Sergio Rossi  ,
Hubert Morin,
François Gionest,
Danielle Laprise
,
Hubert Morin,
François Gionest,
Danielle Laprise
Spatially explicit structure of natural stands dominated by black spruce
Rossi S., Morin H., Gionest F., Laprise D. (2013). Spatially explicit structure of natural stands dominated by black spruce. Silva Fennica vol. 47 no. 3 article id 973. https://doi.org/10.14214/sf.973
Abstract
Black spruce [Picea mariana (Mill.) BSP] regeneration emerges in clusters near the pre-existing boles within a few years after the passage of fire. This paper tested the hypothesis that black spruce forests still maintain the spatial structure deriving from postfire stand initiation. Trees and saplings were monitored during 2000-2007 and the horizontal and vertical structure of the stands were investigated on four permanent plots in the boreal forest of Quebec, Canada. Plots showed 1300-2150 trees ha-1, and were composed of trees with homogeneous sizes and a very small proportion of saplings. These characteristics identify single cohorts generated by complete, or almost-complete, stand replacement by fire. Ripley’s L(r) functions showed that the spatial pattern of trees and saplings ranged from random to aggregated, thus demonstrating that the clustering distribution of the individuals in black spruce forests can be maintained even after 80-120 years from stand initiation. These findings could results from incomplete self-thinning or from an environment with heterogeneous distribution of resources. The practices of ecosystem management recently developed in Eastern Canada should take into account both the horizontal and vertical structure to better modulate the competition among individuals during partial harvesting.
Keywords
boreal forest;
Picea mariana;
mortality;
recruitment;
stand dynamics
-
Rossi,
Département des Sciences Fondamentales, Université du Québec à Chicoutimi, 555 boulevard de l’Université, Chicoutimi (QC), G7H2B1, Canada
E-mail
sergio.rossi@uqac.ca
- Morin, Département des Sciences Fondamentales, Université du Québec à Chicoutimi, 555 boulevard de l’Université, Chicoutimi (QC), G7H2B1, Canada E-mail hubert_morin@uqac.ca
- Gionest, Département des Sciences Fondamentales, Université du Québec à Chicoutimi, 555 boulevard de l’Université, Chicoutimi (QC), G7H2B1, Canada E-mail francois_gionest@uqac.ca
- Laprise, Département des Sciences Fondamentales, Université du Québec à Chicoutimi, 555 boulevard de l’Université, Chicoutimi (QC), G7H2B1, Canada E-mail danielle_laprise@uqac.ca
Received 1 March 2013 Accepted 17 May 2013 Published 12 September 2013
Views 151690
Available at https://doi.org/10.14214/sf.973 | Download PDF

1 Introduction
In Eastern Canada, the silvicultural treatments are the subject of a heated debate. The rationale of ecosystem management intends to substitute the extensive forestry practices of even aged (or low-retention) cutting such as clearcutting and careful logging in order to maintain functioning of the boreal forests over time (Bergeron et al. 2002; Gauthier et al 2009). Partial-cutting is in part substituting the prevalent practices to reduce disturbances and more closely simulate the natural processes occurring in boreal environments. These new practices attempt to artificially maintain or recreate the most appropriate structure of the stands. However, as for conservation or restoration, management also inevitably requires detailed knowledge on ecology and growth dynamics of the tree species in natural stands, particularly when the aim is to recreate a structural condition as natural as possible (Kneeshaw and Gauthier 2003). At stand level, composition, and vertical (e.g. distribution in age and size) and horizontal (spatial distribution of the stems) structures are the key component of a forest, the factors that generate the natural habitats for organisms and maintain biodiversity. Nevertheless, the recent silvicultural approaches for Eastern Canada place most emphasis on composition and vertical structure, while scarce information is generally provided on the horizontal structure of stands (Gauthier et al. 2009).
Fire is one of the most important processes of forest regeneration, and native species of the boreal forest are well adapted to this recurrent ecological factor (Rowe 1983). The disturbance kills a part of or all the previously established vegetation, and the rapid and effective postfire recruitment closely relies on the sprouting ability or seed bank strategy of tree species (Wang and Kemball 2005). Black spruce [Picea mariana (Mill.) BSP] grows in a broad transcontinental band from Alaska, USA, to Newfoundland, Canada, forming extensive closed forests in north-eastern North America. This species provides for prolonged seed storage by maintaining aerial seed banks in semi-serotinous cones. Following heating, the cones open and release the seeds that create, within a few years, a new, even-aged stand (Charron and Greene 2002; Greene and Johnson 1999). Because of its high flammability, the reproduction strategy of black spruce is an adaptation for its dominance in ecosystems where fires are chronic, and stands burn at fire return intervals of 150 years (Le Goff and Sirois 2004).
In stands dominated by black-spruce, the early establishment of postfire recruits show a distinctive spatial pattern that is related to seedbed conditions. A thick organic layer or massive presence of postfire duff or ericaceous shrubs are frequently lethal environments for seedlings, because the small seeds of black spruce produce small and slow-growing germinants that are unable to deepen their short root system as far as the mineral soil, and exhibit low competitive potential against the vegetative recruitment of perennial plants (Greene and Johnson 1999). On the contrary, mineral soils with thin burnt humus or organic layers of a depth less than the hypocotyl length have shown greater seedling survival (Greene and Johnson 1999; Hesketh et al. 2009). Microsites with concave profiles or mosses are colonized more frequently by the seedlings, demonstrating that moisture is crucial for survival of black spruce regeneration (Filion and Morin 1996; Kemball et al. 2006; Wirth et al. 2008).
