Rebecka Mc Carthy  ,
Per Magnus Ekö,
Lars Rytter
,
Per Magnus Ekö,
Lars Rytter
Reliability of stump sprouting as a regeneration method for poplars: clonal behavior in survival, sprout straightness and growth
Mc Carthy R., Ekö P. M., Rytter L. (2014). Reliability of stump sprouting as a regeneration method for poplars: clonal behavior in survival, sprout straightness and growth. Silva Fennica vol. 48 no. 3 article id 1126. https://doi.org/10.14214/sf.1126
Highlights
- Clonal differences were found in stump sprout survival
- Clonal differences occurred in sprout straightness
- It is uncertain if the ability to produce a high number of stump sprouts is beneficial.
Abstract
The interest in poplars (Populus spp.) has increased globally as a result of the current incentive to expand the use of biomass for energy. So far, knowledge about poplar stump sprouting is generally poor. The objective of this study was to investigate poplar stump sprouting as regeneration method in harvested plantations. A 19-year-old poplar clonal test, including 23 different clones, on former arable land was harvested in the winter. After one year, stumps were assessed for sprout survival, sprout straightness, number of sprouts, sprout height and basal diameter. Differences between clones were found for survival and sprout straightness. Clonal differences were also found in mean size of sprouts (basal diameter and height). However, no differences were found between clones when comparing size of the tallest sprout or number of sprouts produced on stumps. This study indicates that stump sprouts from various poplar clones differ in survival, sprout straightness and mean growth, which should be taken into account when planning for the next tree generation. The study needs to be complemented with other studies to better assess sprouting characteristics for different clones.
Keywords
Populus spp.;
coppice regeneration;
vegetative regeneration;
short-rotation forestry
-
Mc Carthy,
Forestry Research Institute of Sweden, Skogforsk, Ekebo 2250, SE-268 90 Svalöv, Sweden & Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden
E-mail
rebecka.mccarthy@skogforsk.se
- Ekö, Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail Per.Magnus.Eko@slu.se
- Rytter, Forestry Research Institute of Sweden, Skogforsk, Ekebo 2250, SE-268 90 Svalöv, Sweden E-mail lars.rytter@skogforsk.se
Received 27 February 2014 Accepted 25 June 2014 Published 29 August 2014
Views 92129
Available at https://doi.org/10.14214/sf.1126 | Download PDF

1 Introduction
Poplar species and hybrids (Populus spp.) are generally fast-growing and have been improved by selection and breeding for productivity and vitality. Globally they are used for veneer, timber, pulp and biomass, as well as for a range of environmental services (Henry 1991; Adler 1996). Interest in poplar management is growing due to fast growth, high biomass yields potential, and high potential for carbon sequestration (Rytter 2012). Thus, the species could contribute to the vision in the Nordic countries to become carbon neutral by 2050 (Nordenergi 2010; OECD/IEA 2013). Even though the interest is large, the area managed for poplar is still relatively small in Sweden (Rytter et al. 2011a). Poplars are mostly located on agriculture land in southern Sweden, but after a severe storm in 2005 the interest has increased also for forest land, especially when reestablishing wind thrown areas. However, knowledge of cultivation and establishment remains limited in northern Europe. Poplars are usually planted as rooted cuttings in Sweden, while cuttings are commonly used in other regions. Reestablishment of poplar plantations through replanting is common in central and southern Europe. In Sweden, the stump sprouting (hereafter referred to as sprouting) in the second generation is also of high interest. This is partly explained by the high price of plant material and need of fencing. Fencing is generally recommended in Sweden (Rytter et al. 2011b) due to high populations of moose and deer. If the chosen clones provide massive coppice regeneration, fencing may not be needed, since there are many sprouts available for future crop tree selection. If coppice regeneration is preferred the parent trees are usually cut during the dormant season to achieve a viable regeneration through sprouts (DeBell and Alford 1972; Kramer and Kozlowski 1979; Strong and Zavitkovski 1983). The sprouting ability may also be affected by species/hybrid, tree size as well as environmental conditions (Blake 1983; Stanturf et al. 2001).
