Lars-Göran Stener  ,
Johan Westin
,
Johan Westin
Early growth and phenology of hybrid aspen and poplar in clonal field tests in Scandinavia
Stener L.-G., Westin J. (2017). Early growth and phenology of hybrid aspen and poplar in clonal field tests in Scandinavia. Silva Fennica vol. 51 no. 3 article id 5656. https://doi.org/10.14214/sf.5656
Highlights
- Growth and survival was in general higher for hybrid aspen than for poplar
- The poor performance of poplar was likely due to poor climatic adaptation or to high soil acidity
- The genetic results indicate good possibilities for effective clonal selection
- The results support current recommendations for utilization of selected hybrid aspen and poplar regeneration material in Sweden.
Abstract
Results on early survival, growth and shoot phenology of hybrid aspen (Populus tremula L. × P. tremuloides Michx.) and poplar clones (P. trichocarpa Torr. & A. Gray, P. balsamifera L., P. maximowiczii A. Henry and their hybrids) in 13 Scandinavian field trials are presented. The trials were established on forest land (7 sites) or former agricultural land (6 sites) within the latitude range of 56° to 65° N and were assessed 3–4 years after establishment. The main aim was to evaluate phenotypic and genetic differences related to early survival, growth and phenology for hybrid aspen and poplar for different site types and latitudes. Growth and survival was generally higher for hybrid aspen than poplar at all sites. The poor performance of poplar compared to hybrid aspen is likely due to climatic maladaptation or high soil acidity. The early growth performance of the species need to be confirmed at a higher age. The genetic variation and genetic control for growth, phenology and survival was in general intermediate to large indicating good possibilities for effective clonal selection. The genetic site x site correlations (rGE) for growth were for hybrid aspen mostly strong, indicating a weak genotype by environment interaction, while rGE were inconsistent for poplars.The result suggests that southern Sweden can be treated as a single test and utilization zone and in northern Sweden the region along the coast may be another zone. It is too early to make any corresponding conclusions for poplar. In addition, the result backs up the current recommendations for utilization of selected hybrid aspen and poplar regeneration material in Sweden.
Keywords
survival;
Populus;
Sweden;
early test;
shoot phenology;
Denmark
-
Stener,
The Forestry Research Institute of Sweden, Ekebo 2250, 268 90 Svalöv, Sweden
E-mail
lars-goran.stener@skogforsk.se
- Westin, The Forestry Research Institute of Sweden, Box 3, 918 21 Sävar, Sweden E-mail johan.westin@skogforsk.se
Received 9 February 2017 Accepted 11 April 2017 Published 8 May 2017
Views 111503
Available at https://doi.org/10.14214/sf.5656 | Download PDF

1 Introduction
The Nordic countries have adopted a target of becoming carbon-neutral in year 2050 (Nordiska Ministerrådet 2009). High-productive forest systems and energy crops pose a potential for expanding the energy supply from both forest and agricultural land. Large areas of productive agriculture land may be used for growing biomass for biofuel in Scandinavia. The area has been estimated from 300 000 to 500 000 ha in each of Sweden and Denmark (Rytter et al. 2016). In addition, there is fertile forest land that may be suitable for biomass production as well.
Using clones of fast growing poplars has been proposed in contexts of both forestry and agriculture (Christersson 2010; Tullus et al. 2012; Rytter and Stener 2014). In Scandinavia, the Eurasian aspen (Populus tremula L.) is the only indigenous poplar species. The attention has mainly focused on introduced species like hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in the Populus section Populus and the Balsam poplars in the Populus section Tacamahaca (P. trichocarpa Torr. & A. Gray, P. balsamifera L. and P. maximowiczii A. Henry). When “species” is used further on, it refers to hybrid aspen and other poplars, respectively.
Poplar plantations in general have been shown to be one of the most efficient transformers of solar radiation energy to chemical energy in the form of wood (Christersson et al. 1987; Christersson 2006, 2010; Rytter et al. 2011). In hybrid aspen and poplars, a mean annual increment (MAI) of up to 25 m3 of stem wood ha–1 and year–1 during a rotation of around 25 years, appears to be realistic on fertile sites in southern Sweden when using the best available selected clonal material (Rytter and Stener 2014; Stener 2010). This corresponds to a dry weight of stem biomass of 8.2 tons of dry matter (DM) ha–1, yr–1 and to about 10 tons DM ha–1, yr–1 if branch biomass is included. Similar production levels have been reported by Nielsen et al. (2014) at two Danish sites but already after only 13 years, i.e. far earlier than the expected optimal rotation age.
The increasing interest in poplars in large parts of Scandinavia is due, not only to the high production potential, but also because at least hybrid aspen constitutes an alternative to reforestation with Norway spruce (Picea abies (L.) Karst.) on fertile sites (Fahlvik et al. 2009). Using several species in forestry could be a way to spread the economic risks while facing an uncertain future. Because of increasing poplar plantations (Rytter et al. 2016), there is an increasing demand for robust regeneration material, where issues like vitality, climatic adaptation and resistance to pathogens are important to consider, in addition to yield.
The Populus material of interest for utilization in Scandinavia mainly originates from North America and is adapted to climatic and photoperiodic conditions differing from that in Scandinavia. Due to an expected maladaptation, the need of genetic tests in the deployment area prior to commercial utilization is important. Large clonal variation in yield in combination with possible interactions between clone and site environment emphasize the importance of field trials at different site conditions for optimal selection of clones to be used commercially (Stener and Karlsson 2004).
The commercial material of hybrid aspen and poplar available for regeneration in Sweden and Denmark is scarce and has a narrow genetic base. Today only 15 clones of hybrid aspen and 16 clones of Balsam poplar have been selected and tested for commercial use in southern Sweden and there is no such material available at all in central and northern Sweden. In Denmark, only one hybrid poplar clone is used, OP42 (P. maximowiczii × P. trichocarpa), which is also the most commonly planted poplar clone in Sweden. Thus, there is a need to broaden the base of tested and selected material of poplars for use in Scandinavia, if fast growing poplars are intended to be a sustainable high-productive forest system.
There are many new candidate clones that, after being tested, may be of commercial interest. The Swedish University of Agricultural Sciences (SLU/SUAS) has presented promising early results from field tests with P. tricocharpa originating from northern USA and Canada (Christersson 2010). There are also Finnish hybrid aspen clones, selected on their performance in Finnish clonal tests (Yu 2001; Heräjärvi and Junkkonen 2006), that are available but not tested in Sweden. For utilization in southern Sweden and Denmark hybrid aspen clones from Germany and selected poplar clones used in central Europe may have a great potential.
Growers of poplars are often advised to use fertile mineral soils for poplar plantations (Dickmann and Kuzovkina 2008). However, the knowledge of the growth potential on forest land in relation to agricultural land is poor. Comparisons between species are often unreliable as trials of poplars and hybrid aspen, with very few exceptions, are established on separate sites. Furthermore, we need more knowledge on how the species perform on a latitudinal cline from south to north.
The main objectives with this study on hybrid aspen and poplar are to:
• estimate phenotypic variation in early survival, growth and shoot phenology
• estimate genetic variation, genetic control and genotype by environment interactions for early survival, growth and shoot phenology
• give recommendations of deployment areas in Sweden.
2 Material and methods
2.1 Material
2.1.1 General
In total 13 field trials with hybrid aspen and poplar clones were established in the years 2010 to 2012 from Denmark in the south up to northern Sweden (Table 1, Fig. 1). Six trials were planted in southern Sweden (SWE_A1–3, SWE_F1–3) in 2010, four sites in northern Sweden (SWE_A4–5, SWE_F4–5) in 2011 and the 3 sites in Denmark (DEN_A1, DEN_F1–2) in 2012. The trial sites covered approximately six climate regions as defined by degree-days and photoperiod (latitude). The geographic distance for one pair of trials (Swe_A2 and Swe_F2) was not optimal due to problems in finding appropriate trial sites. The TS values (Table 1) for the pair, still indicate similar degree-days levels and temperature conditions. The two trials within each region were considered as parallel trials, with one site located on forest land and the other on agricultural land. The only exception was the Danish climate region with two sites on forest land and one on agricultural site. In each parallel trial the same clonal material was tested. Overall, 109 clones of hybrid aspen and 117 clones of poplar were tested. All clones were grouped into 15 groups mainly depending on where they had been selected (Table 2). Unfortunately, the information of origin and pedigree is insufficient. Many of the clones were in common among the different sites as shown in Table 3. The hybrid aspen groups “Swe_S_Com”, “Swe_S_New” and “Finl” together with the poplars “Swe_S_Com” were planted at all sites.