Fire burns the organic matter and consumes about 50% of the organic layer, although the reduction is very heterogeneous, with reductions in depth ranging between 30 and 70% after prescribed fires (Greene et al. 2007). After fire, the thinner organic layers tend to be clustered around the bases of tree boles, where soil moisture is lower, and heat by smoldering is retained better, thus burning longer and consuming the deeper organic matter (Greene et al. 2007; Miyanishi and Johnson 2002). This mosaic of organic layer reduction is typical and frequently observed (Johnstone and Kasischke 2005; Miyanishi and Johnson 2002), and results in a heterogeneous spatial pattern of seedling recruitment within the stand, with successful germinations mainly occurring near the burnt tree boles. As a consequence, it has been observed that black spruce regenerates within three years after fire in clusters near the pre-existing boles (Filion and Morin 1996; St-Pierre et al. 1991).
In natural forests (i.e. forests where the absence of human impact assures that trees have developed under the influence of natural drivers only), stand structure is the result of a number of factors and processes involving the strategy of plant reproduction, the mechanisms of seedling establishment, and the competition for resources occurring within and among species. Thus, the young stands evolve according to the interactions that individuals experience all throughout its life with their neighborhoods. In black spruce, the particular spatial pattern of early postfire recruitment produces local densities of potential competitors, and can logically determine a heterogeneous exploitation and local depletion of resources (Barot et al. 1999). Since individuals with different neighborhoods have different dynamics of growth and survival, the spatial patterns of trees, the competition among neighborhoods, and the environmental conditions heavily influence the evolution of a stand. Does the early clustering pattern of postfire black spruce is maintained in adult stands? The aim of this study was to answer this question by assessing the spatial arrangement of the individuals in mature black spruce stands by means of (i) an eight-years-long monitoring of trees and saplings and (ii) an investigation of the horizontal and vertical structure of the stand on four permanent plots in the boreal forest of Quebec, Canada. A hypothesis was proposed and tested that the mature forest still maintains the spatial features deriving from stand initiation, with trees grouped in clusters according to the pattern of postfire recruitment of regeneration. The spatial patterns of the individuals provide valuable information about how adult trees exploit resources, interact with neighborhoods and tolerate the competition. In turn, this knowledge can be used in implementing the new strategies of forest management and tree selection during partial cutting of black spruce forests.
2 Material and methods
2.1 Study area and stand selection
The study was conducted in the Saguenay-Lac-Saint-Jean area, in the boreal forest of Quebec, Canada. The region has a gently rolling topography with hills reaching 500–700 m a.s.l. on thick and undifferentiated glacial till deposits. Four sites [Simoncouche (abbreviated as SIM), Bernatchez (BER), Mistassibi (MIS) and Camp Daniel (DAN)] were identified in mature even-aged black spruce stands that lacked in evidences of human impact and could be considered as natural forests (Table 1). The region has a typical boreal climate with cold winters and cool summers. According to data collected during the period 2002–2010 by weather stations installed in each stand, the mean annual temperature varied between –0.9 and 2.0 °C, while May-September temperatures are 11.0–13.3 °C (Table 1). The sites are characterized by long winters with the coldest temperature reaching –47.1 °C in DAN. The summers are short, with extreme temperatures exceeding 30 °C in all sites. More details on site characteristics were provided by Boulouf Lugo et al. (2012).
| Table 1. Location and climatic characteristics of the four black spruce stands listed at increasing latitudes of the boreal forest of Quebec, Canada. Climatic data are based on weather measurements covering the period 2002–2010. | |||||||
| Site | Latitude | Longitude | Altitude (m a.s.l.) | Mean temperature | Extreme annual temperature | ||
| Annual (°C) | May-September (°C) | Maximum (°C) | Minimum (°C) | ||||
| SIM | 48°13´N | 71°15´W | 338 | 2.0 | 13.3 | 35.7 | –39.7 |
| BER | 48°51´N | 70°20´W | 611 | 0.3 | 11.4 | 33.1 | –39.8 |
| MIS | 49°43´N | 71°56´W | 342 | 1.0 | 12.7 | 35.1 | –42.4 |
| DAN | 50°41´N | 72°11´W | 487 | –0.9 | 11.0 | 34.2 | –47.1 |
2.2 Plot establishment
Permanent plots of 20 m × 20 m with a buffer zone of 3 m were chosen and delimited by an optical prism to annually monitor tree mortality and establishment from 2000 to 2007, by collecting data on all individuals taller than 3 m (Fig. 1). The living individuals were identified using permanent tags with unique alphanumeric codes. Twenty-five fixed points were established within the plot on a grid at intervals of 5 m and the location of every individual was mapped by measuring the polar distances of the stems from the predefined points to the nearest 1 cm (Fig. 1). For data analysis and mapping, every pair of x–y coordinates was calculated relative to the south-western corner of the study plot.
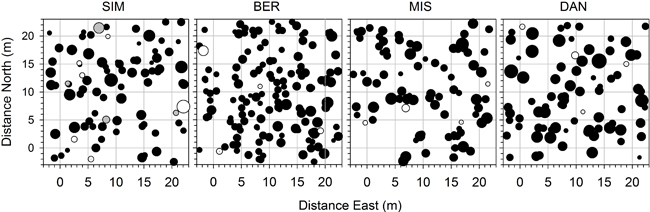
Fig. 1. Locations of the trees at plot establishment in 2000 in the four permanent plots in the boreal forest of Quebec, Canada. Black spruce, balsam fir and white birch are indicated with black, white and grey circles, respectively. Symbol size is proportional to DBH. In each plot, 25 fixed points were assessed at the nodes of the grid represented by the grey lines. Only the individuals located within the plot of 20×20 m were used to assess the stand characteristics.
