Antonín Martiník  ,
Robert Knott,
Jan Krejza,
Jakub Černý
,
Robert Knott,
Jan Krejza,
Jakub Černý
Biomass production of Betula pendula stands regenerated in the region of allochthonous Picea abies dieback
Martiník A., Knott R., Krejza J., Černý J. (2018). Biomass production of Betula pendula stands regenerated in the region of allochthonous Picea abies dieback. Silva Fennica vol. 52 no. 5 article id 9985. https://doi.org/10.14214/sf.9985
Highlights
- Biomass equations for individual components of above-ground wood biomass estimation are presented for stands at the age of 4, 8, 17 and 22 years
- Peak of the mean annual increment was found at the age from 15 to 20 years and reached over 5.0 t ha–1 y–1 of dry biomass
- The share of the stem to the total biomass increased with stand age.
Abstract
The paper deals with production of above-ground biomass of silver birch (Betula pendula Roth) stands in the Czech Republic. One-year biomass dynamics was studied within chronosequence of birch stands at the age of 4–5, 8–9, 17–18 and 22–23 years. With the exception of the youngest stand, which was established by seeding, all experimental birch stands were regenerated naturally after the allochthonous spruce stands. Above-ground biomass (AB) was calculated from plot inventory data and biomass equations were parameterized from destructive sampling of biomass component of sampled trees. Results reveal that the peak of the mean annual increment (MAIABtotal) of birch stands can be expected at the age from 15 to 20 years. Additionally, the stand age, the value of basal area (BA) should be considered as a predictor of stand productivity. If the value of BA varied from 25 to 35 m2 ha–1, the MAI of the birch stands reached the range from 5.0 to 6.5 t of dry biomass per ha y–1 at the age ranging between 15 and 25 years. The stem/branch proportion increased with stand age, the stem relative proportion ranging from 75 to 90% of total above-ground biomass. According to the results of this study, birch stand biomass production and utilization is one of the approaches in terms of forest recovery management in large disturbed areas. Although, no silvicultural treatments were occurred in all analysed stands, the pre-commercial thinning method could increase stand productivity and stability as well.
Keywords
silver birch;
stem biomass;
branch biomass;
rotation period
-
Martiník,
Department of Silviculture, Faculty of Forestry and Wood Technology, Mendel University in Brno, 613 00 Brno, Czech Republic
 https://orcid.org/0000-0002-5906-8830
E-mail
martinik@mendelu.cz
https://orcid.org/0000-0002-5906-8830
E-mail
martinik@mendelu.cz
- Knott, Department of Silviculture, Faculty of Forestry and Wood Technology, Mendel University in Brno, 613 00 Brno, Czech Republic E-mail robert.knott@mendelu.cz
- Krejza, Global Change Research Institute CAS, v.v.i., Bělidla 4a, 603 00 Brno, Czech Republic E-mail krejza.j@czechglobe.cz
- Černý, The Forestry and Game Management Research Institute, Research Station at Opočno, Na Olivě 550, 517 73 Opočno, Czech Republic E-mail cerny@vulhmop.cz
Received 28 March 2018 Accepted 14 November 2018 Published 28 November 2018
Views 119534
Available at https://doi.org/10.14214/sf.9985 | Download PDF

1 Introduction
Natural distribution of silver birch (Betula pendula Roth) covers most of the Eurasian continent (Svoboda 1957; Hynynen et al. 2010). However, currently the composition of this species and its economic significance is high only in Northern and North-eastern Europe. For example, birch composition in Finland and Norway is about 16%, in Estonia it ranges between 20–30%, and in France or Czech Republic it is under 3% of the total volume (Hynynen et al. 2010; Kula 2011).
The birch, as a pioneer tree species with high tolerance to climatic extremes and soil conditions, often naturally occupies abandoned agricultural land and forest area after disturbances (Vacek 1991; Karlsson et al. 1998; Fischer et al. 2002; Huth and Wagner 2006; Suchockas 2002; Špulák et al. 2010). Fast growth and shade intolerance makes birch suitable for biomass production (Ferm 1993; Hynynen et al. 2010). Utilization of tree biomass as a renewable source of energy is a hot topic due to green energy concepts of the European Union (Krejzar 2008; EUCO 2014).
Recently, many studies were focused on biomass production and the ecological function of birch stands in abandoned agricultural land in the Northern and Eastern Europe region (Johansson 1999; Jõgiste et al. 2003; Uri et al. 2007; Varik et al. 2009). In Sweden, Johansson (1999) analysed above-ground biomass of ten Betula pendula stands and the same number of Betula pubescens Ehrh. stands. The age of the B. pendula stands ranged from 8 to 32 years and the total above-ground dry biomass varied from 5.7 to 175.3 t per ha. The mean annual increment (MAI) of these stands varied between 0.71–8.44 t ha–1. The peak of productivity was indicated at the age from 10 to 30 years, depending on the basal area of mentioned stands. Uri et. al. (2007) referred to biomass allocation and nutrient accumulation within an 8-year-old birch stand in Estonia. The total biomass of this stand was about 38.6 t ha–1, the stem biomass was 24 t ha–1 and branches about 3 t ha–1. However, the current above-ground biomass increment at this age was 11.9 t ha–1, the mean annual increment was 3.9 t ha–1, which was higher compared to that published in available literature.
Many investigations have already been conducted on birch biomass production as a forest tree species in the region of Northern and Eastern Europe with the exception of birch growth on abandoned agricultural lands (Repola 2008; Uri et al. 2012). Uri et al. (2012) found that above-ground biomass of birch stands at the age of 6–60 years varies from 25.7 to 220.5 t ha–1. Similarly, as on agricultural lands, the total biomass is related to stand age, growth conditions and stand density. Although, birch stand production in the analysed forests is higher than is stated in yield tables, the growth and production of birch on agricultural lands, under similar conditions, exceeds it even more (Uri et al. 2007, 2012).
