Simon Lebel Desrosiers,
Nicolas Bélanger,
Evelyne Thiffault,
Nelson Thiffault 
Climate change and transformation in forest fire regimes: an opportunity for the implementation of assisted migration of tree species in the Canadian boreal forest?
Lebel Desrosiers S., Bélanger N., Thiffault E., Thiffault N. (2025). Climate change and transformation in forest fire regimes: an opportunity for the implementation of assisted migration of tree species in the Canadian boreal forest? Silva Fennica vol. 59 no. 2 article id 25031. https://doi.org/10.14214/sf.25031
Highlights
- Increasing fire activity is reshaping post-disturbance landscapes in boreal forests
- Post-fire sites offer new opportunities for introducing climate-resilient tree species
- Fire can improve or impair site conditions for forest regeneration
- Assisted migration of tree species may enhance reforestation success after severe wildfires
- Research on post-fire regeneration of introduced species remains critically limited.
Abstract
Climate change is intensifying fire regimes in boreal forests, leading to ecological disruption and raising concerns about forest resilience and post-disturbance recovery. Altered fire dynamics creates novel opportunities for implementing adaptive silviculture for climate change, including assisted migration, the intentional movement and establishment of tree species or tree populations outside their current range of distribution to better match anticipated future climates. Here, we examine how the increasing frequency, severity, and spatial extent of Canadian boreal wildfires can serve as strategic windows for introducing climate-resilient tree species and genotypes. We review how fire influences the availability and suitability of post-fire sites for assisted migration, highlighting how fire-induced changes in soil abiotic and biotic properties may facilitate or hinder the establishment of relocated tree species. While fire can simplify site preparation, reduce biotic competition, and temporarily enhance soil nutrient availability, it may also degrade soil structure by consuming or altering soil organic matter and increasing soil susceptibility to erosion and disrupt essential mycorrhizal associations. We argue that assisted migration of tree species can be a proactive silvicultural tool when used in areas with regeneration failure or where future climate conditions are likely to exceed the tolerance limits of native species. Whilst scientific evidence remains limited on the regeneration success of migrated species and genotypes in post-fire environments, we argue for an integrated adaptation strategy that combines natural regeneration with targeted assisted migration interventions, guided by local site conditions, genetic considerations, and policy support, to build resilient boreal forests under changing disturbance regimes.
Keywords
fire severity;
site suitability;
post-fire regeneration;
climate change adaptation;
forest resilience;
reforestation strategies;
tree migration
-
Lebel Desrosiers,
Laboratoire sur la science des données, Université du Québec (TÉLUQ), 5800, rue Saint-Denis, bureau 1105, Montréal, Québec H2S 3L5, Canada
 https://orcid.org/0009-0007-1592-8505
E-mail
simon.lebeldesrosiers@teluq.ca
https://orcid.org/0009-0007-1592-8505
E-mail
simon.lebeldesrosiers@teluq.ca
- Bélanger, Laboratoire sur la science des données, Université du Québec (TÉLUQ), 5800, rue Saint-Denis, bureau 1105, Montréal, Québec H2S 3L5, Canada E-mail nicolas.belanger@teluq.ca
-
Thiffault,
Centre de recherche sur les matériaux renouvelables, Université Laval, 2425 De la Terrasse St, Québec, Québec G1V 0A6, Canada
https://orcid.org/0000-0001-9586-3834
E-mail
evelyne.thiffault@sbf.ulaval.ca
-
Thiffault,
Service canadien des forêts, Ressources naturelles Canada, 1055, rue Du P.E.P.S., C.P. 10380, Québec, Québec G1V 4C7, Canada
https://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@canada.ca
Received 26 June 2025 Accepted 21 August 2025 Published 1 September 2025
Views 33344
Available at https://doi.org/10.14214/sf.25031 | Download PDF

1 Introduction
Global warming is now a reality, bringing significant social, economic, and ecological disruptions. Despite efforts to limit warming to 1.5 °C, some tipping points may still be reached, as emissions from hard-to-mitigate sectors will persist (Abrams et al. 2023). Evidence of climate change on forest fire regimes is increasing worldwide. In Canada, ca. 17.2 million hectares burned in 2023 (CIFFC 2023), followed by 5.4 million hectares in 2024, far above the average of 2.5 million hectares/year (NRCan 2024). Mega-fires >100 000 hectares are more common, with over 29 such events recorded in Canada in 2023. The record-breaking area burned in 2023 resulted from only ~7100 fires, whereas ~10 000 fires were responsible for burning just half as much area in 1989 (Macdonald and Gauthier 2025).