2.3 Stand structure
In each stand, the specimens with diameter at breast height (dbh) of less and more than 9 cm were considered as saplings and trees, respectively, according to the definition supplied for the Quebec by the Ministère des Ressources Naturelles, de la Faune et des Parcs (2003). Individuals were measured by assessing height and dbh. Each year, the tagged trees were checked for mortality, dbh were re-measured, and any new individuals taller than 3 m found on the plots were incorporated into the dataset. The tagged trees that were found dead in a successive monitoring were not re-measured. Dominant height was defined as the mean height of the 100 trees with larger dbh per hectare (Pardé and Bouchon 1988), which in this work corresponded to the four largest trees per plot.
Trees from a second circular plot of 20 m in diameter located in the same stands than the study sites were harvested at the beginning of the study and two-cm-thick discs were collected at the stem base. Discs were air-dried and sanded with progressively finer grade sandpaper and tree-ring widths were measured to the nearest 0.01 mm using a Henson measuring system along two opposite radii. All ring width series were corrected by cross-dating performed both visually and using the COFECHA computer program to estimate the year of germination of the plants and their age (Holmes 1983).
2.4 Spatial analyses
The spatial patterns of tree and sapling distribution were quantified using Ripley’s K-statistics in order to assess which patterns deviated from random (Ripley 1977). Univariate and bivariate K-statistics were used to investigate the interactions among individuals and between trees and saplings, respectively. In a plot with n stems, the calculation of the K-statistic for a given radius r was based on counting all pairs of points separated by a distance less than r and calculating:

with

where A was the area of the plot, dij was the distance between the points i and j (Fortin and Dale 2005). Since the amplitude of the buffer zone was not sufficiently wide for edge correction, the term wij was used as weight for a geometrical correction of the edge effects, which corresponded to the inverse of the proportion of the circle centered on i and passing through j (Ripley 1977; Zaninetti 2005).
The K-statistics were estimated on all species at intervals of 0.5 m on distances ranging between 0.5 and 10 m, corresponding to half of the plot length, and were then transformed into linear form with stable variance by calculating ![]() (Fortin and Dale 2005). Plotting L(r) as a function of r, close-to-zero values were expected under a Poisson (i.e. spatially random) process. Departures from a random distribution showed negative or positive values, indicating aggregation (clustering) or overdispersion (segregation), respectively. A Monte Carlo approach was used by repetitively estimating the spatial statistics in simulated plots generated 1000 times with random coordinates of the same number of stems (Fortin and Dale 2005). With the distributions of the L(r) calculated by the simulations, the 95% higher and lower confidence limits were extracted to construct the 95% Monte Carlo envelope for testing the significance of the results. Values greater or lower than this envelope were considered to indicate a pattern significantly different from randomness. All analyses were performed using routines specifically written in SAS 9.2 (SAS Institute Inc., Cary, NC).
(Fortin and Dale 2005). Plotting L(r) as a function of r, close-to-zero values were expected under a Poisson (i.e. spatially random) process. Departures from a random distribution showed negative or positive values, indicating aggregation (clustering) or overdispersion (segregation), respectively. A Monte Carlo approach was used by repetitively estimating the spatial statistics in simulated plots generated 1000 times with random coordinates of the same number of stems (Fortin and Dale 2005). With the distributions of the L(r) calculated by the simulations, the 95% higher and lower confidence limits were extracted to construct the 95% Monte Carlo envelope for testing the significance of the results. Values greater or lower than this envelope were considered to indicate a pattern significantly different from randomness. All analyses were performed using routines specifically written in SAS 9.2 (SAS Institute Inc., Cary, NC).
3 Results
3.1 Stand description
A total of 251 trees (individuals taller than 3 m and with dbh > 9 cm) were identified and mapped in the four stands in 2000 (Table 2). At their establishment, plots contained between 52 and 86 trees, which indicated a variability in density from 1300 to 2150 trees ha–1, with the densest plot being BER. The dominant height varied between 18.0 and 20.2 m, gradually decreasing with latitude. Mean dbh was 15.8–19.4 cm, which produced a basal area ranging between 30.1 and 43.5 m2 ha–1. Black spruce was the dominant species in all plots, where it represented 88–97% of trees. Balsam fir [Abies balsamea (L.) P. Mill.] and white birch (Betula papyrifera Marsh.) were also present but only accounted for a modest percentage of the total basal area of the stand (Table 2).
Of the 37 saplings (individuals taller than 3 m and with dbh < 9 cm) mapped in 2000, 65% were black spruce while only 30% were balsam fir (Table 3). White birch was observed only in SIM with 2 saplings. Height and dbh of the saplings were similar between plots, except for MIS where they were smaller. Density varied between 125 and 350 saplings ha–1. Maximum and minimum densities were observed in SIM and BER, respectively (Table 3).