The investigations performed in Central Europe are seldom compared to the birch biomass research conducted in Northern Europe. Zasada et al. (2014) found about 75 t ha–1 of above-ground birch biomass in 15-year-old stand in abandoned agricultural land in Central Poland. Jagodziński et al. (2017) also analysed biomass of birch stands growing on post-agricultural lands in Poland. The birch stand chronosequence showed that total above-ground biomass ranged from 0.5 t ha–1 to 166.5 t ha–1 in the age of 1–19 years. Špulák et al. (2016) showed that the total above-ground birch biomass of a 7-year-old birch stand growing on forest land comprises only 14.9 t ha–1. The MAI for this birch stand growing within the beech vegetation zone was 2.1 t ha–1.
The tree biomass may be estimated directly by the site and species-specific allometric relationship (Cienciala et al. 2006; Roxburgh et al. 2015; Goussanou et al. 2016). Tree biomass is quantified on the base of a known relationship between biomass and easily measurable tree characteristics (i.e. diameter at breast height, tree height, and crown length). This form of biomass estimation is currently considered the most accurate method of determining the amount of biomass on the tree level and also in forest ecosystems (Goussanou et al. 2016).
The current share of renewable sources of energy (RSE) of the total primary energy is about 6% in the conditions of Czech Republic (Weger and Jiránek 2003). By 2040, the share of these sources should increase to 17%. The significant component of RSE is a bioenergy produced from agricultural and forest residues and from energy crops (Jõgiste et al. 2003; Lewandowski et al. 2006).
In the forestry sector, the more effective use of forest processes and dynamics could lead to higher wood productivity and also increase the potential of forest as a source of renewable energy. The wider utilization of the stands of pioneer tree species regenerated after disturbances should be one of these approaches (Pommerening and Murphy 2004; Unselt and Bauhus 2012; Stark et al. 2013). Although silver birch is not deemed as the main commercial forest species in the Czech Republic, it is often regenerated naturally in disturbed forest areas (Martiník et al. 2014). The managing of these stands in terms of birch utilization is a critical question for forest management and research as well.
The working hypothesis of this paper is that birch stands regenerated in large clear-cuts after dieback of allochthonous spruce stands in the region of Czech Republic can, in terms of their productivity, be important source of renewable energy. Forest regeneration in large clearings, originating after spruce stands, has been a serious problem, where the positive environmental effects of pioneer species stands have been known for a long time (Zakopal 1958; Stark et al. 2013; Martiník et al. 2017). The main aim of this study was to determine production of biomass and the increment of such pioneer (birch) stands growing on fertile sites in the Czech Republic.
The specific aims of this study were: (a) to analyse allocation of above-ground biomass depending on the stand age and stand density; (b) to evaluate the share of tree components (stems, branches) on the total above-ground biomass of experimental stands; (c) to establish basic recommendations for birch stand biomass management.
2 Material and methods
2.1 Experimental stands
For the purpose of this study, four young silver birch stands (age 4–22 years in 2015), which regenerated in a clearing after conifer (spruce) stands, were selected. All of these stands are located throughout Czech Republic on rich soils in an oak–beech forest vegetation zone (Table 1, Fig. 1).
| Table 1. Basic characteristic of analysed stands. | ||||||
| Stand | Latitude, longitude | Age (year) | Upper stand height (m) | Average (Min – max.) number of trees (pcs ha-1) | Mean basal area (m2 ha-1) | Average DBH (±SD) (cm) |
| Altitude (m) | ||||||
| Tipecek | 49°19´N, 16°44´E | 4 | 4 | 42 890 (2000 – 95 000) | 2.8670 | 0.72 (0.57) |
| 510 | 5 | 43 670 (4000 – 99 000) | 4.7680 | 0.97 (0.66) | ||
| Breziny | 49°69´N, 18°19´E | 8 | 7 | 18 720 (6400 – 48 000) | 7.2328 | 1.89 (1.18) |
| 285 | 9 | 18 720 (6400 – 48 000) | 9.8538 | 2.24 (1.40) | ||
| Dubska | 49°88´N, 18°10´E | 17 | 15 | 4720 (2800 – 6400) | 22.0204 | 7.00 (3.23) |
| 324 | 18 | 4720 (2800 – 6400) | 23.3443 | 7.18 (3.40) | ||
| Skrchov | 49°59´N, 16°55´E | 22 | 17 | 2800 (2000 – 3600) | 25.7693 | 10.20 (3.66) |
| 475 | 23 | 2800 (2000 – 3600) | 26.4830 | 10.33 (3.74) | ||
Fig. 1. Location of the experimental stands in the Czech Republic.
The oak–beech vegetation zone is generally characterized by long-term annual average temperatures from 6.5 °C to 7.5 °C and with annual precipitation ranging between 650–700 mm (Poleno et al. 2007a, 2007b). However, the climatic change leads to an increase in temperature and a decrease in precipitation in this zone (Hlásny et al. 2014). The rich soil variant of this zone is naturally covered by a productive beech (Fagus sylvatica L.) forest with a mixture of oak (Quercus petraea (Matt.) Liebl.), fir (Abies alba Mill.) or some minority of broadleaf species (Poleno et al. 2007a, 2007b; Ellenberg 2009). During the 19th and 20th centuries, most of these forests were replaced by coniferous trees with Norway spruce (Picea abies (L.) H. Karst.) prevailing. Since the end of the 20th century, most of the allochthonous spruce stands in the region of Central Europe have showed dieback and decline (Fanta 1997; Holuša and Liška 2002; Spiecker et al. 2004).
The first stand Tipecek (area of 0.3 ha) was established by birch seeding in 2010 after wind calamity Antonín. The second stand Breziny (area of 0.25 ha) regenerated naturally in 2007 after wind disturbance Kyrill. The Dubska stand (area of 0.30 ha) and the Skrchov stand (an area of 0.75 ha) regenerated naturally after exploitation cutting by private companies in 1996 and 1993 respectively.
In all analysed stands, birch is the dominant species. The shares of other species are under 5% in terms of number of trees and these species occurred mainly in lower layers of the analysed stands. From these species, larch and sycamore were observed in the case of Tipecek, shrubs in Breziny and oak within the Dubska and Skrchov stands. These trees were excluded from all analyses.