Under climate change, boreal forests are expected to lose more area due to fire compared to temperate forests (Boulanger and Puigdevall 2021). This is partially due to the Arctic amplification effect, where higher latitudes undergo a greater temperature rise than the global average (Previdi et al. 2021). Fire activity is likely to increase in western Canada, whereas eastern regions may see more stable fire occurrences due to changes in the hydrological cycle (Coogan et al. 2021).
Fires play a key role in boreal forest regeneration by enabling tree seed dispersion from serotinous/semi-serotinous cones (Lamont et al. 2020), replenishing soil nutrients, favoring seed germination, reducing competition and preparing a favorable seedbed. However, while fire occurs naturally in many ecosystems, the relationship between climate change and fire regimes introduces uncertainties for forest health and resilience (Gauthier et al. 2015). Once considered stable carbon sinks, boreal forests are now switching to net carbon sources, including the Canadian boreal, mainly due to altered fire dynamics (Zhao et al. 2021).
Assisted migration (AM) involves the human-facilitated movement of species or genotypes from one location (or habitat) to another as a means to cope with changing environmental conditions such as those brought on by climate change (Aitken and Whitlock 2013). In the context of adaptive silviculture for climate change, this approach aims to aid tree species unable to shift at a rate sufficient to track shifting climatic niches. Trees, being stationary organisms reliant on seed dispersal to shift, are more vulnerable at their distribution range peripheries, where more extreme environmental conditions limit their survival (Padullés Cubino et al. 2024). New dynamics of forest disturbances under climate change will exacerbate these extremes, thus leading to new ecosystem trajectories in boreal ecosystems (e.g., favoring of temperate deciduous tree species in its southern regions) (Nayomi et al. 2022).
Assisted migration of tree species is increasingly framed as a strategy to maintain ecosystem services in boreal forests and elsewhere while supporting conservation and forestry objectives (Thiffault et al. 2024). In this paper, we argue that increased fire activity in Canadian boreal ecosystems presents a challenge for resilience, but also a critical opportunity to accelerate the implementation of AM of tree species, as part of adaptive silviculture for climate change. We explore how fire: (1) reshapes site availability and suitability, and (2) creates an opportunity for aligning post-fire reforestation with AM strategies. These complex interactions are synthesized in Fig. 1, which illustrates the intersecting roles of climate change, wildfire regimes, and boreal dynamics in shaping opportunities for AM of tree species (Fig. 1).
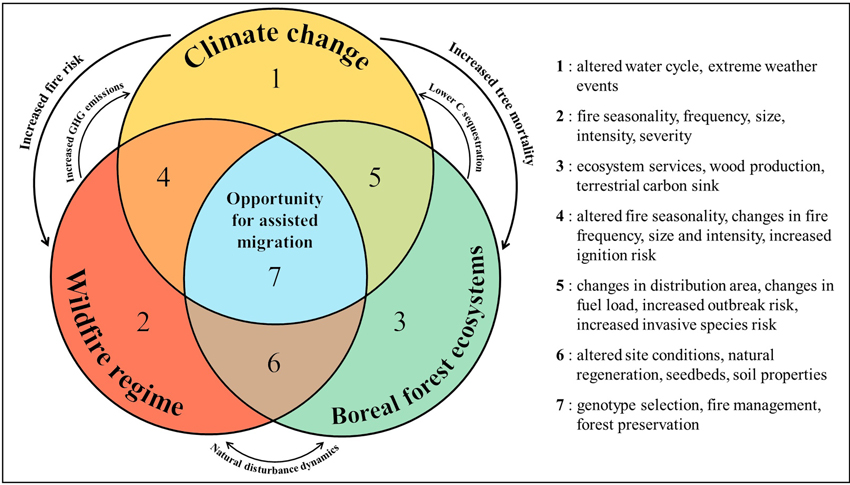
Fig. 1. Conceptual framework illustrating the interacting roles of climate change, wildfire regimes, and boreal forest ecosystems in shaping opportunities for assisted migration. Each circle represents a major driver of forest change, with overlapping zones identifying key consequences (zones 1 to 6) and the central intersection (zone 7) representing the opportunity space for implementing assisted migration strategies. Arrows indicate reinforcing feedbacks between drivers, which contribute to altered disturbance dynamics and ecosystem responses.