| Table 1. Description of the trial sites within each country. The trial Id consists of country where Den = Denmark and Swe = Sweden, site type where F = Forest land and A = Agricultural land and parallel trial series number within country. TS = degree days based on daily mean temperatures >5 °C (Perttu and Moren 1995) and pH = soil acidity. | |||||||
| Trial id | Name | Site type | Planted year | Latitude, Longitude | Alt m | TS Degree days | pH |
| Den_A1 | Give | Agricultural | 2012 | 55°83´N, 09°16´E | 75 | 1552 | |
| Den_F1 | Lournkaer_Nord | Forest | 2012 | 56°72´N, 10°20´E | 1 | 1567 | |
| Den_F2 | Lournkaer-Syd | Forest | 2012 | 56°70´N, 10°14´E | 30 | 1542 | |
| Swe_A1 | Sturup | Agricultural | 2010 | 55°56´N, 13°38´E | 120 | 1466 | 5.9 |
| Swe_F1 | Matteröd | Forest | 2010 | 56°11´N, 13°62´E | 114 | 1438 | 4.2 |
| Swe_A2 | Remningstorp | Agricultural | 2010 | 58°26´N, 13°37´E | 133 | 1291 | 5.1 |
| Swe_F2 | Toftaholm | Forest | 2010 | 56°99´N, 14°05´E | 162 | 1344 | 4.8 |
| Swe_A3 | Harg | Agricultural | 2010 | 60°09´N, 18°25´E | 20 | 1276 | 7.9 |
| Swe_F3 | Karön | Forest | 2010 | 60°29´N, 18°36´E | 25 | 1267 | 7.8 |
| Swe_A4 | Ed | Agricultural | 2011 | 63°25´N, 17°23´E | 75 | 1039 | 5.2 |
| Swe_F4 | “Pommac” | Forest | 2011 | 63°11´N, 17°27´E | 265 | 888 | 4.9 |
| Swe_A5 | Degerbyn | Agricultural | 2011 | 64°78´N, 20°81´E | 20 | 993 | 4.9 |
| Swe_F5 | Degerbyn | Forest | 2011 | 64°78´N, 20°81´E | 20 | 992 | 4.4 |
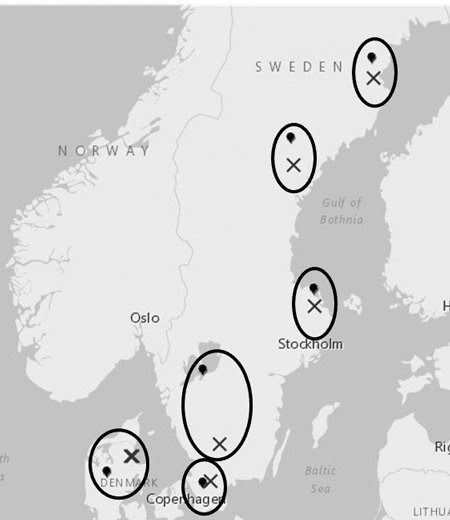
Fig. 1. Location of the trials. The parallel trials i.e. trials with similar climatic conditions and including the same test material are encircled. “X” represents forest land and the balloon symbol represents agricultural land.
| Table 2. Description of material groups per species, deployment areas and number of clones per group. “STT” = SweTree Technologies. | ||||
| Species | Main deployment area | Description | Clones N | |
| Group name | ||||
| Hybrid aspen | ||||
| Swe_S_Com | S. Sweden | Genotypically seleced clones for commercial use | 15 | |
| Swe_S_New | S. Sweden | An additional genotypic selection in trials where Swe_S_Com was tested | 11 | |
| Swe_N | N. Sweden | Phenotypic selection | 23 | |
| Finl | S. Finland | Phenotypic selection | 23 | |
| Latv | Latvia | Phenotypic selection | 22 | |
| Germ | N. Germany | Phenotypic selection | 15 | |
| Poplar | ||||
| Swe_S_Com | S. Sweden | Genotypically selected clones for commercial use (P. trichocarpa or P. maximowiczii × P. trichocarpa). Includes clone OP42 | 16 | |
| Swe_N | N. Sweden | Phenotypic selection (P. trichocarpa) | 23 | |
| Swe_STT_C | C. Sweden | Phenotypic selection STT, (P. trichocarpa) | 5 | |
| Swe_STT_N | N. Sweden | Phenotypic selection STT, (P. trichocarpa) | 9 | |
| Icel | Iceland | Phenotypic selection, (P. trichocarpa) | 7 | |
| Be | Belgium | Phenotypic selection, (P. deltoides x P. trichocarpa) | 43 | |
| Ital | Italy | Commercial clones (AF2, AF8, AF13, Monviso, AF18, AF24, AF28) | 7 | |
| EU_Com | C. Europe | Commercial clones (Grimminge, Koster and Muur). | 3 | |
| Norw | Norway | Phenotypic selection, (P. trichocarpa) | 4 | |
| Sum of clones | 226 | |||
| Table 3. Distribution of the number of hybrid aspen and poplar clones tested on different material groups and sites. See Table 2 for explanation of material groups. | |||||||||||||||
| Trial id | Material groups of Hybrid aspen | Material groups of Poplar | |||||||||||||
| Swe_S _Com | Swe_S _New | Swe _N | Finl | Latv | Germ | Swe_S _Com | Swe _N | Swe _STT_C | Swe _STT_N | Icel | Be | Ital | EU _Com | Norw | |
| Den_A1 | 15 | 11 | 4 | 15 | 6 | ||||||||||
| Den_F1 | 15 | 11 | 4 | 15 | 3 | ||||||||||
| Den_F2 | 15 | 11 | 4 | 15 | |||||||||||
| Swe_A1 | 15 | 11 | 23 | 22 | 12 | 16 | 46 | 4 | 3 | ||||||
| Swe_F1 | 15 | 11 | 23 | 22 | 12 | 16 | 46 | 4 | 3 | ||||||
| Swe_A2 | 15 | 11 | 23 | 22 | 11 | 16 | 5 | 6 | 4 | ||||||
| Swe_F2 | 15 | 11 | 23 | 22 | 11 | 16 | 5 | 6 | 4 | ||||||
| Swe_A3 | 15 | 11 | 23 | 22 | 16 | 5 | 6 | ||||||||
| Swe_F3 | 15 | 11 | 23 | 22 | 16 | 5 | 6 | ||||||||
| Swe_A4 | 15 | 10 | 23 | 23 | 4 | 13 | 5 | 3 | |||||||
| Swe_F4 | 15 | 10 | 22 | 23 | 4 | 11 | 4 | 3 | |||||||
| Swe_A5 | 15 | 10 | 23 | 23 | 15 | 22 | 3 | 7 | 4 | ||||||
| Swe_F5 | 15 | 10 | 21 | 23 | 15 | 21 | 2 | 7 | 4 | ||||||
2.1.2 Propagation
The hybrid aspen material was propagated in years 2009, 2010 and 2011 for the sites in southern Sweden, northern Sweden and Denmark, respectively. All plants were planted as one-year old container plants but different methods were used for the propagation. The Swedish material to the six southern Swedish and the three Danish sites were propagated as herbaceous root sprout cuttings at the Skogforsk research station at Ekebo (55°57´N, 13°07´E, 80 m a.s.l.). The Finnish, Latvian and German material were all micro-propagated at the Finnish Forest Research Institute (60°17´N, 25°03´E), at Silava (56°52´N, 24°21´E) and at Thȕnen Institute of Forest Genetics (52°17´N, 10°26´E), respectively. All hybrid aspen for the four northernmost sites were vegetatively propagated as root segments at the Skogforsk research station at Sävar (63°53´N, 20°33´E, 10 m a.s.l.).
All poplars apart from the Italian clones for southern Sweden were vegetatively propagated as one-year old container plants by rooting of dormant stem segments at the Skogforsk research stations at Ekebo or Sävar. The Italian material for southern Sweden was propagated in the same way but in mid-summer 2010 and planting was performed one month later than all the other test material. Thus, the Italian clones were in active growth in contrast to all other plants.
2.1.3 Sites
The forest sites were selected for high fertility and favorable climatic conditions, where obvious frost prone sites were avoided. The agricultural sites had been set aside from farming and were as such not the most fertile agricultural land for that region. Thus, the selection of sites was according to silvicultural practice for establishment of hybrid aspen and poplars. The agricultural sites were all treated with glyphosate herbicide (Roundup) at one or two occasions before planting to reduce the impact of competing weeds. In Denmark, an additional ploughing and harrowing was carried out. The forest land sites in Sweden were prepared by inverted soil scarification the year before planting by an excavator. In Denmark, a forest disc plough was used.