2.2 Inventory and sample trees
The analyses were based on stand inventory, on destructive measurement of sample birch trees in order to develop above-ground biomass regression equations and on the consecutive application of these equations to the stand level (Johansson 1999). All measurements were conducted repeatedly at the end of the growing seasons of 2015 and 2016.
Stand inventory was conducted for all stands on permanent plots. The plots were established in regular grids to represent the variability of stand structure – ten 10 m2 inventory plots at younger stands (Tipecek, Breziny) and ten 25 m2 inventory plots at older stands (Dubska, Skrchov).
The stem diameter at breast height (DBH) of all live trees higher than 1.3 m within the inventory plots of all stands was measured in the years 2015 and 2016. Sample trees were selected for each stand to represent the diameter distribution of the inventory plots within stands. In total, the 37 and 36 sample trees were felled in young stands in Tipecek and Breziny, respectively; and 8 and 7 sample trees were harvested in old stands in Dubska and Skrchov, respectively in 2016.
The allometric relationship equations are usually based on a limited number of sampled trees (Zianis et al. 2005). The number of sampled trees varies from several to thousands of individuals, but the most common number of sampled trees belongs to the interval from 6 to 40 for the site-specific allometric relationship (Krejza et al. 2017). A lower number of sampled trees can be used when the sampled trees are selected from tree diameter distribution in the forest stand (Roxburgh et al. 2015). The final number of sampled trees was estimated based on the combination of an acceptable amount of work to obtain the measurements (Picard et al. 2012; Roxburgh et al. 2015) and the proper selection of sampled trees in the experimental stands (inventory plots).
The sampled trees were selected from four experimental stands using the technique of quantiles of the total. This technique calculates the diameters of a series of sample trees where each represents the same fraction of the selected biometric parameters in the experimental plot (Čermák et al. 2004). All trees in the inventory plots of experimental stands were sorted according to size of basal area (BA) and cumulatively. The basal area of the experimental stand was divided by the number of sampled trees (k) in experimental stand (Bep = BA / k). The sizes of the sampled trees representing individual fractions were then calculated using Bep multiplied by factors k – 0.5 (Čermák et al. 2004).
Stand inventory and also the measurement of sample birch trees were carried out during the leafless period, which means that birch biomass represented only above-ground (wood) biomass (AB). After sample tree felling, their height (i. e. length after felling) and DBH were measured. The stems were divided into three parts and fresh weights of the stem parts were determined. The stem disc samples were taken from the middle of each section. These discs were weighed and dried at 80 °C until they reached a constant weight for it to be possible to obtain the conversion factor for dry weight of the stem. After a fresh weighing of branches, a representative branch from each third of the crown was selected. These branches were weighed and dried at 80 °C until they reached a constant weight for it to be possible to obtain the conversion factor for dry weight of the branches. Each tree component was weighed accurate to 10 g.
2.3 Data analysis
Allometric relationships between DBH and dry above-ground biomass (ABstem, ABbranch, and ABtotal = ABstem + ABbranch) of sample trees in all stands were calculated (12 biomass equations in total). We used the power model for such relationships (Johansson 1999; Picard et al. 2012):
![]()
where AB is dry above-ground biomass of tree components (kg); DBH is stem diameter at breast height; a, b are estimated parameters of biomass equations for individual tree components (Picard et al. 2012). The nls function (i.e. nonlinear least-squares method) in the R software (R Development Core Team 2012) was used for the allometric relationship fitting.
The parameters of biomass equations are summarized in Table 2. All biomass equations demonstrated high values of coefficient of determination (R2) with DBH (R2 ranged from 0.92 to 0.99).
| Table 2. Estimated parameters (a, b) of biomass equations (Eq. 1) for individual tree components of above-ground dry biomass (AB) of birch. Abbreviations: N – number of sampled tree; R2 – coefficient of determination, SE – standard error of parameter. | |||||||||||
| ABtotal | ABstem | ABbranch | |||||||||
| Stand | Age | N | a (SE) | b (SE) | R2 | a (SE) | b (SE) | R2 | a (SE) | b (SE) | R2 |
| Tipecek | 5 | 37 | 0.14107 (0.00946) | 2.18716 (0.07429) | 0.980 | 0.11431 (0.00672) | 2.01210 (0.06639) | 0.981 | 0.02511 (0.00518) | 2.78590 (0.21770) | 0.917 |
| Breziny | 9 | 36 | 0.10941 (0.00832) | 2.40663 (0.04607) | 0.993 | 0.10189 (0.00813) | 2.24554 (0.04897) | 0.990 | 0.01422 (0.00139) | 2.87939 (0.05751) | 0.993 |
| Dubska | 18 | 8 | 0.22062 (0.06674) | 2.15318 (0.12325) | 0.993 | 0.22533 (0.08166) | 2.08008 (0.14806) | 0.989 | 0.00872 (0.00439) | 2.68412 (0.20206) | 0.989 |
| Skrchov | 23 | 7 | 0.11792 (0.06954) | 2.40272 (0.23525) | 0.971 | 0.12251 (0.07621) | 2.35227 (0.24854) | 0.966 | 0.00192 (0.00104) | 3.05616 (0.21198) | 0.987 |
Dry above-ground biomass (separately for stem, branches, and total = stem + branches) for all individual trees from inventory in all stands was predicted using obtained biomass equations.
The weight of total dry above-ground biomass was calculated as the sum of the AB of all the trees and then recalculated per ha. Variability of AB between plots was evaluated by the coefficient of variation (CV) as the ratio of the standard deviation to the mean value.
We applied the Chapman-Richards growth function (Zeide 1993) to all stand data pertaining to total dry above-ground biomass (ABtotal):
![]()
where a, b, and c are the parameters of the function and t is the stand age.
Curves of the current total dry above-ground biomass annual increment (CAIABtotal) for the same data was constructed by the function:
![]()
The mean total dry above-ground biomass annual increment was calculated (MAIABtotal) by the function:
![]()
where CAIABtotal is the current total dry above-ground biomass annual increment and t is age.