| Table 2. Stand characteristics measured at plot establishment in 2000. | |||||||
| Stand | Number of trees in the plot | Dominant height (m) | Mean DBH (cm) | Stand density (trees ha–1) | Stand basal area (m2 ha–1) | ||
| Black spruce | Balsam fir | White birch | |||||
| SIM | 46 | 4 | 2 | 20.2 | 16.7 | 1300 | 30.1 |
| BER | 84 | 2 | 0 | 20.0 | 15.8 | 2150 | 43.5 |
| MIS | 56 | 2 | 0 | 19.2 | 17.3 | 1450 | 36.2 |
| DAN | 52 | 3 | 0 | 18.0 | 19.4 | 1375 | 43.4 |
| Table 3. Characteristics of the saplings mapped at plot establishment in 2000. | ||||||
| Stand | Number of saplings in the plot | Height (m) | Mean DBH (cm) | Density (saplings ha–1) | ||
| Black spruce | Balsam fir | White birch | ||||
| SIM | 8 | 4 | 2 | 5.9 | 6.6 | 350 |
| BER | 4 | 1 | 0 | 6.8 | 6.8 | 125 |
| MIS | 9 | 2 | 0 | 5.0 | 5.9 | 275 |
| DAN | 3 | 4 | 0 | 5.9 | 6.4 | 175 |
BER, MIS and DAN clearly showed an even-aged structure, with individuals having an average age of 109–132 years (Fig. 2). In these stands, age distribution was narrow and all values were concentrated within 40 years. SIM was the youngest stand with an average age of 96 years, and exhibited a more irregular and wider age distribution, with 80% of trees exhibiting an age of 60–110 years and a bimodal distribution of germination. Moreover, 12% of trees were older than 160 years, which indicated the occurrence of an incomplete stand replacement disturbance (Fig. 2).
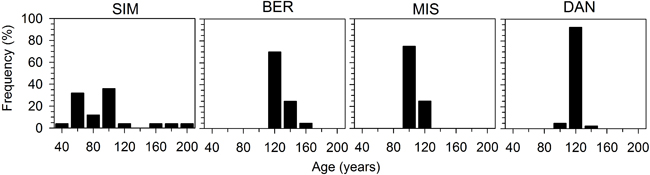
Fig. 2. Frequency distribution of age measured at plot establishment in the four black spruce stands in the boreal forest of Quebec, Canada. Ages represent individuals harvested in sample plots located close to the four permanent plots.
3.2 Stand demography
In general, all stands were composed of trees with homogeneous sizes and a very small proportion of saplings (Figs. 1 and 3). At plot establishment, the distribution of dbh was bell-shaped, except for DAN that showed a more even distribution and wider variation in dbh covering a range of 27.7 cm. BER had the narrower distribution, with 29.3% of individuals included in the dbh class of 16 cm (Fig. 3). During the monitoring period (2000–2007), the number of trees declined gradually in all stands (Fig. 4). The higher tree mortality was observed in SIM and BER, with dead individuals representing 14.8 and 6.8% of the tree population assessed in 2000, respectively, while DAN showed only 1.8% of mortality in eight years. Tree recruitment was observed in SIM, with two saplings classified as trees in 2005, while there was a lack of recruitment in the other stands during the whole monitoring period. Between 2000 and 2007, the number of saplings clearly decreased in SIM and MIS, while in DAN no saplings either died or were recruited (Fig. 4). Over two years, from 2001 to 2003, saplings in SIM decreased from 350 to 225 individuals ha–1. Sapling recruitment was rare and observed in SIM and BER with only one individual per plot recruited during 2000–2007.
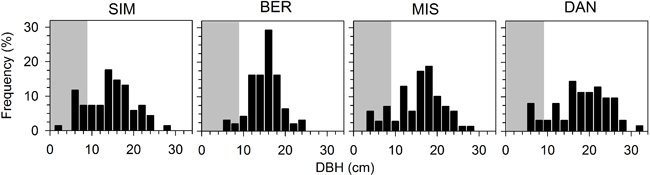
Fig. 3. Frequency distribution of diameter at breast height (DBH) in the four black spruce stands in the boreal forest of Quebec, Canada. Grey background indicates the individuals considered as saplings.
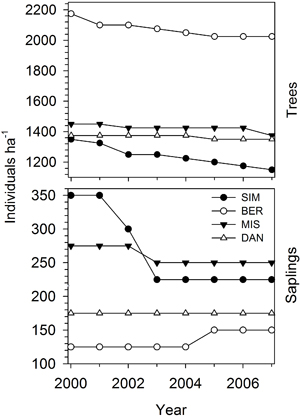
Fig. 4. Variation in the number of trees and saplings during the period 2000–2007 in the four permanent plots of black spruce in the boreal forest of Quebec, Canada.
3.3 Spatial analyses
Ripley’s L(r) functions generally revealed similar patterns between the eight monitoring years, so only results from 2000, at plot establishment, and 2007, at the end of the monitoring period, were reported (Figs. 5 and 6). In all stands, the spatial distributions of trees showed L(r) functions frequently encompassed within the 95% confidence envelopes, which indicated a random pattern of stem distribution in the plot. BER and MIS showed departures from randomness with significant aggregation of trees at distances greater than 5 m and between 1 and 3 m, respectively. SIM showed significant aggregations between 3 and 8 m in 2007, but not in 2000. The pattern of saplings was generally random at all spatial scales, with aggregation and significant L(r) functions estimated in BER at distances of 1–3 m in both 2000 and 2007 (Figs. 5 and 6). Wide confidence envelopes were observed in BER and DAN in 2000 and 2007, and in SIM in 2007 due to the small number of saplings detected in the plots. When considering all individuals together, results were similar to those observed for trees, with random patterns except in BER and MIS, which exhibited significant aggregations in both years. In SIM, the significant aggregations of trees and saplings appeared only in 2007. No significant overdispersion (segregation) was observed for either trees or saplings, except in MIS at 5–6 m. Overall, random distributions were detected also by bivariate L(r) functions in the spatial pattern of trees vs. saplings, which were consistent between the two years (Figs. 5 and 6). However, trees were occasionally segregated from saplings at 1 m in SIM and aggregated at 4 m in DAN.