2.1.4 Trial design
All trials were planted in a randomized block and single tree plot design with 6 blocks at all sites except in the four northern and in the Danish sites, where 8–10 blocks were used. In Swe_F4 and Swe_A4 part of the poplar material was missing, which reduced the number of blocks for the poplars to only 3. Each block was divided into two sub plots, each including clones for either hybrid aspen or poplars. The sub-plot design was used to avoid species competition, as it was expected that the poplar would outgrow the hybrid aspen. The number of plants per clone in each block was mainly one or two and the plant spacing was 3×3 m at all sites. All trials were fenced to avoid damage by roe-deer or moose. The trials in northern Sweden (Swe_F4, Swe_F5 Swe_A5, and especially Swe_A4) were all partly damaged by voles the first summer until mechanical plant protection were in place. Also, damages by hares were evident but were not as severe as the vole damages.
2.2 Methods
2.2.1 Assessments
Individual plant assessments were performed after three (all Danish trials) and four years (all Swedish trials) of growth (Table 4). At the four southern Swedish sites height was also assessed after the first growing season. Phenology was assessed in spring and in autumn during the fourth year at four Swedish sites. Spring phenology was assessed as bud flushing in the living crown and autumn phenology was assessed as coloration or shedding of leaves. To improve the differentiation in autumn phenology among clones and species, scoring was performed at two or three occasions in the two southern trials.
| Table 4. Description of measured traits. | ||
| Trait | Unit | Description |
| Survival | (0–1) | Survival, classified as 0 = dead and 1 = alive |
| Height | cm | Total height |
| Flushing | (1–6) | Spring phenology based on flushing for the upper half of the living crown registered in 6 classes: 1 = dormant, 2 = buds started to swell, 3 = slightly broken buds where little green can be seen at the top, 4 = broken bud, 5 = developed leaves, 6 = fully developed leaves. Classification dates: Swe_A1: April 26, 2013 for hybrid aspen and May 7, 2013 for poplar. Swe_A2: May 11, 2013. Swe_A5: May 28, 2013. Swe_F5: May 24, 2013. |
| Coloration | (0–100) | Autum phenology classified by coloration of leaves and/or lost leaves in 10% classes where 0 = no autumn colors and/or no leaves lost … 100 = 100% autumn coloration and/or 100% lost leaves. Classification dates: Swe_A1: Oct 4 and 16, 2013. Swe_A2: Sep 17, 26 and Oct 8, 2013. Swe_A5: 27 Sept, 2013. Swe_F5: Sept 30, 2013. |
Soil samples were taken in the center of each block at a depth of 30 cm below the humus layer after three years of growth at all Swedish locations. A pH value, using distilled water, was measured for each sample according to Karltun (1996), from which a mean for each site was estimated.
2.2.2 Genetic analysis
The genetical-statistical analysis was based on individual tree observations for each species (hybrid aspen and poplars) separately according to model:
![]()
where yijk = observation k, in block i for clone j, μ = trial mean, bi = fixed effect of block i, cj = random effect of clone j with NID(0,σc2) and eijk = random error term for observation ijk, NID(0,σe2).
To investigate the effect of country where the material had been selected, an additional fixed variable dl = country was included in Eq. 1. Survival was the only trait where data deviated from normal distributions and were transformed to normal scores prior to analysis according to Gianola and Norton (1981). However, since the effect on the result was very marginal, the original untransformed values were kept.
The variances σc2 and σe2 were estimated for different traits according to the REML (Restricted Maximum Likelihood) method, as performed in ASREML software (Gilmour et al. 1999). Genetic parameters for clones were interpreted as σG2 = σc2 and σE2 = σe2, where σG2 = the genotypic variance among clones and σE2 = environmental variance.
The individual-tree broad-sense heritabilities (H2) were calculated as
![]()
where phenotypic variance (σ2P) was σ2G + σ2E
The genotypic (CVG) coefficients of variation were calculated as
![]()
where ![]() is the phenotypic mean.
is the phenotypic mean.
2.2.3 Genetic correlations
Interactions between genotype and environment (GxE) was analysed as genetic site x site correlations, here denoted as rGE, between the same trait measured on two sites, using the following model, expressed in matrix notation:
![]()
where i pertains to the same trait measured in two different trials, yi is the vector of individual tree observations, b is the vector of fixed block effects, c is the vector of random clone effects and e is the vector of random residuals. X and Z are design matrices of the block effects and clone effects, respectively. The random effects are assumed to have a multivariate, normal distribution with expectation zero and may be summarized as ![]() and
and ![]() . The variance-covariance matrix is assumed to be
. The variance-covariance matrix is assumed to be

where G is the matrix with the clonal variances and covariances, R is the matrix with the residual variances and covariances and I is an identity matrix. When traits are measured on different sites, R is reduced to a diagonal matrix including the residual variances only. Finally, ![]() symbolises the direct product.
symbolises the direct product.
The genotypic (rG) correlations between traits within sites were estimated as
![]()
where σG1G2 is the genotypic covariance between two traits, respectively. The genetic site x site correlations (rGE) were evaluated in pairs of trials in the same way as rG. Standard errors of the parameter estimates (H2, rG, rGE) were calculated by first-order Taylor series approximations by the ASREML software.
2.2.4 Phenotypic analysis
An additional statistical analysis, based upon subplot means, was performed by Proc Glm (SAS Institute Inc. 2011) where hybrid aspen and poplars as well as the different material groups within each species were compared. The following model was used:
![]()
where yij = observation ij, μ = mean value, bi = fixed effect of block i, pj = group or species j, eij = random error term for observation ij, NID(0,σe2). For test of significance (p < 0.05) the Tukey studentized range test was used (SAS Institute Inc. 2011).
For test of significant differences between parallel sites, i.e. between forest land and agricultural land individual values with the following model was used:
![]()
where yijk = observation ijk, μ = mean value, si = fixed effect of site i, bj = fixed effect of block j, eijk = random error term for observation ijk, NID(0,σe2).
3 Results
3.1 Phenotypic evaluation of survival, growth and phenology
The survival rates among the sites, based on the mean for all clones included in each trial, varied for hybrid aspen and poplar between 50–95% and 0–89%, respectively (Table 5). The poplars showed generally lower survival than hybrid aspen, with significantly lower (p < 0.05) survival on 8 of the 13 sites. The survival of poplar was extremely poor at one southern site (11% in Swe_F1) and at all four northern sites (<39%).
| Table 5. Average survival (%), height (cm), flushing score (1–6) and autumn coloration (%) of hybrid aspen (Hasp) and poplar (Popp) in different trials. The significance probability value associated with species is given by ”Pr > F” for each trait and site. | ||||||||||||||||
| Trial id | Survival, % | Height, cm | Flushing (1–6) | Coloration, % | ||||||||||||
| Hasp | Popp | All | Pr > F | Hasp | Popp | All | Pr > F | Hasp | Popp | All | Pr > F | Hasp | Popp | All | Pr > F | |
| Den_A1 | 91 | 89 | 91 | 0.6209 | 277 | 258 | 268 | 0.0391 | ||||||||
| Den_F1 | 94 | 70 | 86 | 0.0018 | 401 | 166 | 334 | <0.0001 | ||||||||
| Den_F2 | 94 | 89 | 93 | 0.1295 | 289 | 189 | 254 | <0.0001 | ||||||||
| Swe_A1 | 92 | 69 | 84 | 0.0018 | 351 | 295 | 335 | 0.0178 | 4.4 | 3.3 | 4.0 | 0.0024 | 67 | 49 | 62 | 0.0003 |
| Swe_F1 | 83 | 11 | 59 | <0.0001 | 281 | 100 | 270 | <0.0001 | ||||||||
| Swe_A2 | 95 | 52 | 85 | <0.0001 | 517 | 447 | 508 | 0.1063 | 3.9 | 2.7 | 3.7 | 0.0030 | 59 | 71 | 60 | 0.0045 |
| Swe_F2 | 82 | 76 | 81 | 0.1611 | 270 | 161 | 244 | 0.0007 | ||||||||
| Swe_A3 | 77 | 76 | 77 | 0.6983 | 270 | 260 | 268 | 0.0828 | ||||||||
| Swe_F3 | 93 | 83 | 90 | 0.0269 | 236 | 233 | 235 | 0.8777 | ||||||||
| Swe_A4 | 50 | 0 | 37 | 0.0255 | ||||||||||||
| Swe_F4 | 69 | 33 | 60 | 0.0690 | 192 | 38 | 152 | 0.0042 | ||||||||
| Swe_A5 | 58 | 19 | 38 | <0.0001 | 163 | 128 | 145 | 0.0203 | 5.3 | 5.5 | 5.4 | 0.1592 | 46 | 53 | 50 | 0.1428 |
| Swe_F5 | 82 | 38 | 60 | <0.0001 | 258 | 122 | 190 | <0.0001 | 5.8 | 5.7 | 5.8 | 0.3380 | 33 | 58 | 45 | 0.0047 |
The mean height growth after three (Danish sites) and four years (Swedish sites) varied between 163 to 517 cm for hybrid aspen and between 38 to 447 cm for poplars (Table 5). The highest growth for hybrid aspen was found in Swe_A2 and Den_F1 and for poplar in Swe_A2, where the average growth was more than one meter per year. Growth performance was higher (p < 0.05) for hybrid aspen at nine of the twelve sites and with a similar tendency on the remaining three sites. Mean height growth for hybrid aspen at the northernmost site (Swe_F5) was similar to several sites in southern Sweden, while for poplar, growth was generally quite poor in the north.