3 Results
Despite the high variability in stand density, there was an evident decrease in average birch tree density with stand age – from 43 700 to 2800 trees per ha (Table 1). Also, DBH of birch trees within the stands varies and the average value increases with stand age – from 0.72 to 10.33 cm (Table 1). According to this variability, the DBH of the sample trees for the biomass equations ranged from 0.1 to 2.9 (Tipecek), 0.4 to 6.3 (Breziny), 1.3 to 13.1 (Dubska) and from 5.1 to 14.0 cm (Skrchov).
The total dry above-ground biomass (ABtotal) of birch stands ranged from 5.8 (Tipecek) to 108.9 (Skrchov) t ha–1 at the age from 4 to 23 years (Table 3). The average MAIABtotal of these stands varied from 1.5 to 5.1 t ha–1. The lowest values (1.5 and 1.9 t ha–1) were reached in the stand at the age of 4 (5) years (Tipecek) and the highest values (5.0 and 5.1 t ha–1) were noted in stands Dubska at the age of 17 (18) years. The MAIABtotal value of the 8- and 9-year-old Březiny stand was 2.0 and 2.6 t ha–1, respectively and for the 22- and 23-year-old Skrchov stand it was 4.7 and 4.8 t ha–1, respectively. The highest value of CAIABtotal (7.2 t ha–1) was indicated in the Breziny stand at the age between 8 and 9 years. The lowest value of CAIABtotal was found in the Skrchov stand (2.7 t ha–1) at the age between 22 and 23 years (Table 3). The CAIABtotal value of the Tipecek and Dubska stands were similar – close to 3.5 t ha–1. The variability of AB rapidly decreased after reaching five years and in the following age stages it fluctuated around 30%. The lowest variability of total biomass and its components were found for the Dubska stand at the age 17 and 18, respectively (Fig. 2).
| Table 3. Mean dry above-ground biomass for individual tree components (AB) (t ha–1) for analysed birch stands (MAI – mean annual increment, CAI – current annual increment). | ||||||||
| Stand | Tipecek | Breziny | Dubska | Skrchov | ||||
| Age | 4 | 5 | 8 | 9 | 17 | 18 | 22 | 23 |
| ABbranch | 1.39 | 2.38 | 3.51 | 5.54 | 11.38 | 12.18 | 9.07 | 9.44 |
| ABstem | 4.41 | 6.98 | 12.23 | 17.37 | 75.43 | 77.92 | 97.05 | 99.36 |
| ABtotal | 5.80 | 9.36 | 15.73 | 22.91 | 86.81 | 90.10 | 106.12 | 108.80 |
| MAIABtotal | 1.45 | 1.87 | 1.97 | 2.55 | 5.11 | 5.01 | 4.82 | 4.73 |
| CAIABtotal | 3.55 | 7.18 | 3.29 | 2.67 | ||||
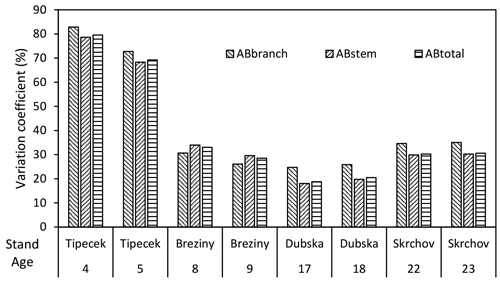
Fig. 2. Variability of total above-ground biomass (AB) and its components (stem and branches) within analysed birch stands.
The proportion of the stem to the total biomass increased with stand age and varied between 75% and 91%, respectively (Fig. 3). The increase in DBH led to a decrease in the proportion of the stem to the total biomass within the same stand on the sample tree level (Fig. 4).
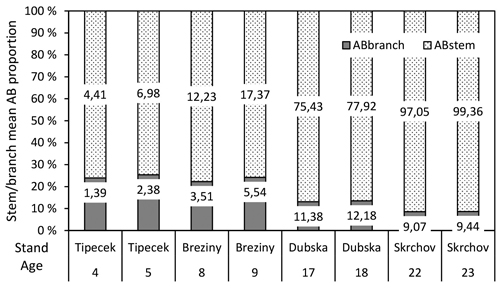
Fig. 3. Stem/branch mean AB proportion (%) within analysed stands and ages (numbers inside bars – AB in t ha-1).
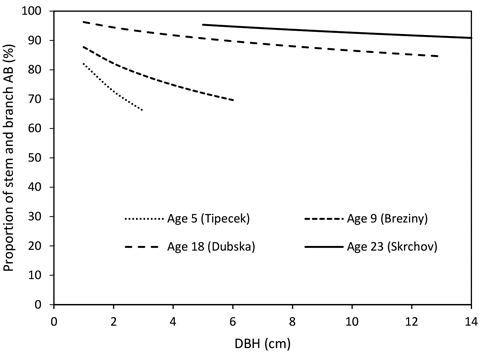
Fig. 4. Proportion of stem and branch above-ground biomass (AB) of sample trees of analysed stands according to DBH (stem AB – below curves, branch AB – above curves).
The ABtotal as well as MAIABtotal showed a positive correlation with the basal area of the plots within the analysed stands, although there was a different trend found in the case of young (Tipecek, Breziny) and old (Dubska, Skrchov) stands (Fig. 5). Maximum value of ABtotal and MAIABtotal for younger stands (4–9 years) was around 40 t ha–1 and 4.5 t ha–1 per year, respectively; both on the level of 20 m2 ha–1 of BA. In the case of older stands, the maximum BA on the level of 35 m2 ha–1 reached an ABtotal of about 150 t ha–1 and a MAIABtotal of around 6.5 t ha–1 per year.
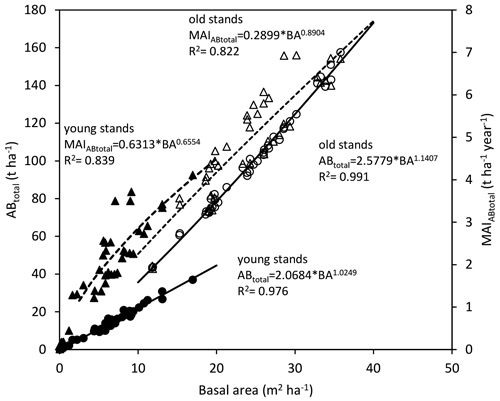
Fig. 5. Total above-ground biomass (ABtotal) and total dry above-ground biomass mean annual increment (MAIABtotal) in relation to basal area on plots in young and old stands (points represent values on each inventory plot: full circles – ABtotal of young stands, empty circles – ABtotal of old stands, full triangles – MAIABtotal of young stands, empty triangles – MAIABtotal of old stands).
