Eva Ring  ,
Ulf Sikström
,
Ulf Sikström
Environmental impact of mechanical site preparation on mineral soils in Sweden and Finland — a review
Ring E., Sikström U. (2024). Environmental impact of mechanical site preparation on mineral soils in Sweden and Finland — a review. Silva Fennica vol. 58 no. 1 article id 23056. https://doi.org/10.14214/sf.23056
Highlights
- Mechanical site preparation is carried out on large areas, but limited research on its environmental impact has been undertaken
- It affects nitrogen and carbon cycling over the first few years, and has a minor initial impact on CO2, CH4 and N2O fluxes
- It increases tree carbon stores and possibly ecosystem carbon stores
- Reducing its soil disturbance intensity is warranted.
Abstract
Mechanical site preparation (MSP) is deliberate soil disturbance which is undertaken to improve the conditions for forest regeneration. Disc trenching and mounding are the dominant MSP practices currently used in Sweden and Finland. In this paper, the impacts of MSP on the soil, water quality, greenhouse gas (GHG) emissions and ground vegetation of mineral soil sites in Sweden and Finland are reviewed. The practices considered are patch scarification, mounding, inverting, disc trenching, and ploughing, which together represent a wide range of soil disturbance intensity. The environmental effects of MSP in this region have not been studied extensively. The environmental impact of MSP derives from the process of creating microsites which involves horizontal and/or vertical redistribution of soil and soil mixing. This typically affects decomposition, element circulation and leaching, vegetation coverage and uptake of nutrients and water, and possibly erosion and sediment exports. Following disc trenching or mounding the effects on GHG emissions appear to be minor over the first two years. For a few years after disc trenching concentrations in soil water collected below ridges are higher than that below furrows for some elements (e.g., NO3-, NH4+, Mg2+, and total or dissolved organic C). The physical and chemical effects of ploughing remain detectable for several decades. There is little evidence about how the effects of forestry activities in upland areas on soil-water chemistry are transferred to adjacent surface water bodies, including what role streamside discharge areas play. MSP increases the tree biomass C store and may increase the total ecosystem C store. The impact of MSP on the cover and abundance of ground vegetation species depends on the composition of the original plant community, MSP intensity, and the establishment rate of different species. Species cover generally seems to decline for late succession understory species, while pioneer and ruderal species can benefit from the microsites created. Areas containing lichens which are used for reindeer forage require special consideration. More research is needed on the environmental effects of MSP, particularly regarding its long-term effects. Further efforts should be made to develop efficient site-preparation practices which better balance the disturbance intensity with what is needed for successful regeneration.
Keywords
carbon;
nitrogen;
soil disturbance;
vegetation;
water;
chemistry;
greenhouse gas
-
Ring,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83, Uppsala, Sweden
 https://orcid.org/0000-0002-8962-9811
E-mail
eva.ring@skogforsk.se
https://orcid.org/0000-0002-8962-9811
E-mail
eva.ring@skogforsk.se
- Sikström, Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83, Uppsala, Sweden E-mail ulf.sikstrom@skogforsk.se
Received 20 September 2023 Accepted 7 February 2024 Published 28 February 2024
Views 145488
Available at https://doi.org/10.14214/sf.23056 | Download PDF

1 Introduction
Mechanical site preparation (MSP) is a common practice undertaken during forest regeneration, usually during snow-free conditions without soil frost, to improve its outcomes by providing favourable planting spots for planted seedlings and good conditions for sown and naturally regenerated seedlings to emerge. During the 2010s, roughly 140 000 ha and 100 000 ha of forest land were subjected to MSP annually, in Sweden and Finland, respectively (Fig. 1). Various types of machinery are used to execute MSP. Usually, a patch scarifier, mounder, or disc trencher is attached to a continuously advancing base machine. Excavators equipped with different types of buckets are also used to create mounds or invert the soil.