Hybrid aspen flushed earlier (p < 0.05) than poplar at the two southern sites, whereas no difference was evident at the northern sites (Table 5). It shall be notified that flushing in Swe_A1 was assessed eleven days later for poplar as for hybrid aspen (Table 4). For autumn coloration, no clear pattern was evident.
For comparison of the performance of hybrid aspen and poplar at parallel sites (Table 6), only clones considered relevant were included. Thus, on the southern sites all hybrid aspen and poplars clones from the “Swe_S_Com” groups were included. For the northern sites, all hybrid aspen clones from the “Swe_N” group were included, while the 16 most northerly poplar clones in Swe_N were used (Table 2). Survival for hybrid aspen differed at 5 of the 7 parallel trials (p < 0.05), where survival was higher at two agricultural sites and higher at three forest sites. Hybrid aspen had higher growth at four agricultural sites and at two forest sites (p < 0.05). For poplars, growth was higher on agricultural land at all four sites where a relevant comparison was possible, while survival differed (higher/lower) at two sites (p < 0.05).
| Table 6. Comparison of survival and growth within parallel trials, i.e. between agricultural land (Agri) and forest land (Forest) for sites within similar climatic conditions. Only clones belonging to the groups “Swe_S_Com” are included in the Danish and the southern Swedish sites, while the northern sites include all hybrid aspen clones from the “Swe_N” group and the 16 most northerly poplar clones in “Swe_N”. The statistical significance between each of the two compared sites is given by ”Pr > F”. | ||||||||||||||
| Site | Hybrid aspen | Poplar | ||||||||||||
| Survival, % | Height, cm | Survival, % | Height, cm | |||||||||||
| Agri | Forest | Pr > F | Agri | Forest | Pr > F | Agri | Forest | Pr > F | Agri | Forest | Pr > F | |||
| Den_A1_F1 | 91 | 93 | 0.4053 | 265 | 394 | <0.0001 | 93 | 70 | <0.0001 | 259 | 173 | <0.0001 | ||
| Den_A1_F2 | 91 | 93 | 0.5641 | 265 | 275 | 0.3590 | 93 | 89 | 0.3387 | 259 | 188 | <0.0001 | ||
| Swe_A1_F1 | 90 | 77 | 0.0005 | 376 | 281 | <0.0001 | ||||||||
| Swe_A2_F2 | 98 | 81 | <0.0001 | 530 | 290 | <0.0001 | 47 | 82 | <0.0001 | 446 | 178 | <0.0001 | ||
| Swe_A3_F3 | 79 | 92 | 0.0001 | 282 | 258 | 0.0063 | 70 | 80 | 0.0556 | 264 | 236 | 0.0068 | ||
| Swe_A4_F4 | 55 | 76 | <0.0001 | 211 | 169 | <0.0001 | ||||||||
| Swe_A5_F5 | 71 | 85 | 0.0005 | 165 | 244 | <0.0001 | ||||||||
The results of survival and height growth for the different groups are summarized per species in Table 7. For hybrid aspen, the German group showed high growth performance in the five southernmost trials (Table 7b), while survival was the poorest (75–78%, Table 7a) for the two most northern of these sites. Survival of the Finnish material (Finl) was generally not different from the other groups in southern Sweden but were among the best in northern Sweden. Height growth for “Finl” was lower than average at four out of the six sites in southern Sweden, while growth did not differ from other groups in the northern sites. The southern Swedish hybrid aspen groups (Swe_S_Com, Swe_S_New) had in general poor survival in the northern sites, but for height growth, at least Swe_S_Com belonged to the best growing groups. For poplar in the southern sites, survival was poor for the Italian group, except in trial Den_F2 (Table 7a). Poor survival (47%) was also found for the commercial clones (Swe_S_Com) at the southern site Swe_A2. For height growth, there was no significant differences at any of the southern sites, if the Italian group was excluded. In the northern sites, survival was generally poor for all groups but the Icelandic (Icel) group belonged to the better ones, both for survival and growth.
| Table 7a. Average survival of different groups of hybrid aspen and poplar in different trials. The statistical significance of differences between the groups is given by Pr > F. Table 7b. Average height (cm) of different groups of hybrid aspen and poplar in different trials. The statistical significance of differences between the groups is given by Pr > F. View in new window/tab. |
3.2 Genotypic evaluation
The heritability (H2) estimates for survival were in general low for hybrid aspen and intermediate for poplars (Table 8). Trial Swe_A2, Swe_A5 and Swe_F5 differed from the general hybrid aspen pattern by higher survival estimates (H2 > 0.15), whereas for poplars all trials had H2 > 0.15 except from the most southern ones (Danish and Swe_F1). The heritability for height growth was in general intermediate for both hybrid aspen and poplars. The estimated coefficients of variation were at a similar level for both hybrid aspen and poplars.
| Table 8. Summary of means, broad sense heritabilities (H2) and genotypic coefficients of variation (CVG) for survival, height, flushing and coloration per site and species. “N_Obs” = Total number of observations, “N_Clone” = Number of clones, “SE” = Standard error of heritability. | ||||||||||||
| Trial id | Hybrid aspen | Poplar | ||||||||||
| N_Obs | N_Clone | Mean | H2 | SE | CVG | N_Obs | N_Clone | Mean | H2 | SE | CVG | |
| Survival (%) | ||||||||||||
| Den_A1 | 244 | 30 | 91 | 0.02 | 0.04 | 170 | 21 | 89 | 0.08 | 0.07 | ||
| Den_F1 | 254 | 30 | 94 | 0.06 | 0.04 | 146 | 18 | 70 | 0.03 | 0.05 | ||
| Den_F2 | 252 | 30 | 94 | 0.04 | 0.04 | 163 | 15 | 89 | 0.03 | 0.05 | ||
| Swe_A1 | 706 | 83 | 92 | 0.10 | 0.03 | 363 | 66 | 69 | 0.20 | 0.06 | ||
| Swe_F1 | 703 | 83 | 83 | 0.03 | 0.03 | 346 | 31 | 11 | 0.03 | 0.03 | ||
| Swe_A2 | 697 | 82 | 95 | 0.21 | 0.05 | 220 | 31 | 52 | 0.41 | 0.08 | ||
| Swe_F2 | 698 | 82 | 82 | 0.01 | 0.02 | 222 | 31 | 76 | 0.32 | 0.08 | ||
| Swe_A3 | 641 | 71 | 77 | - | - | 224 | 27 | 76 | 0.37 | 0.09 | ||
| Swe_F3 | 623 | 71 | 93 | 0.02 | 0.02 | 216 | 27 | 83 | 0.25 | 0.08 | ||
| Swe_A4 | 608 | 71 | 50 | 0.08 | 0.04 | |||||||
| Swe_F4 | 599 | 70 | 69 | 0.06 | 0.03 | 108 | 22 | 33 | 0.15 | 0.10 | ||
| Swe_A5 | 552 | 71 | 58 | 0.23 | 0.05 | 395 | 51 | 19 | 0.31 | 0.06 | ||
| Swe_F5 | 572 | 69 | 82 | 0.22 | 0.06 | 399 | 50 | 38 | 0.16 | 0.05 | ||
| Height (cm) | ||||||||||||
| Den_A1 | 229 | 30 | 277 | 0.23 | 0.07 | 12.3 | 151 | 21 | 258 | 0.14 | 0.08 | 8.7 |
| Den_F1 | 240 | 30 | 401 | 0.26 | 0.07 | 11.0 | 102 | 18 | 166 | - | - | - |
| Den_F2 | 239 | 30 | 289 | 0.32 | 0.08 | 13.7 | 146 | 15 | 189 | 0.25 | 0.09 | 14.1 |
| Swe_A1 | 647 | 83 | 351 | 0.26 | 0.05 | 16.3 | 250 | 66 | 295 | 0.28 | 0.06 | 14.8 |