Model curves carried out on the results from the experimental stands show that the peak of MAI of birch stands, which should be close to 5 t ha–1 per year can be expected at the age from 15 to 20 years (Fig. 6).
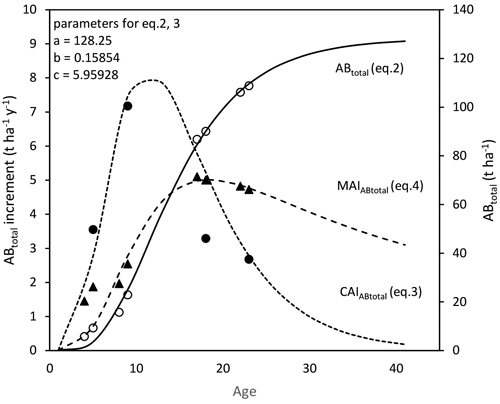
Fig. 6. Observed data and models of total above-ground biomass (ABtotal), current annual increment (CAIABtotal), and dry above-ground biomass mean annual increment (MAIABtotal) (points represent values found in observed years).
4 Discussion
Regarding the birch height, the best site-index classes for all of the analysed stands can be expected (Černý and Pařez 1998; Lockow 1997). This should be the result of natural conditions (lower altitudes and rich soil), but also of previous species composition (spruce monoculture). Long-term cultivation of allochthonous spruce leads to degraded soil properties in the region of Central Europe (Fanta 1997; Holuša and Liška 2002). On the other hand, conditions of previous spruce cultivation seem to be favourable for birch regeneration and growth (Huth and Wagner 2006; Martiník et al. 2017), which is known as a soil improving species (Zakopal 1958; Stark et al. 2013). At present, the huge dieback of these spruce stands in Central Europe is a serious problem and also a challenge for new silviculture treatments and species composition (Brang et al. 2014; Hlásny et al. 2014). One of these treatments is cultivation of birch stands for biomass production. In our experiment we studied above-ground biomass production of four birch stands regenerated after allochtonous spruce stands at the ages from 4 to 22 years during a period of one year.
The ABtotal production of the analysed birch stands increased with stand age to 109 t ha–1 but, from the production and silvicultural points of view, the MAIABtotal is more important than the total biomass (Assmann 1970; Pretzsch 2009). Based on the results of our investigations, the potential of birch stands can reach a MAIABtotal close to 5 t ha–1 y–1 of dry wood mass in the forest area of Central Europe. This estimation corresponds with the results published by many authors for the region of Northern and North-Eastern Europe (Ferm 1993; Johansson 1999; Uri et al. 2012; Zasada et al. 2014). Johansson (1999) found the MAI for a birch stand growing on abandoned farmland in Sweden ranging up to 8.44 t of dry weight ha–1 y–1, including leaves. Also, Uri et al. (2012) found the MAI of birch stands to be under 5 t of dry mass ha–1 y–1, including leaves, in total above-ground biomass. The proportion of the leaves in the total above-ground biomass ranged from 1 to 18% and decreases with stand age (Johansson 1999; Zasada et al. 2014; Špulák et al. 2016). Leaf biomass was not included in this study because felling and the using of the stands for biomass often takes place in leafless periods.
The model curves based on the data from experimental birch stands showed that the MAI of investigated stands culminated in the age ranging from 15 to 20 years. This finding agrees well with results published by Ferm (1993) for Finnish conditions. On the other hand, birch stands analysed by Johansson (1999) show a peak of the MAI at the age of 12 years. This is probably the result of faster growth of birch stands on the abandoned agricultural lands studied by Johansson (1999). The relation of birch stand age and biomass production was not confirmed by Uri et al. (2012). The highest MAI of birch stands was found to be similar at the age 13 and 45 years. The age should be considered as only one of the factors that influence the culmination of the MAI of above-ground biomass of birch stands growing under similar natural conditions.
Additionally, there was a correlation between the BA and MAI of birch stands of a similar age (17–22 and 4–9 years, respectively). Similar results were published by Johansson (1999) who noted that the level of the BA can be considered as a prerequisite for birch stand utilization. According to our results, birch forest stands on fertile soil in the region of Central Europe with a BA ranging from 25 to 35 m2 ha–1 can reach a MAI of 5.0 to 6.5 t of dry mass ha–1 y–1 at the age between 17 to 22 years. Although, a high BA was related to high stand density, there were no silvicultural treatments in any of the analysed stands. According to Uri et al. (2007), birch stand biomass productivity at the same age increased with stand density, but this dependency is not linear and upon the exceeding of a certain limit, further increase in density is not accompanied anymore by an increase in biomass.
High tree densities of naturally regenerated birch and only self-thinned stands are accompanied by a low average tree diameter (Rytter and Werner 2007). This can lead to sensitivity of birch stands to snow damage (Nykänen et al. 1997; Pӓӓtalo 2000; Martiník and Mauer 2012). On the other hand, an appropriate clearing and thinning regime can improve stand stability and tree vitality as well (Nykänen et al. 1997; Zàlïtis and Zàlïtis 2007; Hynynen et al. 2010). Simultaneously, pre-commercial thinning performed in birch stands leads to an increase in BA compared to that in the non-thinned stands (Rytter and Werner 2007; Zàlïtis and Zàlïtis 2007). The thinning method of birch stands, focused on biomass production, should be different from that for the production of high-quality timber, as described by Hein et al. (2009). In general, in the application of this method, fast growing but low stem quality trees are not removed because they can contribute to an extremely increased value of BA in the target age.