Knowledge about regeneration through sprouting is generally poor for clones used in northern Europe. So far, studies have mostly been made on successful sites, with satisfying coppicing regeneration dominated by one clone. Thus, potential problems may not yet been detected. Problems such as unstable sprouts originating from the upper part of the stump (Johansson and Hjelm 2012) and differences in clonal performance have been recorded (Ceulemans et al. 1996; Laureysens et al. 2003).
The objective of this study was to investigate the ability of 23 poplar clones to produce useful sprouts for regeneration after harvest. One-year-old sprouts were assessed in terms of survival, straightness, number of sprouts per stump and growth.
2 Materials and methods
2.1 Experimental design
This study was based on a former clone test located in southern Sweden (55°59´N, 12°59´E, altitude 75 m) on an abandoned agriculture field of 0.7 ha. At establishment, the test site included 120 clones that were randomized with one to three individuals in each of eight blocks planted at a density of 2.5 m × 2.5 m. The soil was an agriculture soil of clayey moraine origin (Stener 2004). The previous stand was thinned two times and exposed to storm once. The final stand consisted of 83 clones and 276 evenly distributed stems per ha. The stand was felled in winter 2010/2011 at the age of 19, with a single-grip harvester which was considered gentle on the remaining stumps. At least three stumps per clone, i.e. living trees at the final felling, were required in the study. This resulted in a total of 23 clones, where P. trichocarpa dominated the study with 16 clones (Table 1). Each clone was thus represented by three to 10 stumps. Because of the limited number of stumps per clone, this study was statistically regarded as one randomized site. The clones in the study were well distributed in the site. Eight clones were among the 12 clones that are commercially used and seven were candidates for becoming commercial. Additionally, eight other clones were included for testing. In the previous stand, harvested trees had a diameter at breast height (dbh) ranging from 13 to 49 cm (Table 1).
| Table 1. Clones included in the study (origin from a clone test (Stener 2004)). | |||||
| Clone ID a) | Taxon | Dbh d) (cm) | No. of stumps | Additional information | |
| Co1 | (5) | P. maximowiczii b) × P. trichocarpa b) | 40.1 | 7 | USA, Commercial name ‘Androscoggin’ |
| Co2 | (4) | P. maximowiczii × P. nigra b) | 36.5 | 10 | USA, Commercial name ‘Rochester’ |
| Co3 | (113) | P. maximowiczii × P. trichocarpa | 46.3 | 4 | USA |
| Co4 | (8) | Balsam type c) | 36.5 | 4 | Unknown origin |
| Co5 | (122) | P. trichocarpa | 35 | 4 | Br. Columbia, Canada |
| Co6 | (108) | P. trichocarpa | 32 | 4 | Br. Columbia, Canada |
| Co7 | (97) | P. trichocarpa | 29.7 | 3 | Alaska, USA |
| Co8 | (38) | Balsam type c) | 38.8 | 4 | Unknown origin |
| Ca1 | (87) | P. trichocarpa | 36.7 | 3 | Washington, USA |
| Ca2 | (37) | P. deltoides b) × P. nigra | 35.3 | 3 | Unknown origin |
| Ca3 | (67) | P. trichocarpa | 30.4 | 5 | Br. Columbia, Canada |
| Ca4 | (103) | P. trichocarpa | 31 | 4 | Br. Columbia, Canada |
| Ca5 | (50) | P. trichocarpa | 29.4 | 5 | Br. Columbia, Canada |
| Ca6 | (111) | P. trichocarpa | 34.8 | 4 | Br. Columbia, Canada |
| Ca7 | (92) | P. trichocarpa | 29 | 4 | Br. Columbia, Canada |
| 1 | (51) | P. trichocarpa | 26.8 | 4 | Oregon, USA |
| 2 | (2) | P. deltoides × P. trichocarpa | 38.3 | 4 | Wageningen, Netherlands, Commercial name ‘Barn’ |
| 3 | (45) | P. trichocarpa | 26.3 | 4 | Washington, USA |
| 4 | (104) | P. trichocarpa | 25.8 | 4 | Washington, USA |
| 5 | (54) | P. trichocarpa | 25.6 | 5 | Oregon, USA |
| 6 | (66) | P. trichocarpa | 23.8 | 5 | Br. Columbia, Canada |
| 7 | (96) | P. trichocarpa | 29.3 | 4 | Oregon, USA |
| 8 | (121) | P. trichocarpa | 19.3 | 4 | Br. Columbia, Canada |
| a) Commercial clones start with clone ID ‘Co’, candidates start with ‘Ca’ and additional clones are numbered in the bottom. In parentheses Skogforsk’s ID is given. b) P. maximowiczii Henry, P. trichocarpa Torr. & Gary, P. deltoides Marsh., P. nigra L. c) Unknown clones of P. trichocarpa, P. balsamifera L. or hybrids of these two species. d) Dbh refers to the felled trees. | |||||
2.2 Measurements
Sprouting ability of clones was inventoried before flushing in spring 2012, one year after final felling. Sprouts were counted on each stump, including those visibly attached to buttresses. The tallest sprout and 10 randomly selected sprouts from each stump were measured for height and basal diameter. All sprouts were selected in case there were fewer than 10 sprouts. The diameter was measured just above the enlarged part at the base of the sprout. Height was measured from the base to the highest living bud. All stumps were assessed for sprout straightness in three classes; 1) straight, 2) crooked, and 3) very crooked. In summer 2012, a re-inventory of stump survival (i.e. stumps supporting live sprouts) was carried out. This was done in two classes; 1) dead, or 2) alive.