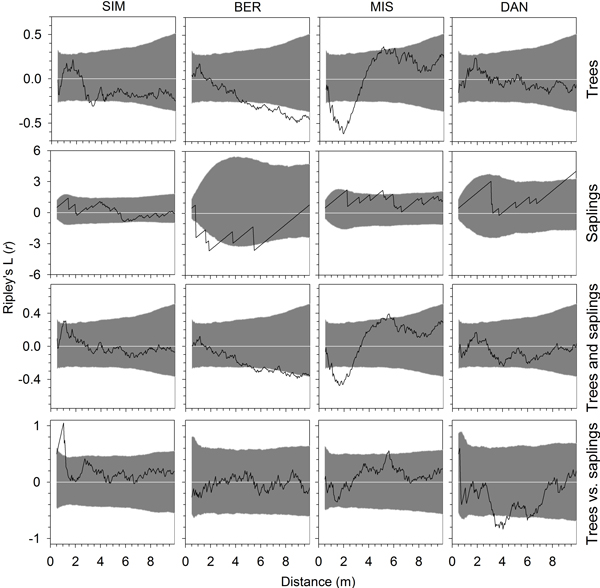
Fig. 5. Univariate and bivariate Ripley’s L(r) functions calculated for the year 2000, at plot establishment, for trees, saplings, all individuals and for trees versus saplings in the four permanent plots of black spruce in the boreal forest of Quebec, Canada. Grey areas represent the 95% confidence envelopes based on Monte Carlo simulations. Values of the functions greater or lower than this envelope indicate a pattern significantly different from randomness and a tendency to overdispersion (segregation) or aggregation (clustering) of individuals, respectively.
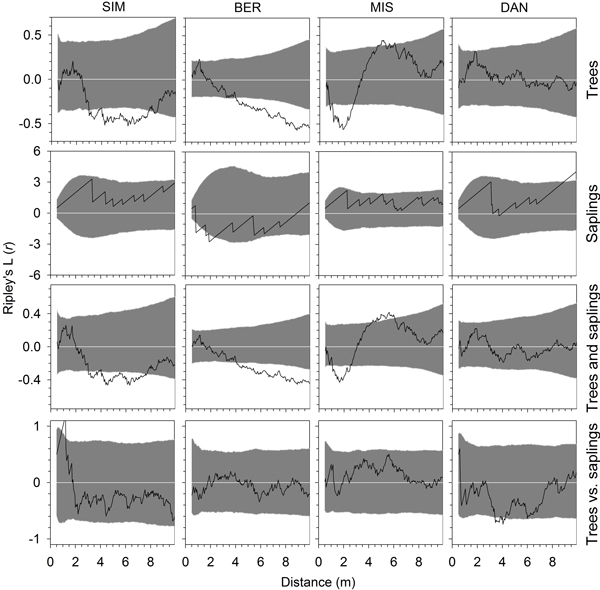
Fig. 6. Univariate and bivariate Ripley’s L(r) functions calculated for the year 2007, at the end of the monitoring period, for trees, saplings, all individuals and for trees versus saplings in the four permanent plots of black spruce in the boreal forest of Quebec, Canada. Grey areas represent the 95% confidence envelopes based on Monte Carlo simulations. Values of the functions greater or lower than this envelope indicate a pattern significantly different from randomness and a tendency to overdispersion (segregation) or aggregation (clustering) of individuals, respectively.
4 Discussion
4.1 Horizontal structure
In black spruce dominated forests, a number of factors such as the anatomical characteristics of seeds, ecological requirements of seedlings, and spatial heterogeneity of seedbed conditions generates clusters of recruits after fire that are located near the pre-existing boles. The present study monitored trees and saplings during the period 2000–2007 and investigated the spatial pattern of the stems on four permanent plots in the boreal forest of Quebec, Canada to verify whether the clusters are also maintained during the adult phase of stand development. The distribution of stems exhibited either random and aggregated patterns, with significant aggregations observed in BER and MIS over the eight monitoring years, and in SIM at the end of the monitoring period. Thus, the initial hypothesis that trees of mature stands still maintain the spatial distributions deriving from postfire recruitment of regeneration was only partially supported in the four sampled stands. These results confirmed the observations by Horton and Groot (1987) and St-Pierre et al. (1991), reporting that spatial distributions of trees ranged from random to aggregated in mature black spruce sites.
Black spruce regenerates within three years, with a peak in the first summer, and recruitment declining to nearly zero by the fourth year after fire (Charron and Greene 2002). This massive regeneration leads to a density-independent mortality ranging between 85 and 98% of individuals during the first year of seedling recruitment. Early mortality is mostly associated with environmental and biotic stresses (Lussier et al. 2002). In the decades following seedling emergence, boreal species still exhibit mortalities of about 10–20% (unpublished data), which is proportional to stand density and related to competition among individuals. Although this mortality may occur only during the juvenile stages (Kenkel 1988), stem exclusion and self-thinning were reported for a variety of tree size in both young and old forests (Carleton et al. 1987; Harper et al. 2005). The reduction in trees and saplings observed in our plots confirmed that substantial rates of mortality can extend until the mature phases of stand development. The heterogeneous growth of trees causes asymmetric competition for resources, with the taller and larger individuals having more access to light and nutrients, and an accentuated mortality of individuals in the smallest size classes (Weiner and Thomas 1986). Once an even-aged forest is established, the self-thinning caused by resource competition generates systematic changes in spatial patterns occurring over time, from an aggregated pattern to a random pattern at intermediate stages of stand development (Kenkel et al. 1997). However, the residual aggregation of trees observed in the two denser plots over the monitoring period and in SIM in 2007 could indicate either an incomplete process of self-thinning or a heterogeneous environment with nutrient-rich patches of soil, where interaction and competition among trees could be altered by the more favourable conditions and the consequent mortality reduced (Barot et al. 1999). Our results clearly demonstrated that black spruce forests can still maintain aggregated arrangements of the individuals after 80–120 years from stand initiation.