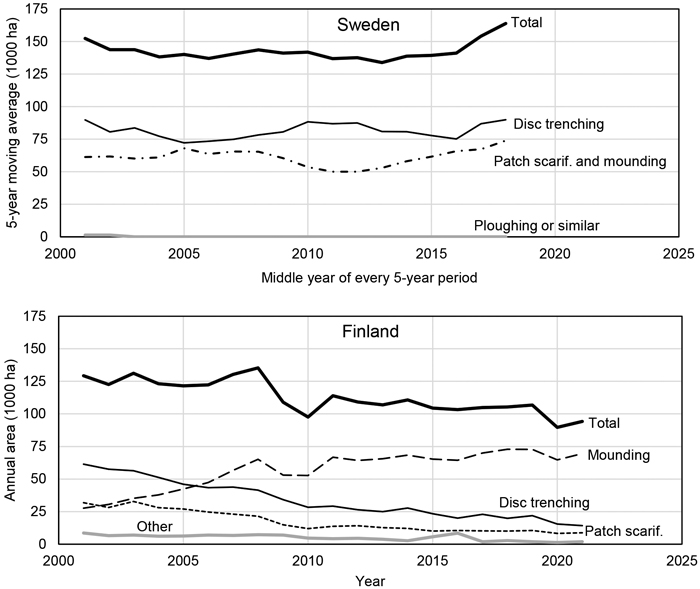
Fig. 1. Annual area subjected to various forms of mechanical site preparation in Sweden (top) and Finland (bottom). The data for Sweden are 5-year moving averages plotted against the middle year of each 5-year period, hence, the first and last data points represent 1999–2003 and 2016–2020, respectively. For Finland, annual areas for the period 2001 to 2021 are presented. The data were provided by the Swedish National Forest Inventory (2022) and Luke Statistics database (http://statdb.luke.fi/PXWeb/pxweb/en/LUKE/LUKE__04%20Metsa__06%20Metsavarat/, accessed on 13 December 2022). “Patch scarif.” refers to patch scarification.
Patch scarification is mainly used on dry sites with fairly low fertility or on stony sites where other MSP practices cannot be performed; mounding is generally practiced on mesic to moist sites, where elevated planting spots are beneficial for seedling survival; soil inversion (henceforth also referred to as inverting) can be used on most sites, except where soils are very dry (to avoid cutting off capillary water supply) or very moist (where elevated planting spots are needed); disc trenching can be used on all types of mineral soils, including stony sites, but is unsuitable for moist sites; and ploughing has mainly been used on moist soils with low nutrient availability (often caused by low soil temperatures) (Sikström et al. 2020). Ploughing is prohibited in Sweden (Swedish Forest Agency 2022) but is performed on a small proportion of the total area prepared in Finland (https://statdb.luke.fi/PXWeb/pxweb/sv/LUKE/). The Finnish FSC standard restricts the maximum plough depth to 25 cm (FSC-STD-FIN-(Ver1-1)-2006 Finland natural forests EN, https://fi.fsc.org/en/node/28015, accessed on 14 June, 2023). Over the last decade, the most common MSP practices by area in Sweden and Finland are disc trenching and mounding, respectively (Fig. 1).
For planted conifer seedlings, MSP generally increases survival rates by 15–20 percent units and the height 10–15 years after planting by 10–25%, compared with non-prepared sites (Sikström et al. 2020). In the longer term, i.e., after about 30 years, the relative difference in stem volume can be even greater (+20–50%) (Hjelm et al. 2019). MSP promotes the emergence and establishment of seedlings from seeding and natural regeneration (Lehto 1956; Hagner 1962; Skoklefald 1995; Wennström et al. 1999). However, with natural regeneration the timing between MSP and a rich seed fall can be crucial to the emergence of a sufficient number of seedlings (Karlsson and Örlander 2000).