2.3 Analyses
Stump survival data was binomial and analyzed by a generalized linear mixed model with a logit link and clone as a random effect. Stump survival in clone groups (commercial, candidates and additional clones) were analyzed with a generalized linear model with a logit link, contrasts were made between clone groups. Clones having less than two living stumps were after the survival analysis excluded from the continuing analyses. To study if any differences in straightness among clones could be detected, the straightness was analyzed through the procedure PROC FREQ (SAS Institute, Cary, NC, USA), and using Fisher’s exact test (Fisher 1922). The mean diameter and height of sampled and tallest sprouts, as well as the number of sprouts, were analyzed by a linear mixed model with clone as a random effect. Significance for all tests was determined at an alpha level of 0.05. The model used was:
![]()
where yij is the observed mean diameter/height/number of sprouts for clone i and stump j, µ is the estimated over all mean, αi is the random clonal effect and εij is an error term with normal distribution, constant variance and expectation 0.
The clone groups were analyzed by an ANOVA for diameter and height (both sampled and tallest sprouts), as well as for the number of sprouts. The model used for the ANOVA was:
![]()
where ygj is the observed mean diameter/height/number of sprouts for clone group g and stump j, µ is the estimated over all mean, αg is fixed effect of the clone group and εgj is an error term with normal distribution, constant variance and expectation 0.
3 Results
3.1 Survival
The relative number of stump survival, i.e. survival frequency, differed between clones (P < 0.0001). At the re-inventory the overall clone mean was 70 %. Nearly 50 % of the clones had 100 % stump survival, while 20 % had less than 50 % survival (Fig. 1). The candidate clones had six out of seven clones with 100 % survival, which was higher than both commercial and additional clones (P = 0.004). The commercial clones did not differ in stump survival from the additional clones (P = 0.55). Furthermore, no pattern of the spatial distribution of dead stumps could be detected over the site (data not shown).
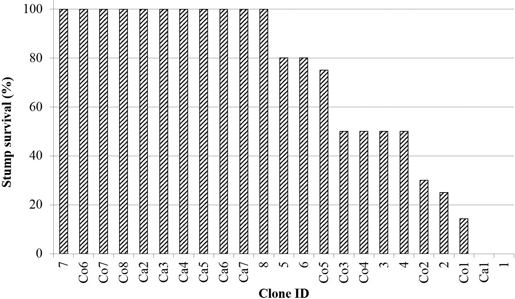
Fig. 1. The relative frequency of stump survival (meaning stumps supporting live sprouts). For clone information, see Table 1.
3.2 Straightness
There was a difference between clones regarding sprout straightness (P = 0.0013). Considering clones where more than one stump was alive, three of the clones had stumps with severely crooked sprouts (mean of classes > 2.5), while seven clones had straight sprouts (mean of classes < 1.5) (Fig. 2). Of all stumps, 35 % were classified as having straight sprouts, 49 % as having crooked, and 16 % as having severely crooked sprouts. Both commercial and candidate clones had stumps represented in all three straightness classes.