2 Fire and site availability for assisted migration of tree species
A significant increase in the number and size of wildfires worldwide, including the Canadian boreal forest, is anticipated under climate change (Grillakis et al. 2022). In addition to the recent mega-fires, fire disturbances could become 10 times more frequent by the end of the century under a medium-high GHG emissions scenario (Jones et al. 2024). Rising temperatures, prolonged droughts and extended warm seasons contribute to tree mortality, increasing the amount of available fuel (Matusick et al. 2018), although frequent fires may also reduce fuel loads and thus lead to less frequent and intense fires over time (Janssen and Veraverbeke 2025). High-intensity fires (releasing much energy as heat) have greater severity, which means more extensive tree mortality (Rossi et al. 2020) and possibly reduced organic-layer depth (Greene et al. 2007). Whereas prolonged droughts enhance fire intensity, cooler/wetter conditions mitigate it (Dong et al. 2021). Human activity also influences fire regimes through ignition, suppression, and prevention measures (e.g., prescribed burning) (Hunter and Robles 2020). As a whole, the annual area burned is projected to rise, although regional variations are expected due to changes in weather, fuel availability and even topography (Finney et al. 2021).
The increasing extent of burned landscapes is expected to create new opportunities for AM efforts. With increasing fire frequency and severity, regeneration failures of common tree species are anticipated to rise (Boucher et al. 2020). Forest regeneration, whether natural or through human intervention, is increasingly at risk. Thus, large-scale reforestation efforts (i.e., through planting or seeding) may be required to maintain forest landscapes and meet management objectives (Thiffault et al. 2025). Hence, land suitable for AM of tree species will not only become more abundant, but will also be available across larger spatial scales. Mega-fires are expected to create extensive areas where large-scale AM projects can be deployed, thus aligning with forest management planning, which is done at large spatial scales and over long periods (Zou et al. 2024). In Canada, forest management typically occurs on public lands under long-term, spatially explicit forest management plans that integrate silvicultural treatments, harvest scheduling, and regeneration objectives. These plans often cover decades and emphasize both timber production and ecological sustainability, providing a framework that can accommodate AM strategies at operational scales (Wotherspoon et al. In press).
Fire amplification effects will be more important in the higher-latitude boreal and temperate forests (Cunningham et al. 2024). Fire intensity and prevalence will more largely increase in these forests, meaning that long-distance AM may be necessary due to rapid shifts in climatic niches. Lower-latitude forests, however, may require strategies that focus more on altitudinal AM. Several AM approaches are possible for tree species, i.e., population migration, range expansion and species migration which are respectively movements (1) among seed zones within existing range limits, (2) at or just beyond existing range limits, and (3) well beyond existing range limits. Regardless of the specific approach, increasing fire activity will expand options for the deployment of AM programs for tree species. Large post-fire openings will create more suitable sites for establishing, through planting and sowing, tree species and genotypes better adapted to future climatic conditions, particularly in areas where natural regeneration is uncertain. Thus, AM of tree species is one potential response to the upcoming challenges that wildfires pose to maintaining productive forests.
3 Fire and site suitability for assisted migration of tree species
The impact of fire on soil varies depending on soil type and fire intensity and severity (thus affecting combustion completeness), as well as past disturbance history, fuel type, topography and post-fire precipitation patterns (Agbeshie et al. 2022). Low-intensity fires promote ash deposition, which contains nutrients such as Ca²⁺, Mg²⁺ and K⁺, thereby increasing their availability for plants, including tree seedlings (Zhang and Biswas 2017). The release of base cations also increases soil pH, as they displace acidity (e.g., H+, Al3+) from exchange sites. This alkalinity increases cation exchange capacity (CEC) and P and N availability (by affecting microbial activity, which is influenced by pH) (Neina 2019). Charcoal and organic matter addition after fire can also increase CEC, and soil microbial activity can be triggered by a heat-induced change in the lability of carbon inputs, as well as a change in the soil C:N ratio (Eckdahl et al. 2023). However, alkalinization and nutrient enrichment are transient and mainly localized to surface soils (Reid and Watmough 2014).
Fire can also lead to the oxidation and volatilization of growth-limiting nutrients in the boreal forest, such as N and P, and the destruction of soil organic matter (Agbeshie et al. 2022). This decreases soil productivity and structure over time. High-intensity fires can also cause mortality of soil microorganisms, particularly fungi, thus affecting nutrient cycling (Nelson et al. 2022). Bacteria can recover quickly, but fungal recovery takes longer and may be incomplete (Prendergast-Miller et al. 2017). This shift in microbial composition can temporarily enhance N availability, but there are long-term implications for ecosystem stability (Wang et al. 2012). Severe fires can also potentially degrade soil structure by destroying organic matter and thus lead to erosion (Mataix-Solera et al. 2011). The formation of hydrophobic layers during fire can increase surface runoff and erosion in some forests, but this is less common in boreal landscapes due to organic-rich and moist soils as well as moderate fire temperatures. In severely burned sites, changes include loss/alteration of soil organic matter, and reduction of essential nutrients, which may impair seedling establishment and long-term forest recovery. The transformation of soil organic matter into pyrogenic compounds may also reduce microbial activity and affect nutrient cycling (Jiménez-Morillo et al. 2020).