The basic principle of MSP is to remove the organic top-soil layer and expose bare mineral soil, or to create a mixture of organic matter and mineral soil. Regeneration is often more successful on elevated spots. As summarised in the review by Sikström et al. (2020), bare mineral soil is generally more favourable for seedling survival than an organic top soil layer, and often the same is true for initial growth, due to higher soil temperature, more stable soil moisture conditions, less vegetation competition, reduced risk of frost damage, and, in areas with occurrence of pine weevil (Hylobius abietis L.), significantly less feeding on seedlings by this beetle. An admixture of organic matter can be beneficial for growth, particularly for some tree species such as Norway spruce (Picea abies (L.) Karst.) (Nilsson et al. 2019). MSP also affects the soil structure by loosening the soil, which can improve root growth (Örlander et al. 1990), and mixing organic matter with mineral soil can stimulate mineralisation and nutrient availability for seedlings (McMinn 1985; Nordborg et al. 2003).
MSP affects several of the physical, chemical, and biological properties of, and processes in, the upper soil. This review provides an overview of the environmental impacts of MSP undertaken during forest regeneration on mineral soil sites in Sweden and Finland. The topics reviewed encompass physical soil disturbance, nitrogen (N), carbon (C), greenhouse gas (GHG) emissions, other chemical variables, and ground vegetation. The aim was to cover a broad spectrum of environmental impacts in a relatively well-defined context. Thus, focus is on Sweden and Finland with many similarities regarding climate, forest types, ground vegetation species, soils, and forest management. Furthermore, MSP is a common measure, performed using the same practices, in both countries. The MSP practices covered are mainly patch scarification, mounding, inverting, disc trenching, and ploughing. Gaps in knowledge are identified, and suggestions on how to reduce undesirable impacts are presented.
2 Materials and methods
A search for peer-reviewed scientific articles on MSP was performed on 9 December 2022 in the databases Web of Science Core Collection, CABI: CAB Abstracts, Scopus, and GreenFILE. Three blocks of search terms were combined to identify articles dealing with relevant 1) MSP practices, 2) forests, and 3) vegetation zones (using the Boolean operator “OR” within blocks and “AND” among blocks). For MSP practices, the search terms used were site prep* (* meaning truncation), soil scarif*, patch scarif*, patching*, disc trench*, scalping, plough*, and harrow*. For forests, the search terms were forest*, forest regen*, clearcut*, harvest*, cutblock* and logging site*, and for vegetation zones the terms boreal, hemi boreal, and temperate were used. Alternate spellings of these words were included where appropriate. These terms were searched for in the title, abstract, or keywords and no limitation on the publication year was applied. In addition, searches were performed using Google Scholar and articles were obtained from the authors’ own literature collections, citations, and reference lists. The articles selected from these searches described the environmental effects of MSP in conifer-dominated forests on mineral soil sites in Sweden and Finland, based on studies which generally included a harvested area subject to MSP treatment and a non-prepared control.
The microsites created by MSP are referred to as “patch” (patch scarification), “mound” (mounding), and “inverted patch” (inverting) for patch-based practices, and the continuous lines created by disc trenching are referred to as “furrows” and “ridges”, with soil from the furrows forming adjacent ridges. The corresponding terms for ploughing are “trenches” and “tilts”, respectively.
3 Studies performed in Sweden and Finland
In total, 44 articles on the environmental impacts of MSP in Sweden and Finland were reviewed (Table 1), reporting investigations performed at 51 sites and findings from synoptic field studies (Fig. 2). Some of the sites were used to study multiple environmental factors, most notably, Kangasvaara (Mannerkoski et al. 2005; Palviainen et al. 2007; Piirainen et al. 2007, 2009, 2015), and Karkkila (K1) and Kuorevesi (K2) (Tanskanen and Ilvesniemi 2004, 2007; Tanskanen et al. 2004; Lindroos et al. 2016; Lindroos and Ilvesniemi 2023) in Finland, and Hagfors (Johansson et al. 2013; Ring et al. 2013; Rappe George et al. 2017) and Billingsjön (Ring 1996; Nohrstedt 2000; Ring et al. 2018) in Sweden.