It is well known that larger samples furnish more statistically-reliable results. Similarly, also Ripley’s functions are sensitive to plot size, and spatial analyses in forest sciences are more suitably employed on plots larger than 0.1 ha (Larson and Churchill 2012). At the latitudes where the study was carried out, the boreal forest generates homogeneous stands dominated by small trees with short branches and high density, reaching 3750 trees ha–1 (Rossi et al. 2009a). In this investigation, data were repetitively collected for eight years from the living individuals growing in remote pristine forests, thus the assessment of plot size obviously resulted in a trade-off between the extent of sampling, spatial accuracy, and resource employment. However, sample size was smaller than that generally adopted for spatial analyses, and individuals were identified as trees and saplings according to the definition supplied for the Quebec by the Ministère des Ressources Naturelles, de la Faune et des Parcs (2003), with a minimum threshold of 3 m in height. This inevitably requires a honest balance of considerations and limitations about the general reliability of the results of the horizontal structure. Care should be taken principally in interpreting the results of saplings, which are sporadic in black spruce forests, with an observed density of between 125 and 350 individuals ha–1 in the study plots.
4.2 Vertical structure
Stand characteristics are species-specific adaptations to natural disturbances and obviously reflect the strategies of reproduction and survival of the boreal species. Consequently, in similar environmental conditions, stands dominated by different tree species can show divergent vertical structures. In the present study, the density of saplings was observed to range between 125 and 350 individuals ha–1. Although no information was collected on their origin, it is assumed that most saplings were produced by layering, which occurs when the lower living branches become embedded in the surface organic material and develop roots (Paquin and Doucet 1992; Rossi et al. 2009a). At the same density and age, balsam fir, which also forms monospecific stands in the studied region and was detected, although only with some rare specimens, in the studied plots, creates very different stand structure profiles by forming a dense understory with up to 1750 saplings ha–1 (Rossi and Morin 2011). Balsam fir establishes its survival strategy by maintaining a dense seedling bank to replace dead trees after insect outbreaks, which generally leave small plants in the understory less damaged (Bergeron et al. 1995; Kneeshaw and Bergeron 1999). In contrast, the scarcity of advance regeneration under black spruce trees and their extraordinary ability to disperse seeds after fire are closely related to this chronic ecological factor that primarily drives the dynamics of mortality and stand replacement of this species (Le Goff and Sirois 2004; Wang and Kemball 2005).
Three stands clearly showed a single cohort and distributions of diameters with the typical bell-shaped pattern representing trees established in an open environment after a stand replacing disturbance (Rossi et al. 2009b, 2010). The age of these stands, approximately 100–130 years, falls within the longevity range of the species, which was estimated as varying between 100 and 200 years, and corresponds to the timings of overstory breakup and initiation of sapling recruitment (De Grandpré et al. 2000; Harper et al. 2003). Accordingly, the transition towards an uneven-aged structure should occur mainly between 120 and 200 years after stand initiation (Rossi et al. 2009a, Tremblay et al. 2011). Consequently, in the absence of another intense disturbance that could again kill the whole stand, the advance regeneration constituted by the saplings is likely to form the next recruitment for gap filling in the case of smaller-scale, secondary disturbances such as dying or falling of the larger and older trees. In fact, because of its hardiness and despite its low density, advance growth of black spruce remains vigorous even after prolonged periods of suppression and is able to respond rapidly and successfully to canopy opening (Rossi et al. 2009a, Tremblay et al. 2011).
The irregular age distribution of SIM, including trees from almost every class, showed the presence of a bimodal structure. The stand had two clear cohorts, the older one probably deriving from an incomplete stand replacement after fire and being constituted by trees surviving the previous disturbance. However, the ages reported for BER, MIS and DAN also revealed wide intervals of variation ranging between 20 and 40 years that apparently contrast with the period reported for black spruce to regenerate after fire, which is three years (St-Pierre and Gagnon 1992). There is evidence that the true root collar of this species is habitually located below ground level following the recurrent growth of adventitious roots up the stem and buildup of the forest floor over time (DesRochers and Gagnon 1997). Thus, despite the accuracy of cross-dating, substantial divergences between the real and measured year of germination should be expected, and inevitably accepted, when assessing black spruce age using conventional dendrochronological analyses on the current stem base.