| Swe_F1 | 580 | 83 | 281 | 0.23 | 0.05 | 16.5 | 39 | 28 | 100 | - | - | - |
| Swe_A2 | 653 | 82 | 517 | 0.20 | 0.04 | 11.5 | 110 | 31 | 447 | 0.26 | 0.11 | 13.9 |
| Swe_F2 | 569 | 82 | 270 | 0.38 | 0.05 | 17.7 | 167 | 31 | 161 | 0.23 | 0.09 | 16.8 |
| Swe_A3 | 477 | 71 | 270 | 0.20 | 0.05 | 12.9 | 167 | 27 | 260 | 0.27 | 0.09 | 12.4 |
| Swe_F3 | 579 | 71 | 236 | 0.12 | 0.04 | 14.5 | 179 | 27 | 233 | 0.11 | 0.06 | 10.5 |
| Swe_A4 | 301 | 71 | 221 | 0.23 | 0.06 | 16.3 | ||||||
| Swe_F4 | 412 | 70 | 192 | 0.31 | 0.06 | 21.8 | 36 | 22 | 38 | 0.25 | 0.28 | 20.0 |
| Swe_A5 | 322 | 64 | 163 | 0.22 | 0.07 | 17.5 | 74 | 25 | 128 | 0.04 | 0.21 | 9.1 |
| Swe_F5 | 466 | 63 | 258 | 0.28 | 0.06 | 17.2 | 151 | 37 | 122 | 0.00 | - | - |
| Flushing (1–6) | ||||||||||||
| Swe_A1 | 531 | 83 | 4.4 | 0.39 | 0.05 | 20.1 | 232 | 66 | 3.3 | 0.77 | 0.04 | 23.8 |
| Swe_A2 | 631 | 82 | 3.9 | 0.72 | 0.04 | 14.9 | 107 | 31 | 2.7 | 0.35 | 0.12 | 11.6 |
| Swe_A5 | 337 | 64 | 5.2 | 0.19 | 0.08 | 14.7 | 85 | 29 | 5.5 | 0.46 | 0.12 | 16.8 |
| Swe_F5 | 468 | 63 | 5.8 | 0.00 | - | - | 201 | 41 | 5.8 | 0.03 | 0.06 | 2.7 |
| Coloration (%) | ||||||||||||
| Swe_A1 | 611 | 83 | 67 | 0.81 | 0.03 | 37.4 | 235 | 66 | 49 | 0.68 | 0.05 | 44.1 |
| Swe_A2 | 639 | 82 | 59 | 0.71 | 0.04 | 36.5 | 107 | 31 | 70 | 0.75 | 0.07 | 29.6 |
| Swe_A5 | 325 | 64 | 46 | 0.57 | 0.06 | 42.0 | 79 | 27 | 53 | 0.35 | 0.16 | 25.5 |
| Swe_F5 | 468 | 63 | 33 | 0.43 | 0.06 | 40.0 | 177 | 41 | 58 | 0.19 | 0.09 | 18.2 |
The H2 estimates for the phenology traits in southern Sweden were generally high for both hybrid aspen and poplars, while H2 was lower at the northern sites. Autumn phenology was scored at one to three occasions per trait in the two southern sites and only once in the two northern sites, but only estimates from the date with the highest H2 are presented for each site in Tables 8 to 10.
The genetic correlations (rG) between growth and phenology traits within sites were mainly intermediate and significant for hybrid aspen while poor and not significant for poplar (Table 9). For growth x flushing, the rG estimates were all positive while growth x coloration was negative, i.e. early flushing and late coloration were correlated with good height performance.
| Table 9. Genetic correlations (rG) and their standard errors (SE) between height and phenology traits within site. Bold figures are significant at the 5% level. | ||||
| Trial id | Hybrid aspen | Poplar | ||
| rG | SE | rG | SE | |
| Height × Flushing | ||||
| Swe_A1 | 0.58 | 0.11 | 0.24 | 0.17 |
| Swe_A2 | 0.52 | 0.11 | 0.37 | 0.32 |
| Swe_A5 | 0.67 | 0.17 | ||
| Height × Coloration | ||||
| Swe_A1 | –0.70 | 0.08 | 0.05 | 0.19 |
| Swe_A2 | –0.51 | 0.12 | –0.15 | 0.28 |
| Swe_A5 | –0.46 | 0.18 | –0.11 | 0.75 |
| Swe_F5 | –0.46 | 0.15 | ||
The genetic age-age correlations (rG) between tree height at age 1 and 4 at the four southern Swedish sites were for hybrid aspen strong and above 0.7 at three of the sites, whereas for poplar no clear pattern was evident (data not presented).
The estimation of genetic correlations (rGE) between sites (Table 10) were in general based on clonal material in common. However, when present, Belgian and Italian poplar clones were excluded to create a more homogenous material. The evaluation of genetic correlations for the Danish sites could not be carried out due to the low number of clones. It shall be noted that the missing rGE estimates in Table 10 refers to none-estimable correlations, mainly due to low H2 estimates for one or both sites or few clones in common between the two sites. Furthermore, there are two rGE estimates of 1.00 where the standard errors are missing and they refer to evaluations where the genetic-statistical model (Eq. 4) did not converge. For hybrid aspen, the significant rGE estimates for height growth and survival were all positive and varied between 0.62–0.98 and 0.47–0.85, respectively. For poplar, the only two significant rGE estimates for height growth were contrasting; –0.86 and 0.82, while the rGE estimates for survival were all positive and those being significant varied between 0.69–0.99. Between southern and northern sites the rGE estimates for survival and growth were mostly non-significant (data not presented), except for hybrid aspen at site Swe_A5 where the genetic correlation with the southern sites were positive and varied between 0.53 and 0.78. The phenology traits had strong rGE values (>0.82) except for poplars in the northern trial (data not presented).
| Table 10. Genetic site x site correlations (rGE) for survival and height for hybrid aspen and poplar. Bold figures are significant at the 5% level. “SE” = Standard error. | |||||||||||
| Trial 1 | Trial 2 | Hybrid aspen | Poplar | ||||||||
| Survival | Height | Survival | Height | ||||||||
| rGE | SE | rGE | SE | rGE | SE | rGE | SE | ||||
| Swe_A1 | Swe_F1 | 0.78 | 0.09 | ||||||||
| Swe_A1 | Swe_A2 | 0.47 | 0.19 | 0.88 | 0.08 | 0.44 | 0.34 | –0.86 | 0.21 | ||
| Swe_A1 | Swe_F2 | 0.85 | 0.07 | 0.99 | 0.45 | –0.29 | 0.37 | ||||
| Swe_A1 | Swe_F3 | 0.77 | 0.13 | 0.54 | 0.33 | 0.41 | 0.40 | ||||
| Swe_A1 | Swe_A3 | 0.98 | 0.07 | 0.69 | 0.25 | –0.54 | 0.30 | ||||
| Swe_F1 | Swe_A2 | 0.76 | 0.11 | ||||||||
| Swe_F1 | Swe_F2 | 0.80 | 0.08 | ||||||||
| Swe_F1 | Swe_F3 | 0.64 | 0.16 | ||||||||
| Swe_F1 | Swe_A3 | 0.86 | 0.11 | ||||||||
| Swe_A2 | Swe_F2 | 0.75 | 0.10 | 0.50 | 0.35 | 0.17 | 0.34 | ||||
| Swe_A2 | Swe_F3 | 0.62 | 0.16 | 0.84 | 0.13 | 0.25 | 0.39 | ||||
| Swe_A2 | Swe_A3 | 0.86 | 0.10 | 0.76 | 0.14 | 0.82 | 0.21 | ||||
| Swe_F2 | Swe_F3 | 0.75 | 0.13 | 0.80 | 0.38 | 0.61 | 0.33 | ||||
| Swe_F2 | Swe_A3 | 0.81 | 0.10 | 0.84 | 0.30 | 0.31 | 0.29 | ||||
| Swe_F3 | Swe_A3 | 0.67 | 0.17 | 0.88 | 0.12 | 0.51 | 0.31 | ||||
| Swe_A4 | Swe_F4 | 0.70 | 0.24 | 1.00 | |||||||
| Swe_A5 | Swe_F5 | 0.85 | 0.12 | 0.78 | 0.14 | 0.71 | 0.16 | ||||
| Swe_A5 | Swe_A4 | 0.56 | 0.22 | 0.85 | 0.14 | ||||||
| Swe_A5 | Swe_F4 | 0.90 | 0.10 | 0.03 | 0.40 | 1.00 | |||||
| Swe_F5 | Swe_A4 | 0.51 | 0.23 | 1.00 | |||||||
| Swe_F5 | Swe_F4 | 0.30 | 0.27 | 0.86 | 0.12 | 0.55 | 0.40 | ||||
4 Discussion
4.1 Phenotypic data – survival, growth, phenology
Overall, hybrid aspen showed a higher survival rate and a higher growth performance than poplar on the studied sites. Survival was extremely poor for poplars at all four northern sites (<39%). Exclusion of less relevant material groups in different climatic regions, did not change this general result (Table 5, 6).