The extent to which these fire-induced changes in soil properties aid or harm post-fire regeneration of tree species potential depends on fire characteristics and the local soil and environment. Fire can also possibly facilitate the establishment of tree species and genotypes, not just by improving soil quality, but also by removing competing vegetation and thus, creating pathways that are favourable for the next tree cohort (Dawe et al. 2022). A key challenge in AM will be to select appropriate tree species and genotypes according to climate and soils (MacKenzie and Mahony 2021). The success of relocated tree species depends on matching their ecological requirements with the conditions of the host environment (Williams and Dumroese 2013).
Assisted migration may offer an opportunity to introduce such climate-resilient tree populations to sites where natural regeneration is likely to fail. Species such as eastern white pine (Pinus strobus L.), northern red oak (Quercus rubra L.), red pine (Pinus resinosa Sol. ex Aiton), and white oak (Quercus alba L.), native to more southern forest regions in Canada, may be suitable candidates for AM into boreal landscapes affected by high-severity fires, due to their tolerance to drought, nutrient-poor soils, and fire disturbance (Swanston et al. 2016). Other tested species for AM that have shown positive results include whitebark pine (Pinus albicaulis Engelm.), southern genotypes of trembling aspen (Populus tremuloides Michx.) and white spruce (Picea glauca (Moench) Voss) (e.g., Casmey et al. 2022). High-severity fires could even create potentially favourable conditions for nutrient-sensitive species such as sugar maple (Acer saccharum Marshall), yellow birch (Betula alleghaniensis Britton), or American beech (Fagus grandifolia Ehrh.) (Collin et al. 2017). However, such efforts must be informed by site-specific constraints and the ecological traits of potential candidate species. Some negative outcomes may hinder tree regeneration, particularly where mycorrhizal associations are essential for nutrient uptake and thus, seedling establishment (Bennett et al. 2017). In addition to selecting tree species and genotypes adapted to emerging post-fire conditions, using seedlings with mycorrhizae-inoculated roots is thus one approach that could enhance regeneration success. Silvicultural trials are beginning to explore how different management systems interact with forest AM (Thiffault et al. 2024), but long-term data on regeneration pathways and successes are still needed to guide effective deployment strategies (Thiffault et al. 2025). This could enhance forest stability and ecosystem function by ensuring that regenerating stands are better adapted to future climate stressors, including warmer temperatures, prolonged droughts, and changing soil microbial communities and nutrient availability.
A critical operational challenge for AM at the scale required after high-severity fires is the availability of appropriate seed sources. Large fires can affect tens of thousands of hectares, requiring rapid access to genetically suitable and climatically adapted reproductive material. In Canada, seed for reforestation is typically sourced either from seed orchards or collected from natural stands within established seed zones (Pedlar et al. 2012). However, current seed production capacity may not be sufficient to support large-scale implementation of AM, particularly for tree species outside their historical range. Also, planning of seed collection and storage can be complicated by species that exhibit specific mast-year seed production cycles. Strategic investments in seed supply infrastructure, including new orchard establishment, improved access to wild seed sources, and forward-looking seed planning, are essential research and policy priorities to support climate-adaptive reforestation strategies (Wotherspoon et al. In press).
Since most wildfires occur independently of human intervention, they eliminate, at least in part, the ecological or social concerns associated with removing native species to create space for AM efforts for trees, especially if natural regeneration is compromised. However, for other sites where natural regeneration is expected to be successful, a decision must be made between allowing regeneration of local species or introducing new ones that will be better suited to future climatic conditions. Other biotic interactions that play a role in determining whether relocated species can integrate successfully without disrupting the existing ecosystem must also be considered, such as potential predation (animals and insects) and diseases (Kracke et al. 2021).
4 Final thoughts on the possibility of a coherent adaptation strategy
Recent large-scale fires in the boreal forest raise a key question: can post-fire landscapes be leveraged to accelerate the implementation of AM strategies for tree species? The answer is not straightforward. As illustrated in Fig. 1, the convergence of altered fire dynamics, environmental stress, and climate pressures creates a multi-dimensional opportunity space for implementing AM of tree species. However, while fire can simplify site preparation and improve soil quality in the short term, it also introduces variability and unpredictability in post-disturbance conditions. Fire thus acts as both a catalyst and a constraint for AM deployment.