4.3 Implications for management
In Eastern Canada, the recent ecosystem management is attempting to modify the extensively-used forestry by using practices with minor environmental impacts and closely adapted to the various regional differences. This new approach to silviculture is mainly based on the best available ecological knowledge to establish the most suitable harvesting strategies and targets (Gauthier et al. 2009). Partial cuttings simulating disturbances of moderate severity have been proposed to open the canopy and create variability and complexity, although most implementations ignore interventions on the horizontal structure of stands. In this paper, the spatial patterns of stems were investigated in permanent plots of stands that had developed under the influence of natural disturbances only in order to provide information about how individuals are spatially arranged in a natural stand. The temporal and spatial characteristics of the analysis that concerned eight years of measures in remote regions of the boreal forest, allowed only a limited number of small plots to be monitored, thus the results of the horizontal structure should wait further confirmations for generalization. The findings of this investigation showed that mature black spruce exhibited individuals with spatial distributions from aggregated to random, while overdispersion or regular distributions were lacking. Vertical structure was simplified, and constituted by just one dominant cohort, with rare saplings in the understory. To adequately imitate nature in controlling the natural competition for resources among individuals, tree selection in mature stands should aim to maintain such spatial structures during partial harvesting.
References
Barot S., Gignoux J., Menaut J.-C. (1999). Demography of a savanna palm tree: predictions from comprehensive spatial pattern analyses. Ecology 80: 1987–2005. http://dx.doi.org/10.1890/0012-9658(1999)080[1987:DOASPT]2.0.CO;2.
Bergeron Y., Leduc A., Morin H., Joyal C. (1995). Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Canadian Journal of Forest Research 25: 1375–1384. http://dx.doi.org/10.1139/x95-150.
Bergeron Y., Leduc A., Harvey B.D., Gauthier S. (2002). Natural fire regime: a guide for sustainable management of the Canadian boreal forest. Silva Fennica 36: 81–95.
Boulouf Lugo J., Deslauriers A., Rossi S. (2012). Duration of xylogenesis in black spruce lengthened between 1950 and 2010. Annals of Botany 110: 1099–1108. http://dx.doi.org/10.1093/aob/mcs175.
Carleton T.J., Wannamaker B.A. (1987). Mortality and self-thinning in postfire black spruce. Annals of Botany 59: 621–628.
Charron I., Greene D.F. (2002). Post-wildfire seedbeds and tree establishment in the southern mixedwood boreal forest. Canadian Journal of Forest Research 32: 1607–1615. http://dx.doi.org/10.1139/x02-085.
De Grandpré L., Morissette J., Gauthier S. (2000). Long-term post-fire change in the northeastern boreal forest of Quebec. Journal of Vegetation Science 11: 791–800. http://dx.doi.org/10.2307/3236549.
DesRochers A., Gagnon R. (1997). Is ring count at ground level a good estimation of black spruce age? Canadian Journal of Forest Research 27: 1263–1267. http://dx.doi.org/10.1139/x97-086.
Filion J., Morin H. (1996). Distribution spatiale de la régéneration de l’épinette noire 8 ans après un feu en forêt boréale (Québec). Canadian Journal of Forest Research 26: 601–610. http://dx.doi.org/10.1139/x26-069.
Fortin M.-J., Dale M. (2005). Spatial analysis: a guide for ecologists. Cambridge University Press, Cambridge, UK.
Gauthier S., Vaillancourt M.-A., Kneeshaw D., Drapeau P., De Grandpré L., Claveau Y., Paré D. (2009). Forest ecosystem management: origins and foundations. In: Gauthier S., Vaillancourt M.A., Leduc A., De Grandpré L., Kneeshaw D., Morin H., Drapeau P., Bergeron Y. (eds.). Ecosystem management in the boreal forest. Presses de l’Université du Québec, Quebec.
Greene D.F., Johnson E.A. (1999). Modelling recruitment of Populus tremuloides, Pinus banksiana, and Picea mariana following fire in the mixedwood boreal forest. Canadian Journal of Forest Research 29: 462–473. http://dx.doi.org/10.1139/x98-211.
Greene D.F., Macdonald S.E., Haeussler S., Domenicano S., Noël J., Jayen K., Charron I., Gauthier S., Hunt S., Gielau E.T., Bergeron Y., Swift L. (2007). The reduction of organic-layer depth by wildfire in the North American boreal forest and its effect on tree recruitment by seed. Canadian Journal of Forest Research 37: 1012–1023. http://dx.doi.org/10.1139/X06-245.
Harper K., Broudreault C., De Grandpré L., Drapeau P., Gauthier S., Bergeron Y. (2003). Structure, composition, and diversity of old-growth black spruce boreal forest of the Clay Belt region in Quebec and Ontario. Environmental Reviews 11: S79-S98. http://dx.doi.org/10.1139/a03-013.
Harper K.A., Bergeron Y., Drapeau P., Gauthier S., De Grandpré L. (2005). Structural development following fire in black spruce boreal forest. Forest Ecology and Management 206: 293–306. http://dx.doi.org/10.1016/j.foreco.2004.11.008.
Hesketh M., Greene D.F., Pounden E. (2009). Early establishment of conifer recruits in the northern Rocky Mountains as a function of postfire duff depth. Canadian Journal of Forest Research 39: 2059–2064. http://dx.doi.org/10.1139/X09-120.
Holmes R.L. (1983). Computer-assisted quality control in tree-ring dating measurement. Tree-Ring Bulletin 43: 69–78.
Horton B.J., Groot A. (1987). Development of second-growth black spruce stands on peatlands in northeastern Ontario. Horton Forestry Services Limited, Stouffville, Ontario. 30 p.
Johnstone J.F., Kasischke E.S. (2005). Stand-level effects of soil burn severity on postfire regeneration in a recently burned black spruce forest. Canadian Journal of Forest Research 35: 2151–2163. http://dx.doi.org/10.1139/x05-087.
Kemball K.J., Wang G.G., Westwood R. (2006). Are mineral soils exposed by severe wildfire better seed-beds for conifer regeneration? Canadian Journal of Forest Research 36: 1943–1950. http://dx.doi.org/10.1139/x06-073.