Competition from herbaceous species is a major concern when establishing broad-leaved species, especially on agricultural land (Czapowskyj and Safford 1993). Besides increasing competition for light, water and nutrients, vegetation can also cause mechanical damage to the plants. Furthermore, competing vegetation provides a favorable environment for voles which can cause severe damage in young plantations (Hytönen and Jylhä 2008). Vole damages and to some extent damages caused by hares were an important factor at the northern sites, especially at Swe_A4, where vegetation was substantial, despite herbicide treatment the year prior to planting. Vole damages could explain the poor result in survival at the northern sites. However, there were no clear differences in vole damage between poplar and hybrid aspen.
Soil pH can impact growth as it may influence the availability of the nutrients. In this study, the soil pH obtained from the Swedish trials, varied between 4.9 to 7.9 on agricultural land and between 4.2 to 7.8 on forest land (Table 1). The lowest pH-value (4.2) in the study was found in Swe_F1, where the survival of poplar in comparison with the other southern sites was extremely low (11%). This site was well prepared before planting by inverted soil scarification and there was only minor damage by hare and vole, so the most likely explanation to the low survival, is the soil acidity. Low soil pH (4.4) is most likely also an important reason for poor survival at site Swe_F5, in addition to a location in an area with an overall harsh climate. The soil acidity for these two sites did not seem to have any obvious impact on hybrid aspen, at least not so far, since survival and growth performance was similar to the other neighboring sites on forest land. Hjelm and Rytter (2016) showed that soil acidity had a significant effect on the development of poplar plants. Thus, height and biomass increased with increasing pH. An impact of soil pH was also shown by Eriksson and Lindsjö (1981), where root and shoot growth for poplars and Salix was restrained for pH values 3.8–4.3. A pH value above 5.0 seemed to result in acceptable growth, which is also in consistence to site recommendations for poplars in Canada (Boysen and Strobl 1991). The high pH-value on the forest site, Swe_F3, indicates a different land utilisation previous that of the Norway spruce stand, which was clear-cut prior the establishment of the trial.
Another reason for high mortality is poor climatic adaptation. Adaptability is a complex trait that refers to general vitality, frost tolerance, cold hardiness, resistance to pathogens and robustness over a wide range of environments. Climatic adaptation is emphasized at northern latitudes and especially for introduced species such as poplars, most likely not fully adapted to the present climate. The identification of varieties with a relevant timing of budburst and growth cessation is crucial for a high growth performance in the boreal regions (Hannerz 1998) and will probably be more important in respect of the climate change (Brázdil et al. 2010; Jönsson and Bärring 2011).
Assessments of survival in late autumn of the first growing season in all southern Swedish sites (data not presented), showed a high survival (88–100%) for both species. After that, survival was significantly reduced as demonstrated in Swe_A2 and Swe_A3. Survival for both hybrid aspen and poplar decreased here from 100% year 1, to 96% and 80% for hybrid aspen and to 55% and 79% for poplars, respectively, in year 2. Thereafter, survival rates were stable (Table 5). Thus, it seems that the winter between the first and second growing season had large impact on survival, indicating that it is related to climatic adaptation.
Climate conditions are quite varying for the trials in the study since they are located from latitude 55.8° to 64.8°, still no clear trend from south to north for growth nor survival was found for the material groups in common for all sites. The commercial material of hybrid aspen and poplar is currently recommended to be used on mild locations in southern Sweden up to latitude 60°. This recommendation is supported by the result from this study. The results also support a use of these hybrid aspen and poplar clones in Denmark.
Currently, no explanation exists for the low survival of the commercial poplar group (Swe_S_Com) at site Swe_A2 (Table 7a). Only specific clones (8 out of 16) showed low survival and only at this site. Thus, at more southerly and northerly sites these clones showed high survival. No relation was evident between survival and phenology and the plant material was treated and randomized in the same way as for all other materials and sites.
Finnish hybrid aspen is currently recommended for use at mild sites in northern Sweden based on field tests and selections performed in Finland. The results in this study support this recommendation, at least for latitudes 60° to 65°, since survival was among the best and growth was not significantly different from the more northerly Swedish material groups. Finnish hybrid aspen material is currently also used in southern Sweden, which seems to be a less optimal choice (Table 7b). German hybrid aspen material and Italian poplar, may provide alternatives in Denmark and the very southern part of Sweden but it is too early to make any final recommendations (Table 7a, b).
It is also too early to make any recommendations for utilization of poplars in northern Sweden, but specific clones within the materials from northern Sweden, Norway and Iceland are so far, the most promising ones. However, based on this study, it is obvious that poplars selected for use in southern Sweden are not a suitable choice for northern Sweden (Table 7a).
When agricultural and forest land were compared within the same climate region, the growth performance was in general higher on agricultural land than on forest land (Table 6). The result is reasonable as the site fertility is higher on agricultural land. However, this is not always the case, as is shown for hybrid aspen at the forest site Swe_F5, located in a south-facing slope, compared to the nearby but more frost prone agricultural site at Swe_A5. Frost related damages are probably the main reason for the diverging result. As large areas of productive agriculture land may be considered for growing biomass for biofuel in Scandinavia, potential frost prone sites must be avoided, if possible.
The Den_F1 site had a superior growth for hybrid aspen, a growth performance as high as Swe_A2. The soil type is marine sand-clay (GEUS 2016) which together with the very low elevation indicate a good water supply. The forest soil is, however, expected to be acidic, which seems to have reduced the growth of poplar considerably.
It is surprising that growth of the hybrid aspen recommended in southern Sweden (Swe_S_Com, Swe_S_New) is quite high and not statistically different from the best material groups at the most northern site, Swe_F5, Degerbyn (Table 7b). This is also the case for survival, at least for Swe_S_Com (Table 7a), and indicates that hybrid aspen is not very sensitive to northern latitude transfer, if the local plantation site is favorable.
It shall be noted that the Finnish, Latvian and German hybrid aspen clones were not propagated in the same nursery as the Swedish material. Differences in cultivation may have influenced the establishment and growth of the plants in the trials. Most likely this effect is rather small since the plant material seemed to be quite homogenous at the time of planting, except for the Italian poplar clones which in the Swedish sites were planted in active growing conditions. Therefore, the results for the Italian clones are more unreliable at the Swedish sites but not at the Danish sites.
The groups include quite an inhomogeneous clonal material, especially for poplars, which complicates comparisons of the phenology traits for the two species. Thus, flushing and coloration showed significant group effects in southern as well as in northern trials (not presented). This may explain that no clear difference in autumn coloration was evident between the species. Still it is quite clear from the two southern trials (Table 5), that hybrid aspen initiates growth earlier than poplars. The late developmental stage at which flushing was observed at the northern sites, indicated by the high average scores, may conceal an actual difference in timing of flushing between the species.
4.2 Genotypic data
4.2.1 Genetic control
For growth, the heritability (H2) estimates generally showed intermediate genetic control for both species (Table 8). Poplars at the northern sites diverged from this result and this is likely due to the low survival, resulting in a statistically poor material as indicated by the standard errors for H2.
The reduction of H2 by introducing country of selection in Eq. 1 was marginal for all traits and trials except for height at Swe_A1 and Swe_F2, where H2 was reduced from 0.26 to 0.15 and from 0.38 to 0.27 respectively. The estimation of H2 can be biased upwards when material of very different origins is included, but the influence of this effect on the main results in this study was considered to be of minor importance. This study confirms results from previous studies of intermediate to strong clonal control and large clonal variations. The H2 estimates were in agreement with many other studies (Einspahr et al. 1963, 1967; Ilstedt 1996; Stener and Karlsson 2004). The genetic variation among clones for growth (CVG, Table 8) was substantial and was of the same magnitude as for instance in Stener and Karlsson (2004). The overall conclusion is that genetic variation together with genetic control is of such magnitude that there are good possibilities for an effective clonal selection.
Previous studies on hybrid aspen in southern Sweden have found a weak genetic control for survival (Stener and Karlsson 2004), which is supported by our results (Table 8). This indicates that survival, at least based on the material tested so far, is not a key trait for clonal selection and further breeding in Denmark and the southern parts of Sweden. It is the opposite way in northern Sweden, where climatic conditions are harsher. Survival is for instance an essential breeding trait for Norway spruce, Scots pine and Silver birch in these regions (Rosvall et al. 2001), which also is suggested from our results at the most northern sites Swe_A5 and Swe_F5 (Table 8). The intermediate to strong genetic control for survival of poplar on most of the sites is quite logic since poplars are an entirely introduced species adapted to conditions differing from those in Scandinavia. There were some exceptions though. In Swe_F1, where survival for poplars was 11% probably due to the low pH, H2 was close to 0, i.e. all clones died regardless of clonal origin. Poor genetic control for survival was also found in the Danish sites, but this was likely influenced by the rather few number of clones tested, as indicated by the standard errors for H2.