Compared to high-quality birch timber tree management, the biomass-oriented system includes branches in the total stand productivity (Repola 2008; Bronisz et al. 2016). On the sample tree level, the proportion of branches ranged from 5 to 35% of the total above-ground biomass and increased together with DBH within the same stand. This should be in contrary to the result published by Johansson (1999). He observed that the proportion of branches decreased with the increase in DBH, but his result was based on dominant sample trees from stands of different ages. Also, a decreasing proportion of branches relating to tree age was confirmed in our experiment and this, additionally, leads to an increase in the stem/branch proportion with the stand age. The share of the proportion of branches in total dry above-ground biomass ranged from 10 to 25 % and corresponded with the results by Uri et al (2012) and Zasada et al. (2014) for birch stands of similar age, growing on abandoned agricultural lands. On the other hand, the proportion of branches is very sensitive to stand density (Nykänen et al. 1997; Hynynen et al. 2010).
Although, the production of fast growing tree species on farmlands can reach from 10 to 15 t of dry mass ha–1 y–1 in the Central Europe region, the practical results are often lower (Weger 2003; Trnka et al. 2008; Martiník et al. 2015). Based on the present study, the production of birch stands growing on fertile forest soil can be lower than maximum but close to practical production of intensive managed woody biomass. Simultaneously, the management costs of birch stands growing on forest soil are very low in comparison to intensive plantations. Similarly, low costs can be expected in the case of extensive establishment of birch stands growing on farmland where productivity can be comparable to that of forest lands (Johansson 1999; Uri et al. 2007; Špulák et al. 2014). Most stands (3) analysed in this study were regenerated naturally and only in the case of Tipecek, low-cost seeding was used as a regeneration method (Martiník at al. 2014).
5 Conclusion
Presented results show that production of above-ground biomass of birch stands in the Central Europe region can be comparable to that published for North-Eastern Europe. Naturally or artificially established (i.e. by seeding) birch stands growing on fertile soil after allochthonous spruce stands can reach even more than 5 t ha–1 y–1 of dry biomass. The peak of the MAI (and also of the rotation period), for these stands, can be expected in the age exceeding 15 (20) years. Furthermore, the birch stand age and the basal area of mentioned stands can be considered as factors influencing stand productivity; the greater the basal area, the higher the biomass production is expected.
Acknowledgements
This study was supported by Internal Grant Agency (IGA) project no. LDF_PSV_2018002 and by TACR, project no. TJ01000309.
References
Assmann E. (1970). The principles of forest yield study. Pergamon press, Oxford. 505 p.
Brang P., Spathelf P., Larsen J.B., Bauhus J., Bončína A., Chauvin C., Drössler L, García-Güemes C, Heiri C, Kerr G, Lexer MJ, Mason B, Mohren F, Mühlethaler U, Nocentini S, Svoboda M. (2014). Suitability of close-to-nature silviculture for adapting temperate european forests to climate change. Forestry 87(4): 492–503. https://doi.org/10.1093/forestry/cpu018.
Bronisz K., Strub M., Cieszewski C., Bijak S., Bronisz A., Tomusiak R., Wojtan R., Zasada M. (2016). Empirical equations for estimating aboveground biomass of Betula pendula growing on former farmland in central Poland. Silva Fennica 50(4) article 1559. https://doi.org/10.14214/sf.1559.
Čermák J., Kučera J., Nadezhdina N. (2004). Sap flow measurements with some thermodynamic methods, flow integration within trees and scaling up from sample trees to entire forest stands. Trees 18(5): 529–546. https://doi.org/10.1007/s00468-004-0339-6.
Černý M., Pařez J. (1998). Růstové tabulky dřevin České republiky. [Growth tables of trees in the Czech Republic]. Jílové u Prahy, Ústav pro výzkum lesních ekosystémů. 119 p.
Cienciala E., Černý M., Tatarinov F., Apltauer J., Exnerová Z. (2006). Biomass functions applicable to Scots pine. Trees 20(4): 483–495. https://doi.org/10.1007/s00468-006-0064-4.
Ellenberg H. (2009). Vegetation ecology of Central Europe. Cambridge University Press, Cambridge. 756 p.
European Council conclusions (EUCO), No. EUCO 169/14 (2014). European Council, Brussel. 16 p. http://www.consilium.europa.eu/uedocs/cms_data/docs/pressdata/en/ec/145397.pdf. [Cited 14 March 2018].
Fanta J. (1997). Rehabilitating degraded forests in Central Europe into self–sustaining forest ecosystems. Ecological Engineering 8(4): 289–297. https://doi.org/10.1016/S0925-8574(97)00024-4.
Ferm A. (1993). Birch production and utilization for energy. Biomass and Bioenergy 4(6): 391–404. https://doi.org/10.1016/0961-9534(93)90061-8.
Fischer A., Lindner M., Abs C., Lasch P. (2002). Vegetation dynamics in central european forest ecosystems (near-natural as well as managed) after storm events. Folia Geobotanica 37(1): 17–32. https://doi.org/10.1007/BF02803188.
Goussanou C.A., Guendehou S., Assogbadjo A.E., Kaire M., Sinsin B., Cuni-Sanchez A. (2016). Specific and generic stem biomass and volume models of tree species in a West African tropical semi-deciduous forest. Silva Fennica 50(2) article 1473. https://doi.org/10.14214/sf.1474.
Hein S., Winterhalter D., Wilhelm G., Kohnle U. (2009). Wertholzproduktion mit der Sandbirke (Betula pendula Roth): waldbauliche Möglichkeiten und Grenzen. [Timber production with silver birch [Betula pendula Roth]: Chances and silvicultural constraints]. Allgemeine Forst Und Jagdzeitung 180: 206–219. [In German with English summary].
Hlásny T., Mátyás C., Seidl R., Kulla L., Merganičová K., Trombik J., Dobor L., Barcza Z., Konôpka B. (2014). Climate change increases the drought risk in Central European forests: what are the options for adaptation? Forestry Journal 60(1) 5–18. https://doi.org/10.2478/forj-2014-0001.
Holuša J., Liška J. (2002). Hypotéza hynutí smrkových porostů ve Slezsku (Česka republika). [Hypothesis of spruce forests decline and dying in Silesia (Czech Republic)]. Zprávy lesnického výzkumu 47: 9–15. [In Czech with English summary].