Kenkel N.C. (1988). Pattern of self-thinning in jack pine: testing the random mortality hypothesis. Ecology 69: 1017–1024. http://dx.doi.org/10.2307/1941257.
Kenkel N.C., Hendrie M.L., Bella I.E. (1997). A long-term study of Pinus banksiana population dynamics. Journal of Vegetation Science 8: 241–254. http://dx.doi.org/10.2307/3237353.
Kneeshaw D., Bergeron Y. (1999). Spatial and temporal patterns of seedling and sapling recruitment within canopy gaps caused by spruce budworm. Ecoscience 6: 214–222.
Kneeshaw D., Gauthier S. (2003). Old growth in the boreal forest: a dynamic perspective at the stand and landscape level. Environmental Reviews 11: S99-S114. http://dx.doi.org/10.1139/a03-010.
Larson A.J., Churchill D. (2012). Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. Forest Ecology and Management 267: 74–92. http://dx.doi.org/10.1016/j.foreco.2011.11.038.
Le Goff H., Sirois L. (2004). Black spruce and jack pine dynamics simulated under varying fire cycles in the northern boreal forest of Quebec, Canada. Canadian Journal of Forest Research 34: 2399–2409. http://dx.doi.org/10.1139/x04-121.
Lussier J.-M., Morin H., Gagnon R. (2002). Mortality in black spruce stands of fire or clear-cut origin. Canadian Journal of Forest Research 32: 539–547. http://dx.doi.org/10.1139/x01-201.
Ministère des Ressources Naturelles, d.l.F.e.d.P. (2003). Manuel d’aménagement forestier. Gouvernement du Québec, Charlesbourg.
Miyanishi K., Johnson E.A. (2002). Process and patterns of duff consumption in the mixedwood boreal forest. Canadian Journal of Forest Research 32: 1285–1295. http://dx.doi.org/10.1139/x02-051.
Paquin R., Doucet R. (1992). Croissance en hauteur à long terme de la régénération préétablie dans des pessières noires boréales régénérées par marcottage, au Québec. Canadian Journal of Forest Research 22: 613–621. http://dx.doi.org/10.1139/x92-081.
Pardé J., Bouchon J. (1988). Dendrométrie. ENGREF, Nancy, France.
Ripley B.D. (1977). Modelling spatial patterns. Journal of the Royal Statistical Society, Series B Methodological 39: 172–212.
Rossi S., Morin H. (2011). Demography and spatial dynamics in balsam fir stands after a spruce budworm outbreak. Canadian Journal of Forest Research 41: 1112–1120. http://dx.doi.org/10.1139/x11-037.
Rossi S., Tremblay M.-J., Morin H., Levasseur V. (2009a). Stand structure and dynamics of Picea mariana on the northern border of the natural closed boreal forest in Quebec, Canada. Canadian Journal of Forest Research 39: 2307–2318. http://dx.doi.org/10.1139/X09-152.
Rossi S., Tremblay M.-J., Morin H., Savard G. (2009b). Growth and productivity of black spruce in even- and uneven-aged stands at the limit of the closed boreal forest. Forest Ecology and Management 258: 2153–2161. http://dx.doi.org/10.1016/j.foreco.2009.08.023.
Rossi S., Morin H., Tremblay M.-J. (2010). Growth and productivity of black spruce (Picea mariana) belonging to the first cohort in stands within and north of the commercial forest in Quebec, Canada. Annals of Forest Science 67: 807. http://dx.doi.org/10.1051/forest/2010043.
Rowe J.S. (1983). Concepts of fire effects on plant individuals and species. In: Wein R.W., MacLean D.A. (eds.). The role of fire in Northern circumpolar ecosystems. John Wiley & Sons, New York.
St-Pierre H., Gagnon R. (1992). Régénération après feu de l’épinette noire (Picea mariana) et du pin gris (Pinus banksiana) dans la forêt boréale, Québec. Canadian Journal of Forest Research 22: 474–481. http://dx.doi.org/10.1139/x92-062.
St-Pierre H., Gagnon R., Bellefleur P. (1991). Distribution spatiale de la régéneration après feu de l’épinette noire (Picea mariana) et du pin gris (Pinus banksiana) dans la forêt boréale, Réserve faunique Ashuapmushuan, Québec. Canadian Journal of Botany 69: 717–721. http://dx.doi.org/10.1139/b91-097.
Tremblay M.-J., Rossi S., Morin H. (2011). Growth dynamics of black spruce in stands located between the 51st and 52nd parallels in the boreal forest of Quebec, Canada. Canadian Journal of Forest Research 41: 1769–1778. http://dx.doi.org/10.1139/x11-094.
Wang G.G., Kemball K.J. (2005). Balsam fir and white spruce seedling recruitment in response to understory release, seedbed type, and litter exclusion in trembling aspen stands. Canadian Journal of Forest Research 35: 667–673. http://dx.doi.org/10.1139/x04-212.
Weiner J., Thomas S.C. (1986). Size variability and competition in plant monocultures. Oikos 47: 211–222. http://dx.doi.org/10.2307/3566048.
Wirth C., Lichstein J.W., Dushoff J., Chen A., Chapin III F.S. (2008). White spruce meets black spruce: dispersal, postfire establishment, and growth in a warming climate. Ecological Monographs 78: 489–505. http://dx.doi.org/10.1890/07-0074.1.
Zaninetti J.-M. (2005). Statistique spatiale: métodes et applications géomatiques. Lavoisier, Paris.
Total of 44 references