For phenology, the results showed a pronounced clonal effect for growth initiation and termination (Table 8). Some studies have shown phenology traits to be normally highly heritable (e.g. Hannerz 1998; Pliura et al. 2014). Other studies have shown almost no clonal differences for flushing but large differences for budset (e.g. Howe et al. 2000), but this cannot be confirmed in the present study. The weak genetic control for flushing in Swe_A5 and Swe_F5 can be explained by too late field registrations, reducing the resolution among clones.
4.2.2 Genetic correlations
The genetic correlation between growth and flushing for hybrid aspen (Table 9) suggest that clones with early flushing have better growth, whereas the correlation between growth and autumn coloration showed that clones with less coloration, i.e. terminating growth late, have better growth. For poplars, the correlations were all weak and not significant and no conclusions can be drawn. This result was probably due to low survival (in the north) and few tested clones (in the south). The correlations for hybrid aspen were intermediate and consistent for the southern and the northern sites. The results suggest that a long growing period to some extent results in increased growth.
Similar results were obtained by Pliura et al. (2014), Yu (2001) and Ceulemans et al. (1992). This implies that a selection based merely on growth will increase the risk of frost damage since a long growth period is linked with late growth cessation and to some extent with early flushing. On the other hand, if growth starts too late or stops too early the growing season will not be optimally used, reducing the growth potential. Thus, growth rhythm is an essential trait to consider at practical breeding and need to be even more emphasized in the future due to the consequences of the climate change (Langvall 2011). The strong genetic control as well as the large genetic variation for the phenology traits is promising in this respect.
The results for genetic age-age correlations between tree height at age 1 and 4 suggest that the first year of growth is only little or even negatively related to the growth after 4 years for poplars. Similar result was for instance presented by Ilstedt (1996). This is once more an indication of poor adaptation and indicates longer test periods for poplar than for hybrid aspen.
For growth, the genetic correlation (rGE) for hybrid aspen among the southern sites and among the northern sites were all positive and mainly strong (Table 10). The results suggest that the growth performance of the clones is similar in different environments and that clonal rankings are quite stable across sites. For poplar, the rGE estimates were inconsistent and seldom statistically significant. The results for poplars are mainly due to a low number of tested clones in combination with a weak genetic correlation among the sites. The rGE for growth for hybrid aspen, between all the southern Swedish sites and the most northern site Swe_A5, varied from 0.53 to 0.78, and indicate an intermediate stability in clonal ranking and supports the discussion earlier, that hybrid aspen is not very sensitive to northern latitude transfer if the local plantation site is favorable.
The phenological traits were stable across environments as suggested by the strong rGE estimates. The results support previous findings (e.g. Hannerz 1998).
Other studies of genotype by environment (GxE) interactions for hybrid aspen and poplar in northern Europe are inconsistent in respect of genetic site interaction effects. In a study by Yu and Pulkkinen (2003) GxE for growth was evaluated for 25 clones planted at the age of 4 on two agricultural and two forest sites in Finland. The ranking of clones differed significantly between site types, indicating that GxE interactions were present. However, the correlation between the two agricultural sites was quite strong (0.62 for height and 0.72 for diameter). A strong GxE interaction was also found in a study of two hybrid poplar clonal tests in Lithuania (Pliura et al. 2014). On the other hand, only a weak GxE interaction for growth was found by Stener and Karlsson (2004) from four pairs of trials including up to 100 hybrid aspen clones on agricultural land in southern Sweden. Furthermore, in a Danish study of 36 poplar clones on two agricultural sites, no significant interactions were found (Nielsen et al. 2014). The latter findings support our results and indicate that at least agricultural land in southern Sweden can be treated as a single test and utilization zone for hybrid aspen. This will simplify future hybrid aspen breeding programs, if growth is the main breeding objective in the deployment zone. Furthermore, northern Sweden, at least the coastal region up to latitude 65, can be treated as one utilization zone. However, here are no other studies to back up these findings, so further measurements of these trials and establishment of new additional trials are needed to verify the results which indeed is also needed for poplars in all of Sweden.
4.2.3 Clonal differences
In Table 7 we have demonstrated differences among different material groups. However, aiming at selection of well performing clones for commercial use, this result is not so relevant. We have concluded that there are generally intermediate to large clonal differences for all traits as indicated by the H2 and CVG estimates in Table 8, which indeed can be practically used. This is illustrated in Fig. 2 where the clonal values for height growth within each material group of hybrid aspen at site Swe_A1 are presented. It is quite clear that superior clones can be found within each of the material groups. From a multi-trait evaluation over sites it is possible to optimize the selection of the very best clones for future use. This will be carried out after approximately another 4 years of growth, i.e. when close to half of the rotation time has been reached and growth data is more reliable.
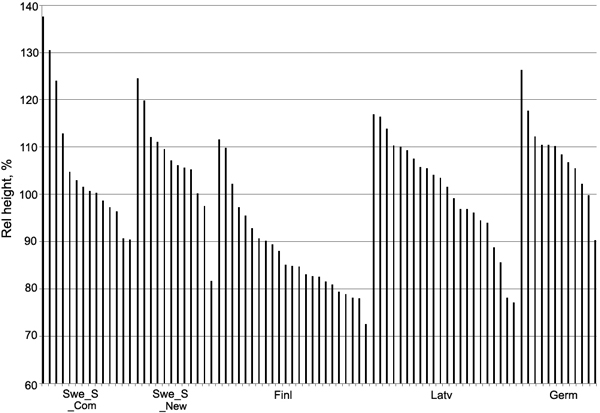
Fig. 2. Relative (%) clonal values (BLUP) for height in relation to the overall trial mean, sorted from the best to the worst clone, within each material group of hybrid aspen at site Swe_A1. Note that the y-axis starts at 60%.
5 Conclusions
The main conclusions of this study are:
• The growth performance was generally higher for hybrid aspen than poplar.
• The survival rates were generally higher for hybrid aspen than for poplar. North of approximately latitude 62 the poplar clones in the study appear to need very favorable climatic conditions to perform well.
• The poor performance of poplar compared to hybrid aspen is likely due to climatic maladaptation or to high soil acidity (pH < 4.5) commonly associated with forest land types. In this sense, hybrid aspen seems to be a more robust species.
• The genetic variation and genetic control for growth, phenology and to some extent survival, were in general intermediate or large for both species, which indicate good possibilities for effective clonal selection to improve growth performance.
• The site x site correlations for growth were for hybrid aspen mostly strong, while they were inconsistent for poplars. For hybrid aspen, the results suggest, at least on agricultural land, that the entire southern part of Sweden and the coastal areas of northern Sweden, respectively, can be treated as two separate test and deployment areas. It is too early to make any corresponding conclusions for poplar.
• The growth performance of both species needs to be confirmed at a higher age but the results in this study support current recommendations for utilization of selected hybrid aspen and poplar regeneration material in Sweden.
Acknowledgements
This study was financially supported by the Swedish Energy Agency (Project no 35136-1, Improved regeneration material for hybrid aspen and poplar) and the Nordic Energy Research (within project wood based energy systems from Nordic forests, ENERWOODS), to whom we are very grateful. We express our thanks to the support and efforts of establishing the trial by Skellefteå Kraft, Svenska Kyrkan, Södra Skogsägarna, Hargs gods, Henrik Tham and all field personnel at Skogforsk. We are also grateful to the Lounkær foundation and Prof. J. Bo Larsen for providing inspiration, initial contacts, economical support and areas for the Danish part of the study and Brian Ørnhøj Petersen for providing both agricultural sites and technical support. Finally, we extend special thanks to Palle Madsen at the University of Copenhagen and to the two reviewers, for valuable comments to the manuscript.
References
Boysen B., Strobl S. (eds.) (1991). A grower’s guide to hybrid poplar. Ministry of Natural Resource, Peterborough, Ontario, Canada. 148 p.
Brázdil R., Dobrovolný P., Luterbacher J., Moberg A., Pfister C., Wheeler D., Zorita E. (2010). European climate of the past 500 years: new challenges for historical climatology. Climatic Change 101(1–2): 7–40. https://doi.org/10.1007/s10584-009-9783-z.
Ceulemans R., Scarascia-Mugnozza G., Wiard B., Braatne J., Hinckley T., Stettler R., Isebrands J., Heilman P. (1992). Production physiology and morphology of Populus species and their hybrids grown under short rotation. Clonal comparisons of 4-year growth and phenology. Canadian Journal of Forest Research 22(12): 1937–1948. https://doi.org/10.1139/x92-253.