Fire creates a temporal and spatial window to introduce new species or genotypes without displacing established native vegetation, hence mitigating ethical concerns often associated with AM (Kracke et al. 2021). However, where natural regeneration of tree species is expected to succeed, a decision must still be made between allowing natural recovery and actively favouring tree species better suited to future climatic conditions. At the same time, AM strategies for tree species must navigate climate-related constraints, such as mismatches in phenology, spring frost risk, and photoperiod sensitivity (Ren et al. 2020). Mixed results from AM experiments for tree species such as sugar maple or oak reflect this complexity in the Canadian boreal forest (Kellman 2004). Provenance-based approaches, such as the migration of southern seed sources of white spruce or hybrid spruce (P. glauca × P. engelmannii), have demonstrated success in improving survival and growth under changing conditions (e.g., Casmey et al. 2022).
Assisted migration of tree species offers opportunities to match planting material to altered soil conditions, either by selecting genotypes pre-inoculated with compatible mycorrhizae or by favouring species with greater adaptability to soil disturbance (Bennett et al. 2017). Moreover, fire-driven mortality of overstory trees can reduce biotic competition and facilitate colonization by planted species, a phenomenon observed in initial succession following wildfire (Dawe et al. 2022). Despite this, regeneration pathways using AM tree species and genotypes under post-fire conditions remains largely undocumented.
Ultimately, fire reshapes both the urgency and the opportunity space for AM in the boreal forest. Leveraging this disturbance requires integration with silvicultural decision-making and forest policy. A hybrid strategy may be needed, supporting natural regeneration where viable, while introducing AM material in areas of high severity or uncertain recovery. Doing so aligns AM with climate-informed reforestation efforts (Cyr et al. 2022), while contributing to the long-term resilience of boreal forests in an era of increasing fire disturbance.
As fire activity intensifies in boreal regions, we thus argue that post-fire landscapes are emerging as strategic entry points for AM of tree species. These disturbances offer not only ecological conditions favourable to establishment but also a social and operational rationale for accelerating intervention. Fig. 1 provides a conceptual foundation for guiding such integrated responses to climate and disturbance-driven forest change. Assisted migration of tree species, when deployed thoughtfully alongside natural regeneration, can help align reforestation with future climatic realities. However, because empirical evidence remains scarce on regeneration outcomes of climate-adapted species and genotypes in post-fire environments, research and policy must converge to evaluate these pathways at the operational scale (Wotherspoon et al. In press). Doing so would position AM of tree species not as a last resort, but as an integrated, proactive tool for climate-adaptive forest management.
Authors contributions
SLD: Writing-original draft; NT: Funding acquisition, Text review and commentary; ET: Funding acquisition, Text review and commentary; NB: Funding acquisition, Project administration, Text review and commentary
Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the authors used DeepL SE’s DeepL and OpenAI’s ChatGPT in order to respectively translate from French to English and to improve the quality of language and expression. After using these tools/services, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Acknowledgements
We thank Dr. David Paré, Dr. Dan Kneeshaw and an anonymous reviewer for their constructive advice on an earlier version of this work.
Funding
This work was supported by a grant (reference ALLRP 572865-22) from the Natural Sciences and Engineering Research Council of Canada (PI: Nicolas Bélanger), obtained in collaboration with Domtar Inc. and Natural Resources Canada. It also benefited from a grant (reference 2020-RI-278290) from the Fonds de recherche du Québec – Nature et Technologies (PI: Evelyne Thiffault).
References
Abrams JF, Huntingford C, Williamson MS, Armstrong McKay DI, Boulton CA, Buxton JE, Sakschewski B, Loriani S, Zimm C, Winkelmann R, Lenton, TM (2023) Committed global warming risks triggering multiple climate tipping points. Earths Future 11, article id 2022EF003250. https://doi.org/10.1029/2022EF003250.
Agbeshie AA, Abugre S, Atta-Darkwa T, Awuah R (2022) A review of the effects of forest fire on soil properties. J Forestry Res 33: 1419–1441. https://doi.org/10.1007/s11676-022-01475-4.
Aitken SN, Whitlock MC (2013) Assisted gene flow to facilitate local adaptation to climate change. Ann Rev Ecol Evol Syst 44: 367–388. https://doi.org/10.1146/annurev-ecolsys-110512-135747.