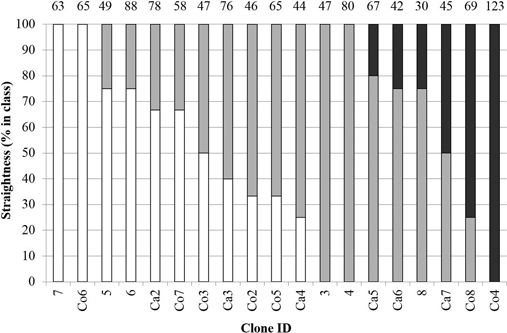
Fig. 2. Sprout straightness (white = straight (class 1), grey = crooked (class 2), black = severely crooked (class 3)). Numbers on top represent the mean number of sprouts per stump. For clone information, see Table 1.
3.3 Growth
There were no significant differences in number of sprouts per stump produced by the different clones (P = 0.6049) (Fig. 3). Overall, clones produced between 6 and 137 sprouts, with a considerable within clone variation. The overall clone mean was 62 sprouts per stump.
The mean diameter and mean height of sampled sprouts showed differences between clones (P = 0.0071 and P = 0.0293). The diameters ranged from 2 to 31 mm with a clone mean of 8.6 mm, and the heights ranged from 12 to 275 cm with a clone mean of 105 cm. No clonal differences were found among the diameter and height of the tallest sprouts (P = 0.3268 and P = 0.1786). The tallest sprouts ranged between 4 and 31 mm in diameter with clone mean 17.1 mm, and the height ranged between 91 and 275 cm with clone mean of 175 cm.
The three clone groups, commercial, candidate and additional clones, did not show any differences in growth characteristics.
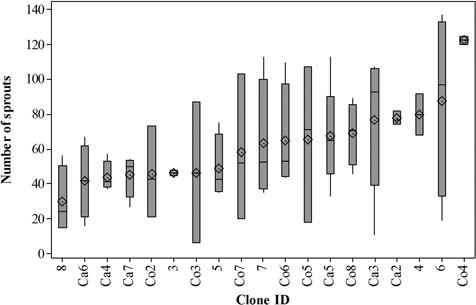
Fig. 3. Quartile range box of number of sprouts per stump. Vertical lines are the upper and lower whiskers, representing the boarders of the data. Top line in the box is the third quartile, middle line is the median, and bottom line is the first quartile. Mean value is shown by a diamond. For clone information, see Table 1.
4 Discussion
This study showed differences among poplar clones in stump sprouting ability and sprout straightness. Two P. trichocarpa clones, one commercial and one additional clone, showed 100 % survival and 100 % straightness. There were also other clones performing well in survival and straightness, thus suitable for regeneration through sprouts. On the other hand, some clones should be avoided if coppice regeneration is intended. The candidate clones showed a higher survival than the commercial and additional clones, which is a desired character for stump sprout regeneration.
Other studies have also shown that poplar clones can perform very differently in sprouting regimes (Herve and Ceulemans 1996; Laureysens et al. 2003). Johansson and Hjelm (2012) looked at sprouting capacity and found survival rates between 57 and 100 % among stumps in different sites in Sweden. However, their material was dominated by one clone. Laureysens et al. (2003), on the other hand, reported a stump survival that averaged 35 to 93 % after four years among 17 clones in Belgium. The overall mean survival found in this study, 70 %, is in agreement with the studies mentioned above. It has also been stated by Zasada et al. (2001) that coppice regeneration is not always reliable for e.g. P. deltoides, P. trichocarpa and P. balsamifera L. This study supports the statement since it showed clonal differences in stump survival.
The stem diameter of the mother tree (at breast height) was added as a covariate in the survival model, since this possibly could have an influence. In this study, the diameter of the harvested trees did not show any significance for survival and the covariate was therefore removed from the final model.
Focus is rarely on sprouting performance when selecting clones. Instead, clones are usually selected for growth rate, resistances against pests and diseases, stability, and desirable wood properties (Bisoffi and Gullberg 1996; Stener 2004). The difference in survival in this study might depend on the criteria used for selection. Some clones produced no or a limited number of sprouts, but performed well in the first generation according to previous measures (Table 1). It would be optimal to find clones having a fast growth and producing high timber quality, but also having good sprouting ability, since economically it could be favorable to use coppice regeneration rather than replanting. The advantages would be reduced establishment costs due to an initially fast growth (Zasada et al. 2001). This study showed that suitable clones for regeneration with sprouts are included in the Swedish clones, although care must be taken to select the useful ones. It should also be noted that by using stump sprout regeneration there is a risk that new selected clones, with improved growth and vitality, will not be introduced at an optimal rate, but this can be taken care of by using both planting and natural regeneration in future afforestation of poplar.