Huth F., Wagner S. (2006). Gap structure and establishment of Silver birch regeneration (Betula pendula Roth.) in Norway spruce stands (Picea abies L. Karst.). Forest Ecology and Management 229(1–3): 314–324. https://doi.org/10.1016/j.foreco.2006.04.010.
Hynynen J., Niemistö P., Viherä-Aarnio A., Brunner A., Hein S., Velling P. (2010). Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83(1): 103–119. https://doi.org/10.1093/forestry/cpp035.
Jagodziński A.M., Zasada M., Bronisz K., Bronisz A., Bijak S. (2017). Biomass conversion and expansion factors for a chronosequence of young naturally regenerated silver birch (Betula pendula Roth) stands growing on post-agricultural sites. Forest Ecology and Management 384: 208–220. https://doi.org/10.1016/j.foreco.2016.10.051.
Jõgiste K., Vares A., Sendrós M. (2003). Restoration of former agricultural fields in Estonia: comparative growth of planted and naturally regenerated birch. Forestry: An International Journal of Forest Research 76(2): 209–219. https://doi.org/10.1093/forestry/76.2.209.
Johansson T. (1999). Biomass equations for determining fractions of common and grey alders growing on abandoned farmland and some practical implications. Biomass and Bioenergy 18(2): 147–159. https://doi.org/10.1016/S0961-9534(99)00078-1.
Karlsson A., Albrektson A., Forsgren A., Svensson L. (1998). An analysis of successful natural regeneration of downy and silver birch on abandoned farmland in Sweden. Silva Fennica 32(3): 229–240. https://doi.org/10.14214/sf.683.
Krejza J., Světlík J., Bednář P. (2017). Allometric relationship and biomass expansion factors (BEFs) for above- and below-ground biomass prediction and stem volume estimation for ash (Fraxinus excelsior L.) and oak (Quercus robur L.). Trees 31(4): 1303–1316. https://doi.org/10.1007/s00468-017-1549-z.
Krejzar T. (ed.) (2008). Národní lesnický program II na období do roku 2013. [National Forest Program II for the period up to 2013]. ÚHÚL, Praha. 20 p. http://www.uhul.cz/nase-cinnost/narodni-lesnicky-program. [Cited 14 March 2018].
Kula E. (2011). Bříza a její význam pro trvalý rozvoj lesa v imisních oblastech. [Birch and its importance for the sustainable development of a forest in air-polluted areas]. Kostelec nad Černými lesy, Lesnická práce. 276 p. [In Czech].
Lewandowski I., Weger J., van Hooijdonk A., Havlickova K., van Dam J., Faaij A. (2006). The potential biomass for energy production in the Czech Republic. Biomass and Bioenergy 30(5): 405–421. https://doi.org/10.1016/j.biombioe.2005.11.020.
Lockow K.W. (1997). Die neue Ertragstafel für Sandbirke-Aufbau und Bestandesbehandlung. [The new yield table for Silver birch – construction and stand treatment]. Beiträge für Forstwirtschaft und Landschaftsökologie 31(2): 75–84.
Martiník A., Mauer O. (2012). Snow damage to birch stands in Northern Moravia. Journal of Forest Science 58(4): 181–192. https://doi.org/10.17221/48/2011-JFS.
Martiník A., Dobrovolný L., Hurt V. (2014). Comparison of different forest regeneration methods after windthrow. Journal of Forest Science 60: 190–197. https://doi.org/10.17221/66/2013-JFS.
Martiník A., Adamec Z., Knott R., Stuchlý O. (2015). Production and economic parameters of a poplar (J 105) coppice plantation with different length of the first rotation in the conditions of the bohemian-moravian highlands. Acta Universitatis Agriculturae Et Silviculturae Mendelianae Brunensis 63(5): 1499–1504. https://doi.org/10.11118/actaun201563051499.
Martiník A., Adamec Z., Houška J. (2017). Production and soil restoration effect of pioneer tree species in a region of allochthonous norway spruce dieback. Journal of Forest Science 63(1): 34–44. https://doi.org/10.17221/98/2016-JFS.
Nykänen M.-L., Broadgate M., Kellomäki S., Peltola H., Quine C. (1997). Factors affecting snow damage of trees with particular reference to European conditions. Silva Fennica 31(2): 193–213. https://doi.org/10.14214/sf.a8519.
Päätalo M.-L. (2000). Risk of snow damage in unmanaged and managed stands of Scots pine, Norway spruce and birch. Scandinavian Journal of Forest Research 15(5): 530–541. https://doi.org/10.1080/028275800750173474.
Picard N., Saint-Andre L., Henry M. (2012). Manual for building tree volume and biomass allometric equations: from field measurement to prediction. Food and Agricultural Organization of the United Nations, Rome, and Centre de Coopération Internationale en Recherche Agronomique pour le Développement, Montpellier. 215 p.
Poleno Z., Vacek S. et al. (2007a). Pěstování lesů I. Ekologické základy pěstování lesů. [Silviculture I. Ecological bases of silviculture]. Kostelec nad Černými lesy, Lesnická práce. 313 p. [In Czech].
Poleno Z., Vacek S. et al. (2007b). Pěstování lesů II. Teoretická východiska pěstování lesů. [Silviculture II. Theoretical background of silviculture]. Kostelec nad Černými lesy, Lesnická práce. 464 p. [In Czech].
Pommerening A., Murphy S.T. (2004). A review of the history, definitions and methods of continuous cover forestry with special attention to afforestation and restocking. Forestry 77(1): 27–44. https://doi.org/10.1093/forestry/77.1.27.
Pretzsch H. (2009). Forest dynamics, growth and yield: from measurement to model. Springer-Verlag, Berlin Heidelberg. 664 p.
R Development Core Team. (2012). R: a language and environment for statistical computing. Vienna, Austria. http://www.r-project.org/.
Repola J. (2008). Biomass equations for birch in Finland. Silva Fennica 42(4): 605–624. https://doi.org/10.14214/sf.236.
Roxburgh S.H., Paul K.I., Clifford D., England J.R., Raison R.J. (2015). Guidelines for constructing allometric models for the prediction of woody biomass: how many individuals to harvest? Ecosphere 6(3): 1–27. https://doi.org/10.1890/ES14-00251.1.