Bennett JA, Maherali H, Reinhart KO, Lekberg Y, Hart MM, Klironomos J (2017) Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 355: 181–184. https://doi.org/10.1126/science.aai8212.
Boucher D, Gauthier S, Thiffault N, Marchand W, Girardin M, Urli M (2020) How climate change might affect tree regeneration following fire at northern latitudes: a review. New For 51: 543–571. https://doi.org/10.1007/s11056-019-09745-6.
Boulanger Y, Puigdevall JP (2021) Boreal forests will be more severely affected by projected anthropogenic climate forcing than mixedwood and northern hardwood forests in eastern Canada. Landsc Ecol 36: 1725–1724. https://doi.org/10.1007/s10980-021-01241-7.
Casmey M, Hamann A, Hacke UG (2022) Adaptation of white spruce to climatic risk environments in spring: implications for assisted migration. For Ecol Manage 525, article id 120555. https://doi.org/10.1016/j.foreco.2022.120555.
CIFFC (Canadian Interagency Forest Fire Centre) (2023) Canada report: 2023 fire season. https://ciffc.ca/sites/default/files/2024-03/03.07.24_CIFFC_2023CanadaReport%20%281%29.pdf. Accessed 3 May 2025.
Collin A, Messier C, Bélanger N (2017) Conifer presence may negatively affect sugar maple’s ability to migrate into the boreal forest through reduced foliar nutritional status. Ecosystems 20: 701–716. https://doi.org/10.1007/s10021-016-0045-4.
Coogan SC, Daniels LD, Boychuk D, Burton PJ, Flannigan MD, Gauthier S, Kafka V, Park JS, Wotton BM (2021) Fifty years of wildland fire science in Canada. Can J For Res 51: 283–302. https://doi.org/10.1139/cjfr-2020-0314.
Cunningham CX, Williamson GJ, Bowman DMJS (2024) Increasing frequency and intensity of the most extreme wildfires on Earth. Nat Ecol Evol 8: 1420–1425. https://doi.org/10.1038/s41559-024-02452-2.
Cyr D, Splawinski TB, Puigdevall JP, Valeria O, Leduc A, Thiffault N, Bergeron Y, Gauthier S (2022) Mitigating post-fire regeneration failure in boreal landscapes with reforestation and variable retention harvesting: at what cost? Can J For Res 52: 568–581. https://doi.org/10.1139/cjfr-2021-0180.
Dawe DA, Parisien MA, Van Dongen A, Whitman E (2022) Initial succession after wildfire in dry boreal forests of northwestern North America. Plant Ecol 223: 789–809. https://doi.org/10.1007/s11258-022-01237-6.
Dong L, Leung LR, Qian Y, Zou Y, Song F, Chen X (2021) Meteorological environments associated with California wildfires and their potential roles in wildfire changes during 1984–2017. J Geophys Res Atmos 126, article id e2020JD033180. https://doi.org/10.1029/2020JD033180.
Eckdahl JA, Kristensen JA, Metcalfe DB (2023) Climate and forest properties explain wildfire impact on microbial community and nutrient mobilization in boreal soil. Front For Glob Change 6, article id 1136354. https://doi.org/10.3389/ffgc.2023.1136354.
Finney MA, McAllister SS, Forthofer JM, Grumstrup TP (2021) Wildland fire behaviour: dynamics, principles and processes. CSIRO Publishing, Clayton. https://doi.org/10.1071/9781486309092.
Gauthier S, Bernier P, Kuuluvainen T, Shvidenko AZ, Schepaschenko DG (2015) Boreal forest health and global change. Science 349: 819–822. https://doi.org/10.1126/science.aaa9092.
Greene DF, Macdonald SE, Haeussler S, Domenicano S, Noël J, Jayen K, Charron I, Gauthier S, Hunt S, Gielau ET, Bergeron Y, Swift L (2007) The reduction of organic-layer depth by wildfire in the North American boreal forest and its effect on tree recruitment by seed. Can J For Res 37: 1012–1023. https://doi.org/10.1139/x06-245.
Grillakis M, Voulgarakis A, Rovithakis A, Seiradakis KD, Koutroulis A, Field RD, Kasoar M, Papadopoulos A, Lazaridis M (2022) Climate drivers of global wildfire burned area. Environ Res Lett 17, article id 045021. https://doi.org/10.1088/1748-9326/ac5fa1.
Hunter ME, Robles MD (2020) Tamm review: the effects of prescribed fire on wildfire regimes and impacts: a framework for comparison. For Ecol Manage 475, article id 118435. https://doi.org/10.1016/j.foreco.2020.118435.