It should be noted that the occurrence of root suckers was not investigated in this study. Balsam poplars (P. balsamifera) may regenerate through root suckers (Zasada et al. 2001), but a visual inspection revealed only few root suckers in the study area.
The reason to include all living sprouts in the study, also the small ones, is the difficulty to forecast survival and development in an early stage. The measurements took place one season after harvest. According to Ceulemans and Deraedt (1999) the expected survival rate of sprouts is less than 10 % in 3–4 years, or according to Johansson and Hjelm (2012) approximately 3 sprouts per stump will remain after 7 years.
The number of sprouts found after harvest was higher in this study than generally reported in the literature (Strong and Zavitkovski 1983; Ceulemans and Deraedt 1999; Laureysens et al. 2003). A major reason to the difference is probably the age of the harvested trees and thereby the size of the stumps, and that all small sprouts may not have been counted in all studies. The harvested stand in our study was 19 years, but in most other studies not more than 5 years. Johansson and Hjelm (2012) investigated sprouting in older poplar stands containing a few clones and here the initial sprout number (after 1 year) was closer to our result, although somewhat lower. Differences in plant material, i.e. clones, may influence the sprout numbers seen in this study.
It has been observed that sprouts originating from the upper part of the stump may be less stable due to breakage from the stump (Johansson and Hjelm 2012). It was also observed at the study site that sprouts could be unstable and break, and most of these sprouts were located in the upper part of the stump. It is also likely that crooked sprouts could later cause breakage and instability.
The sampled sprouts showed differences in diameter and height among clones, although the tallest sprouts did not since the variance between the tallest sprouts was large. It is likely that a restricted number of dominant or co-dominant sprouts will outcompete other sprouts in the long run, resulting in few sprouts per stump after a period as forecasted by Ceulemans and Deraedt (1999) and Johansson and Hjelm (2012). The tallest sprouts are the most likely candidates for being future crop trees. If the number of sprouts per stump is high, there is a possibility that the development of the dominant sprouts is disfavored by competition. It could therefore be of interest to single out sprouts on stumps at a quite early stage in order to favor diameter development (Stanturf et al. 2001). The timing should of course be weighted against the risk of browsing damages. Furthermore, Zasada et al. (2001) claimed that a certain amount of sprouts are needed to keep the parent root system alive.
The number of sprouts was added as a covariate to the analyses of sprout diameter and height in model 1, since this may lead to competition and affect the growth. The covariate did not show any significance in any of these analyses and was therefore removed from the final model. It is possible that the covariate needs more time with competition to give an effect.
This study was carried out in one site with varying replicates of stumps per clone. It was an imbalance among clones as a result of previous commercial thinnings. This leads to a limited base from which to draw general conclusions. Therefore, the purpose in this study was mainly to discuss if we can rely on satisfying regenerations from sprouts in the second generation and promote further research. Further research is needed since knowledge of poplar sprouting is generally poor, not least in northern Europe. Thus, the topic is of interest for many countries where poplars are cultivated.
5 Conclusions
In this study we found significant differences among clones in stump survival. Also the ability to produce straight sprouts differed. For practical implications it is not obvious that an ability to produce a high number of stump sprouts is beneficial for future growth. Internal competition will probably be high and reduce growth of surviving sprouts.
Acknowledgements
This study was financially supported by Nordic Energy Research and is part of their project Wood based energy systems from Nordic Forests (ENERWOODS). We would like to address special thanks to Karin Johansson and Magnus Löf for helpful comments on the manuscript, as well as to Johan Malm for kind assistance during fieldwork.
References
Adler T. (1996). Botanical cleanup crews: using plants to tackle polluted water and soil. Science News 150: 42–43.
Bisoffi S., Gullberg U. (1996). Poplar breeding and selection strategies. In: Stettler R., Bradshaw H., Heilman P. (eds.). Biology of Populus and its implications for management and conservation. NRC Research Press. p. 139–158.
