172. https://doi.org/10.3732/ajb.91.8.1164.
Roberts MR (2004) Response of the herbaceous layer to natural disturbance in North American forests. Can J Bot 82: 1273–1283. https://doi.org/10.1139/b04-091.
Rodríguez-Calcerrada J, Mutke S, Alonso J, Gil L, Pardos JA, Aranda I (2008) Influence of overstory density on understory light, soil moisture, and survival of two underplanted oak species in a Mediterranean montane Scots pine forest. For Syst 17: 31–38. https://doi.org/10.5424/srf/2008171-01021.
Saetre P, Saetre LS, Brandtberg PO, Lundkvist H, Bengtsson J (1997) Ground vegetation composition and heterogeneity in pure Norway spruce and mixed Norway spruce – birch stands. Can J For Res 27: 2034–2042. https://doi.org/10.1139/x97-177.
Saucier JP, Robitaille A, Grondin P (2009) Cadre bioclimatique du Québec. [Quebec’s Bioclimatic Framework]. In: Doucet R, Côté M (eds) Manuel de foresterie. [Forestry Manual], 2nd ed. Ordre des ingénieurs forestiers du Québec, Éditions Multimondes, Québec, pp 186–205.
Shaoxiong C, Minsheng Y, Liping W (2012) Effect of spacing on volume, storm-resistance, and wood quality of Eucalyptus urophylla. For Res 11: 435–438.
Smith DM, Larson BC, Kelty MJ, Ashton PMS (1997) The practice of silviculture. Applied Forest Ecology, 9th ed. John Wiley & Sons, Inc.
Sonnentag O, Chen JM, Roulet NT, Ju W, Govind A (2008) Spatially explicit simulation of peatland hydrology and carbon dioxide exchange: influence of mesoscale topography. J Geophys Res Biogeosci 113, article id G02005. https://doi.org/10.1029/2007JG000605.
Sutherland BJ, Foreman FF (1995) Guide to the use of mechanical site preparation equipment in northwestern Ontario. Great Lakes Forestry Centre, Canadian Forest Service, Natural Resources Canada, Sault Ste. Marie.
Sutherland BJ, Foreman FF (2000) Black spruce and vegetation response to chemical and mechanical site preparation on a boreal mixedwood site. Can J For Res 30: 1561–1570. https://doi.org/10.1139/x00-087.
Taylor SJ, Carleton TJ, Adams R (1988) Understory vegetation change in a Picea mariana chronosequence. J Veg Sci 73: 63–72. https://doi.org/10.1007/BF00031853.
Thiffault N, Jobidon R (2006) How to shift unproductive Kalmia angustifolia – Rhododendron groenlandicum heath to productive conifer plantation. Can J of For Res 36: 2364–2376. https://doi.org/10.1139/x06-090.
Thiffault N, Lenz PRN, Hjelm K (2023) Plantation Forestry, Tree Breeding, and Novel Tools to Support the Sustainable Management of Boreal Forests. In: Montoro Girona M, Morin H, Gauthier S, Bergeron Y (eds) Boreal forests in the face of climate change. Sustainable management. Advances in Global Change Research 74, Springer, Cham, pp 383–401. https://doi.org/10.1007/978-3-031-15988-6_14.
Tinya F, Márialigeti S, Király I, Németh B, Ódor P (2009) The effect of light conditions on herbs, bryophytes, and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant Ecol 204: 69–81. https://doi.org/10.1007/s11258-008-9566-z.
Vincent J, Hardy L (2011) L’évolution et l’extension des lacs glaciaires Barlow et Ojibway en territoire québécois. [The evolution and extension of glacial lakes Barlow and Ojibway in Quebec]. Géogr Phys Quatern 3–4: 357–372. https://doi.org/10.7202/1000283AR.
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007) Let the concept of trait be functional! Oikos 116: 882–892. https://doi.org/10.1111/j.2007.0030-1299.15559.x.
Wallertz K, Björklund N, Hjelm K, Petersson M, Sundblad LG (2018) Comparison of different site preparation techniques: quality of planting spots, seedling growth and pine weevil damage. New For 49: 705–722. https://doi.org/10.1007/s11056-018-9634-8.
Wei L, Fenton NJ, Lafleur B, Bergeron Y (2019) The combined role of retention pattern and post-harvest site preparation in regulating plant functional diversity: a case study in boreal forest ecosystems. Forests 10, article id 1006. https://doi.org/10.3390/f10111006.
Wei L, Thiffault N, Barrette M, Fenton NJ, Bergeron Y (2021) Can understory functional traits predict post-harvest forest productivity in boreal ecosystems? For Ecol Manage 495, article id 119375. https://doi.org/10.1016/j.foreco.2021.119375.
Zhou X, Zhu H, Wen Y, Goodale UM, Li X, You Y, Ye D, Liang H (2018) Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. For Ecol Manage 410: 164–173. https://doi.org/10.1016/j.foreco.2017.11.015.
Zotz G, Schweikert A, Jetz W, Westerman H (2000) Water relations and carbon gain are closely related to cushion size in the moss Grimmia pulvinata: research cushion size and moss ecophysiology. New Phytol 148: 59–67. https://doi.org/10.1046/j.1469-8137.2000.00745.x.
Total of 80 references.