Janssen TAJ, Veraverbeke S (2025) What are the limits to the growth of boreal fires? Glob Change Biol 31, article id e70130. https://doi.org/10.1111/gcb.70130.
Jiménez-Morillo NT, Almendros G, De la Rosa JM, Jordán A, Zavala LM, Granged AJP, González-Pérez JA (2020) Effect of a wildfire and of post-fire restoration actions in the organic matter structure in soil fractions. Sci Total Environ 728, article id 138715. https://doi.org/10.1016/j.scitotenv.2020.138715.
Jones MW, Kelley DI, Burton CA, Di Giuseppe F, Barbosa MLF, Brambleby E, Hartley AJ, Lombardi A, Mataveli G, McNorton JR, Spuler FR, Wessel JB, Abatzoglou JT, Anderson LO, Andela N, Archibald S, Armenteras D, Burke E, Carmenta R, Chuvieco E, Clarke H, Doerr SH, Fernandes PM, Giglio L, Hamilton DS, Hantson S, Harris S, Jain P, Kolden CA, Kurvits T, Lampe S, Meier S, New S, Parrington M, Perron MMG, Qu Y, Ribeiro NS, Saharjo BH, San-Miguel-Ayanz J, Shuman JK, Tanpipat V, van der Werf GR, Veraverbeke S, Xanthopoulos G (2024) State of wildfires 2023–2024. Earth Syst Sci Data 16: 3601–3685. https://doi.org/10.5194/essd-16-3601-2024.
Kellman M (2004) Sugar maple (Acer saccharum Marsh.) establishment in boreal forest: results of a transplantation experiment. J Biogeogr 31: 1515–1522. https://doi.org/10.1111/j.1365-2699.2004.01128.x.
Kracke I, Essl F, Zulka KP, Schindler S (2021) Risks and opportunities of assisted colonization: the perspectives of experts. Nat Conserv 45: 63–84. https://doi.org/10.3897/natureconservation.45.72554.
Lamont BB, Pausas JG, He T, Witkowski ETF, Hanley ME (2020) Fire as a selective agent for both serotiny and nonserotiny over space and time. Crit Rev Plant Sci 39: 140–172. https://doi.org/10.1080/07352689.2020.1768465.
Macdonald SE, Gauthier S (2025) Collection: changing wildfire regimes, their impacts, and the path forward. Can J For Res 55: 1–4. https://doi.org/10.1139/cjfr-2024-0227.
MacKenzie WH, Mahony CR (2021) An ecological approach to climate change-informed tree species selection for reforestation. For Ecol Manage 481, article id 118705. https://doi.org/10.1016/j.foreco.2020.118705.
Mataix-Solera J, Cerdà A, Arcenegui V, Jordán A, Zavala LM (2011) Fire effects on soil aggregation: a review. Earth-Sci Rev 109: 44–60. https://doi.org/10.1016/j.earscirev.2011.08.002.
Matusick G, Ruthrof KX, Kala J, Brouwers NC, Breshears DD, Hardy GEJ (2018) Chronic historical drought legacy exacerbates tree mortality and crown dieback during acute heatwave-compounded drought. Environ Res Lett 13, article id 095002. https://doi.org/10.1088/1748-9326/aad8cb.
Nayomi KA, Neary B, Chen J, Mayor SJ (2022) A critical review of successional dynamics in boreal forests of North America. Environ Rev 30: 563–594. https://doi.org/10.1139/er-2021-0106.
Neina D (2019) The role of soil pH in plant nutrition and soil remediation. Appl Environ Soil Sci 2019, article id 5794869. https://doi.org/10.1155/2019/5794869.
Nelson AR, Narrowe AB, Rhoades CC, Fegel TS, Daly RA, Roth HK, Chu RK, Amundson KK, Young RB, Steindorff AS, Mondo SJ, Grigoriev IV, Salamov A, Borch T, Wilkins MJ (2022) Wildfire-dependent changes in soil microbiome diversity and function. Nat Microbiol 7: 1419–1430. https://doi.org/10.1038/s41564-022-01203-y.
NRCan (Natural Resources Canada) (2024) National wildland fire situation report. Government of Canada. https://cwfis.cfs.nrcan.gc.ca/report. Accessed 3 May 2025.
Padullés Cubino J, Vilà-Cabrera A, Retana J (2024) Tree species abundance changes at the edges of their climatic distribution: an interplay between climate change, plant traits and forest management. J Ecol 112: 2785–2797. https://doi.org/10.1111/1365-2745.14419.