Christersson L. (2006). Biomass production of intensively grown poplars in the southernmost part of Sweden: observations of characters, traits and growth potential. Biomass and Bioenergy 30(6): 497–508. https://doi.org/10.1016/j.biombioe.2005.10.003.
Christersson L. (2010). Wood production potential in poplar plantations in Sweden. Biomass and Bioenergy 34(9): 1289–1299. https://doi.org/10.1016/j.biombioe.2010.03.021.
Christersson L., von Fircks H., Sihe Y. (1987). Damage to conifer seedlings by summer frost and winter drought. In: Li P.H. (ed.). Plant cold hardiness. Allan R. Liss Inc., New York, USA. p. 203–210.
Czapowskyj M., Safford L. (1993). Site preparation, fertilization, and 10-year yields of hybrid poplar on a clearcut forest site in eastern Maine, USA. New Forests 7(4): 331–344.
Dickmann D., Kuzovkina J. (2008). Poplars and willows of the world, with emphasis on silviculturally important species. FAO Forest Management Division Working Paper IPC/9-2, Rome, Italy.
Einspahr D.W., Van Buijtenen J., Peckham J. (1963). Natural variation and heritability in triploid aspen. Silvae Genetica 12(2): 51–58.
Einspahr D.W., Benson M., Peckham J. (1967). Variation and heritability of wood and growth characteristics of five-year-old Quaking Aspen. Institute of Paper Chemistry. Genetics and Physiology, Note 1, Madison W1: 1–6.
Eriksson T., Lindsjö I. (1981). The influence of pH on growth and nutrition of some energy forest tree species [Alnus incana, grey alder, Betula verrucosa, birch, Populus trichocarpa, poplar, Salix viminalis, twiggy willow]. Teknisk Rapport-Projekt Energiskogsodling.
Fahlvik N., Johansson U., Nilsson U. (2009) Skogsskötsel för ökad tillväxt. In: Larsson S., Lundmark T., Ståhl G. (eds.). Möjligheter till intensivodling av skog. [Prospects of intensive forest cultivations]. Slutrapport från regeringsuppdrag Jo 2008/1885. ISBN 978-91-86197-40-7.
GEUS (2016). Geologisk kort over de overfladenære jordarter “Jordartskort 1:200.000”. [Geological cards for upper soil types]. http://www.geus.dk/publications/grundvandsovervaagning/index.htm.
Gianola D., Norton H.W. (1981). Scaling threshold characters. Genetics 99: 357–364.
Gilmour A., Cullis B., Thompson R. (1999). Analysis of genetic models and multivariate data using ASREML. Orange: NSW Agriculture.
Hannerz M. (1998). Genetic and seasonal variation in hardiness and growth rhythm in boreal and temperate conifers – a review and annotated bibliography. Report no. 2. SkogForsk, Uppsala. 140 p.
Heräjärvi H., Junkkonen R. (2006). Wood density and growth rate of European and hybrid aspen in southern Finland. Baltic Forestry 12(1): 2–8.
Hjelm K., Rytter L. (2016). The influence of soil conditions, with focus on soil acidity, on the establishment of poplar (Populus spp.) New Forests 47(5): 731–750. https://doi.org/10.1007/s11056-016-9541-9.
Howe G., Saruu P., Davis J., Chen T. (2000). Quantitative genetics of bud phenology, frost damage and winter survival in an F2 family of hybrid poplars. Theoretical and Applied Genetics 101(4): 632–642. https://doi.org/10.1007/s001220051525.
Hytönen J., Jylhä P. (2008). Fifteen-year response of weed control intensity and seedling type on Norway spruce survival and growth on arable land. Silva Fennica 42(3): 355–368. https://doi.org/10.14214/sf.242.
Ilstedt B. (1996). Genetics and performance of Belgian poplar clones tested in Sweden. Forest Genetics 3(4): 183–195.
Jönsson A.M., Bärring L. (2011). Ensemble analysis of frost damage on vegetation caused by spring backlashes in a warmer Europe. Natural Hazards and Earth System Science 11(2): 401–418. https://doi.org/10.5194/nhess-11-401-2011.
Karltun E. (1996). Markkemiska analyser inom ståndortskarteringen – metodbeskrivningar. [Soil chemical analyses of site mapping. Method description]. Ståndortskarteringen, Rapport. SLU, Umeå.
Langvall O. (2011). Impact of climate change, seedling type and provenance on the risk of damage to Norway spruce (Picea abies (L.) Karst.) seedlings in Sweden due to early summer frosts. Scandinavian Journal of Forest Research 26(S11): 56–63. https://doi.org/10.1080/02827581.2011.564399.
Nielsen U.B., Madsen P., Hansen J.K., Nord-Larsen T., Nielsen A.T. (2014). Production potential of 36 poplar clones grown at medium length rotation in Denmark. Biomass and Bioenergy 64: 99–109. https://doi.org/10.1016/j.biombioe.2014.03.030.
Nordiska Ministerrådet (2009). Hållbar utveckling – en ny kurs för Norden. [Sustainable development – a new course for the Nordic countries]. Nordiska Ministerrådet, Copenhagen, Denmark. http://norden.diva-portal.org/smash/get/diva2:701331/FULLTEXT01.pdf.
Perttu K., Morén A. (1995) Regionala klimatindex – verktyg vid bestämning av skogsproduktion. [Regional climate index – tools for decision of forest yield]. Fakta Skog, nr. 13. SLU, Umeå.
Pliura A., Suchockas V., Sarsekova D., Gudynaitė V. (2014). Genotypic variation and heritability of growth and adaptive traits, and adaptation of young poplar hybrids at northern margins of natural distribution of Populus nigra in Europe. Biomass and Bioenergy 70: 513–529. https://doi.org/10.1016/j.biombioe.2014.09.011.
Rosvall O., Jansson G., Andersson B., Ericsson T., Karlsson B., Sonesson J., Stener L.-G. (2001). Genetiska vinster i nuvarande och framtida fröplantager och klonblandningar. [Genetic gain from present and future seed orchards and clone mixes]. Redogörelse Vol. 1. Skogforsk, Uppsala. 41 p.
Rytter L., Ingerslev M., Kilpeläinen A., Torssonen P., Lazdina D., Löf M., Madsen P., Muiste P., Stener L.-G. (2016). Increased forest biomass production in the Nordic and Baltic countries – a review on current and future opportunities. Silva Fennica 50(5) article 1660. https://doi.org/10.14214/sf.1660.
Rytter L., Johansson T., Karačić A., Weih M. (2011). Orienterande studie om ett svenskt forskningsprogram för poppel. [Investigation of a Swedish research program on the genus Populus]. SkogForsk, Uppsala, Sweden.
Rytter L., Stener L.-G. (2014). Growth and thinning effects during a rotation period of hybrid aspen in southern Sweden. Scandinavian Journal of Forest Research 29(8): 747–756. https://doi.org/10.1080/02827581.2014.968202.
Rytter L., Dimitriou I., Engqvist G., Hjerpe K., Nilsson B., Ramstedt M. (2016). Country report 2012–2015. Report for International Poplar Commission. Nation Poplar Commission of Sweden. 12 p. http://www.fao.org/forestry/ipc2016/91148/en/.
SAS Institute Inc. (2011). SAS/STAT User’s guide 9.3. SAS Institute, Inc. Cary, NC.
Stener L.-G. (2010). Tillväxt, vitalitet och densitet för kloner av hybridasp och poppel i sydsvenska fältförsök. [Growth, vitality and density for clones of hybrid aspen and poplars in south Swedish field trials]. Arbetsrapport nr. 717. Skogforsk, Uppsala.
Stener L.-G., Karlsson B. (2004). Improvement of Populus tremula × P. tremuloides by phenotypic selection and clonal testing. Forest Genetics 11(1): 13–24.
Tullus A., Rytter L., Tullus T., Weih M., Tullus H. (2012). Short-rotation forestry with hybrid aspen (Populus tremula L.× P. tremuloides Michx.) in Northern Europe. Scandinavian Journal of Forest Research 27(1): 10–29. https://doi.org/10.1080/02827581.2011.628949.
Yu Q. (2001). Selection and propagation of hybrid aspen clones for growth and fibre quality. Dissertation for Ph.D. Department of Applied Biology, University of Helsinki, Finland. 41 p.
Yu Q., Pulkkinen P. (2003). Genotype–environment interaction and stability in growth of aspen hybrid clones. Forest Ecology and Management 173(1): 25–35. https://doi.org/10.1016/S0378-1127(01)00819-2.
Total of 39 references.