Rytter L., Werner M. (2007). Influence of early thinning in broadleaved stands on development of remaining stems. Scandinavian Journal of Forest Research 22(3): 198–210. https://doi.org/10.1080/02827580701233494.
Spiecker H., Hansen J., Klimo E., Skovsgaard J.P., Sterba H., Teuffel K. (ed.) (2004). Norway spruce conversion options and consequences. Brill – Leiden, Boston. 269 p.
Špulák O., Souček J., Bartoš J., Kacálek D. (2010). Potenciál mladých porostů s dominancí břízy vzniklých sukcesí na neobhospodařované orné půdě. [Potential of young stands with birch dominance established by succession on abandoned agricultural land]. Zprávy lesnického výzkumu 55(3): 165–170. [In Czech with English summary].
Špulák O., Souček J., Leugner H. (2014). Variabilita struktury mladých převážně březových porostů vzniklých sukcesí na holinách kalamitního charakteru. [Stand structure variability of young stands with birch dominance established by succession on extreme clear-cut areas]. In: Štefančík I. (ed.). Proceedings of Central European Silviculture. Zvolen, 9.–10. 9. 2014. Národné lesnícke centrum, Zvolen. p. 68–74. [In Czech with English summary].
Špulák O., Souček J., Leugner J. (2016). Nadzemní biomasa, živiny a spalné teplo v mladém sukcesním porostu přípravných dřevin. [Aboveground biomass, nutrients and combustion heat of young successional forest stand of pioneer species]. Zprávy lesnického výzkumu 61(2): 132–137. [In Czech with English summary].
Stark H., Nothdurft A., Bauhus J. (2013). Allometries for widely spaced Populus ssp. and Betula ssp. in nurse crop systems. Forests 4(4): 1003–1031. https://doi.org/10.3390/f4041003.
Suchockas V. (2002). Seed dispersal and distribution of silver birch (Betula pendula) naturally regenerating seedlings on abandoned agricultural land at forest edges. Baltic Forestry 8(2): 71–76.
Svoboda P. (1957). Lesní dřeviny a jejich porosty, Část III. [Forest trees and forest stands, Part III.]. SZN, Praha. 457 p. [In Czech].
Trnka M., Fialová J., Koutecký V., Fajman M., Žalud Z., Hejduk S. (2008). Biomass production and survival rates of selected poplar clones grown under a short-rotation system on arable land. Plant Soil Environment 54(2): 78–88. https://doi.org/10.17221/437-PSE.
Unselt R, Bauhus J. (2012). Energie-Vorwälder – Alternative Bewirtschaftungsformen zur Steigerung der energetisch nutzbaren Biomasse im Wald durch Integration von schnell wachsenden Baumarten. [Energy-Pioneer forest – alternative forms of management for the increase of the energetically usable biomass in the forest by integration of fast-growing tree species]. Waldbau-Institut Albert-Ludwigs-Universität Freiburg. 2013 p.
Uri V., Vares A., Tullus H., Kanal A. (2007). Above-ground biomass production and nutrient accumulation in young stands of silver birch on abandoned agricultural land. Biomass and Bioenergy 31(4): 195–204. https://doi.org/10.1016/j.biombioe.2006.08.003.
Uri V., Varik M., Aosaar J., Kanal A., Kukumägi M., Lõhmus K. (2012). Biomass production and carbon sequestration in a fertile silver birch (Betula pendula Roth) forest chronosequence. Forest Ecology and Management 267: 117–126. https://doi.org/10.1016/j.foreco.2011.11.033.
Vacek S. (1991). Porostotvorné schopnosti břízy a jeřábu pod vlivem imisí. [Forest stand establishment capabilities of birch and wild service tree under the influence of immissions]. Zprávy lesnického výzkumu 3: 19–23. [In Czech with English summary].
Varik M., Aosaar J., Uri V. (2009). Biomass production and carbon accumulation in silver birch stands in Oxalis site type. Forestry Studies / Metsanduslikud Uurimused 51: 5–16. https://doi.org/10.2478/v10132-011-0073-y.
Weger J. (2003). Pěstování rychle rostoucích dřevin r. r. d. ve velmi krátkém obmýtí na zemědělské půdě pro produkci biomasy na energetické a průmyslové využití. [Growing of fast-growing woody plants in a very short time on agricultural land for the production of biomass for energy and industrial use]. In: Weger J., Havlíčková K. a kol. 2003. Biomasa. Obnovitelný zdroj energie v krajině. [Biomass. Renewable energy source in the landscape]. VÚKOZ, Průhonice. p. 21–35. [In Czech].
Weger J., Jiránek J. (2003). Specifikace stávajícího potenciálu biomasy k energetickému využití v ČR. [Specification of the existing biomass potential for energy use in the Czech Republic]. In: Weger J., Havličkova K. a kol. 2003. Biomasa. Obnovitelný zdroj energie v krajině. [Biomass. Renewable energy source in the landscape]. VÚKOZ, Průhonice. p. 4–11. [In Czech].
Zakopal V. (1958). Vliv březových porostů na půdní stav holin v oblasti Křivokládské. [The influence of birch stand to soil properties of clear cut area in Křivoklát]. Lesnictví 10: 877–896. [In Czech].
Zàlïtis T., Zàlïtis P. (2007). Growth of young stands of silver birch (Betula pendula Roth.) depending on pre-commercial thinning intensity. Baltic Forestry 13(1): 61–67.
Zasada M., Bijak S., Bronisz K., Bronisz A., Gawęda T. (2014). Biomass dynamics in young silver birch stands on post-agricultural lands in central Poland. Drewno 57(192): 29–39. https://doi.org/10.12841/wood.1644-3985.S07.02.
Zeide B. (1993). Analysis of growth equations. Forest Science 39(3): 594–616. https://doi.org/10.1111/j.1461-0248.2006.00883.x.
Zianis D., Muukkonen P., Mäkipää R., Mencuccini M. (2005). Biomass and stem volume equations for tree species in Europe. Silva Fennica Monographs 4. 63 p.
Total of 61 references.
Send to email