Pedlar JH, McKenney DW, Aubin I, Beardmore T, Beaulieu J, Iverson L, O’Neill GA, Winder RS, Ste-Marie C (2012) Placing forestry in the assisted migration debate. BioScience 62: 835–842. https://doi.org/10.1525/bio.2012.62.9.10.
Prendergast-Miller MT, de Menezes AB, Macdonald LM, Toscas P, Bissett A, Baker G, Farrell M, Richardson AE, Wark T, Thrall PH (2017) Wildfire impact: natural experiment reveals differential short-term changes in soil microbial communities. Soil Biol Biochem 109: 1–13. https://doi.org/10.1016/j.soilbio.2017.01.027.
Previdi M, Smith KL, Polvani LM (2021) Arctic amplification of climate change: a review of underlying mechanisms. Environ Res Lett 16, article id 093003. https://doi.org/10.1088/1748-9326/ac1c29.
Reid C, Watmough SA (2014) Evaluating the effects of liming and wood-ash treatment on forest ecosystems through systematic meta-analysis. Can J For Res 44: 867–885. https://doi.org/10.1139/cjfr-2013-0488.
Ren P, Liang E, Raymond P, Rossi S (2020) Bud break in sugar maple submitted to changing conditions simulating a northward migration. Can J For Res 51: 842–847. https://doi.org/10.1139/cjfr-2020-0365.
Rossi JL, Chatelon FJ, Marcelli T (2020) Fire intensity. In: Manzello SL (ed) Encyclopedia of Wildfires and Wildland-Urban Interface (WUI) Fires. Springer International Publishing, Cham, pp 391–397. https://doi.org/10.1007/978-3-319-52090-2_51.
Swanston CW, Janowiak MK, Brandt LA, Butler PR, Handler SD, Shannon PD, Derby Lewis A, Hall K, Fahey RT, Scott L, Kerber A, Miesbauer JW, Darling L (2016) Forest adaptation resources: climate change tools and approaches for land managers, 2nd edition. Gen. Tech. Rep. NRS-GTR-87-2. U.S. Department of Agriculture, Forest Service, Northern Research Station, Newtown Square, PA.
Thiffault N, Fera J, Hoepting MK, Jones T, Wotherspoon A (2024) Adaptive silviculture for climate change in the Great Lakes- St. Lawrence Forest Region of Canada: background and design of a long-term experiment. For Chron 100: 155–164. https://doi.org/10.5558/tfc2024-016.
Thiffault N, Nordin P, Wotherspoon A, Hjelm K, Olofsson E (2025) A trans-Atlantic perspective on successful plantation establishment in boreal ecosystems: lessons learned and research opportunities. New For 56, article id 16. https://doi.org/10.1007/s11056-024-10086-2.
Wang Q, Zhong M, Wang S (2012) A meta-analysis on the response of microbial biomass, dissolved organic matter, respiration, and N mineralization in mineral soil to fire in forest ecosystems. For Ecol Manage 271: 91–97. https://doi.org/10.1016/j.foreco.2012.02.006.
Williams MI, Dumroese RK (2013) Growing assisted migration: synthesis of a climate change adaptation strategy. In: Haase DL, Pinto JR, Wilkinson KM (eds) National proceedings: forest and conservation nursery associations – 2012. Proc. RMRS-P-69. USDA Forest Service, Rocky Mountain Research Station, Fort Collins, CO, pp 90–96.
Wotherspoon A, D’Orangeville L, Thiffault N, Pedlar JH, Raymond P, Ravn J, Spearing M, Isaac-Renton M, Godbout J, Gravel-Grenier J, McPhee D. Challenges and opportunities for the operationalization of forest assisted migration in Canada. Can J For Res. In press.
Zhao B, Zhuang Q, Shurpali N, Köster K, Berninger F, Pumpanen J (2021) North American boreal forests are a large carbon source due to wildfires from 1986 to 2016. Sci Rep 11, article id 7723. https://doi.org/10.1038/s41598-021-87343-3.
Zhang Y, Biswas A (2017) The effects of forest fire on soil organic matter and nutrients in boreal forests of North America: a review. In: Rakshit A, Abhilash P, Singh H, Ghosh S (eds) Adaptive soil management: from theory to practices. Springer, Singapore, pp 391–407. https://doi.org/10.1007/978-981-10-3638-5_21.
Zou Y, Backus GA, Safford HD, Sawyer S, Baskett ML (2024) Quantifying the capacity for assisted migration to achieve conservation and forestry goals under climate change. J Biogeogr 51: 2440–2455. https://doi.org/10.1111/jbi.14999.
Total of 50 references.