Blake T. (1983). Coppice systems for short-rotation intensive forestry: the influence of cultural, seasonal and plant factors. Australian Forest Research 13: 279–291.
Ceulemans R., Deraedt W. (1999). Production physiology and growth potential of poplars under short-rotation forestry culture. Forest Ecology and Management 121: 9–23. http://dx.doi.org/10.1016/S0378-1127(98)00564-7.
Ceulemans R., McDonald A., Pereira J. (1996). A comparison among eucalypt, poplar and willow characteristics with particular reference to a coppice, growth-modelling approach. Biomass and Bioenergy 11: 215–231. http://dx.doi.org/10.1016/0961-9534(96)00035-9.
DeBell D.S., Alford L. (1972). Sprouting characteristics and cutting practices evaluated for cottonwood. Tree Planters’ Notes 23: 1–3.
Fisher R.A. (1922). On the interpretation of χ2 from contingency tables, and the calculation of P. Journal of the Royal Statistical Society: 87–94.
Henry C.L. (1991). Nitrogen dynamics of pulp and paper sludge amendment to forest soils. Water Science and Technology 24: 417–425.
Herve C., Ceulemans R. (1996). Short-rotation coppiced vs non-coppiced poplar: a comparative study at two different field sites. Biomass and Bioenergy 11: 139–150. http://dx.doi.org/10.1016/0961-9534(96)00028-1.
Johansson T., Hjelm B. (2012). The sprouting capacity of 8–21-year-old poplars and some practical implications. Forests 3: 528–545. http://dx.doi.org/10.3390/f3030528.
Kramer P.J., Kozlowski T.T. (1979). Physiology of woody plants. Academic Press. 272 p.
Laureysens I., Deraedt W., Indeherberge T., Ceulemans R. (2003). Population dynamics in a 6-year old coppice culture of poplar. I. Clonal differences in stool mortality, shoot dynamics and shoot diameter distribution in relation to biomass production. Biomass and Bioenergy 24: 81–95. http://dx.doi.org/10.1016/S0961-9534(02)00105-8.
Nordenergi (2010). A CO2-neutral vision for the Nordic countries for 2050 – the Nordic perspective. Nordenergi Secretariat. 13 p.
OECD/IEA (2013). Nordic energy technology perspectives – pathways to a carbon neutral energy future. OECD/IEA & Nordic Energy Research. 211 p.
Rytter L., Johansson T., Karačić A., Weih M., Börjesson P., Fogdestam N., Hannerz M., Ingvarsson P., Rosenqvist H., Stener L.-G. (2011a). Investigation for a Swedish research program on the genus Populus. Arbetsrapport 733. The Forestry Research Institute of Sweden (Skogforsk). [In Swedish with English summary].
Rytter L., Stener L.-G., Övergaard R. (2011b). Cultivation of hybrid aspen and poplar. Guidance. The Forestry Research Institute of Sweden (Skogforsk). [In Swedish].
Rytter R.-M. (2012). The potential of willow and poplar plantations as carbon sinks in Sweden. Biomass and Bioenergy 36: 86–95. http://dx.doi.org/10.1016/j.biombioe.2011.10.012.
Stanturf J.A., Van Oosten C., Netzer D.A., Coleman M.D., Portwood C.J. (2001). Ecology and silviculture of poplar plantations. In: Dickmann D.I., Isebrands J.G., Eckenwalder J.E., Richardson J. (eds.). Poplar culture in North America. NRC Research Press, Ottawa. p. 153–206.
Stener L.-G. (2004). Results from clonal tests of poplar in Southern Sweden. Arbetsrapport 571. The Forestry Research Institute of Sweden (Skogforsk). [In Swedish].
Strong T.F., Zavitkovski J. (1983). Effect of harvesting season on hybrid poplar coppicing. In: Hansen E.A. (ed.). Intensive plantation culture: 12 years research. General Technical Report, NC-91. United States Department of Agriculture, Forest Service, Northern Central Forest Experimental Station, Saint Paul, Minnesota, USA. p. 54–57.
Zasada J., David A., Gilmore D., Landhausser S. (2001). Ecology and silviculture of natural stands of Populus species. In: Dickmann D.I., Isebrands J.G., Eckenwalder J.E., Richardson J. (eds.). Poplar culture in North America. NRC Research Press, Ottawa. p. 119–151.
Total of 21 references