| Table 1. Articles and number of articles reporting impacts of mechanical site preparation (MSP) on various environmental variables, based on studies performed in Sweden and Finland, sorted by MSP practice. The MSP practice “Other” refers to MSP performed experimentally by hand or excavator, or by the MSP practice HuMinMix (see section 9). | ||||||
| Environmental variable | MSP practice | |||||
| Patch scarification | Mounding | Inverting | Disc trenching | Ploughing | Other | |
| Nitrogen1 | 2 | 4 | 1 | 10 | 5 | 7 |
| (Nordborg et al. 2006; Egnell et al. 2015) | (Johansson 1994; Smolander et al. 2000; Smolander and Heiskanen 2007; Nieminen et al. 2012) | (Smolander and Heiskanen 2007) | (Johansson 1994; Lundmark-Thelin and Johansson 1997; Mannerkoski et al. 2005; Palviainen et al. 2007; Piirainen et al. 2007; Johansson et al. 2013; Närhi et al. 2013; Ring et al. 2013; Piirainen et al. 2015; Rappe George et al. 2017) | (Johansson 1994; Kubin 1995, 1998; Tanskanen and Ilvesniemi 2004; Sutinen et al. 2019) | (Ring 1996; Örlander et al. 1996; Nohrstedt 2000; Nordborg et al. 2006; Egnell et al. 2015; Mjöfors et al. 2015; Ring et al. 2018) | |
| Carbon and/or biomass1 | 2 | 4 | 1 | 10 | 6 | 4 |
| (Nordborg et al. 2006; Egnell et al. 2015) | (Johansson 1994; Smolander et al. 2000; Smolander and Heiskanen 2007; Mjöfors et al. 2017) | (Smolander and Heiskanen 2007) | (Johansson 1994; Lundmark-Thelin and Johansson 1997; Mannerkoski et al. 2005; Palviainen et al. 2007; Piirainen et al. 2007; Johansson et al. 2013; Närhi et al. 2013; Ring et al. 2013; Piirainen et al. 2015; Mjöfors et al. 2017) | (Johansson 1994; Tanskanen and Ilvesniemi 2004; Tanskanen et al. 2004; Tanskanen and Ilvesniemi 2007; Mjöfors et al. 2017; Sutinen et al. 2019) | (Örlander et al. 1996; Nordborg et al. 2006; Egnell et al. 2015; Mjöfors et al. 2015) | |
| GHG emission | 2 | 2 | 0 | 3 | 0 | 2 |
| (Strömgren and Mjöfors 2012; Strömgren et al. 2017) | (Strömgren et al. 2012, 2016) | (Strömgren and Mjöfors 2012; Strömgren et al. 2016, 2017) | (Pumpanen et al. 2004; Mjöfors et al. 2015) | |||
| Other chemical variables2 | 0 | 4 | 1 | 9 | 9 | 1 |
| (Johansson 1994; Smolander et al. 2000; Smolander and Heiskanen 2007; Nieminen et al. 2012) | (Smolander and Heiskanen 2007) | (Johansson 1994; Lundmark-Thelin and Johansson 1997; Mannerkoski et al. 2005; Palviainen et al. 2007; Piirainen et al. 2007, 2009; Johansson et al. 2013; Närhi et al. 2013; Ring et al. 2013) | (Johansson 1994; Kubin 1995; Tanskanen and Ilvesniemi 2004; Tanskanen et al. 2004; Sutinen et al. 2006, 2010, 2019; Lindroos et al. 2016; Lindroos and Ilvesniemi 2023) | (Örlander et al. 1996) | ||
| Ground vegetation3 | 2 | 2 | 0 | 5 | 3 | 2 |
| (Norberg et al. 1997; Tonteri et al. 2022) | (Roturier and Bergsten 2006; Tonteri et al. 2022) | (Eriksson and Raunistola 1990; Bergstedt et al. 2008; Roturier et al. 2011; Vanha-Majamaa et al. 2017; Tonteri et al. 2022) | (Eriksson and Raunistola 1990; Bergstedt et al. 2008; Tonteri et al. 2022) | (Roturier and Bergsten 2006; Roturier et al. 2011) | ||
| 1 Including decomposition, mineralisation (nitrification for N), water and soil concentrations, leaching and soil and vegetation stores. 2 Including one or several chemical variables other than N and C, for example electrical conductivity, pH, cations and anions. 3 Including vegetation coverage and species composition. | ||||||
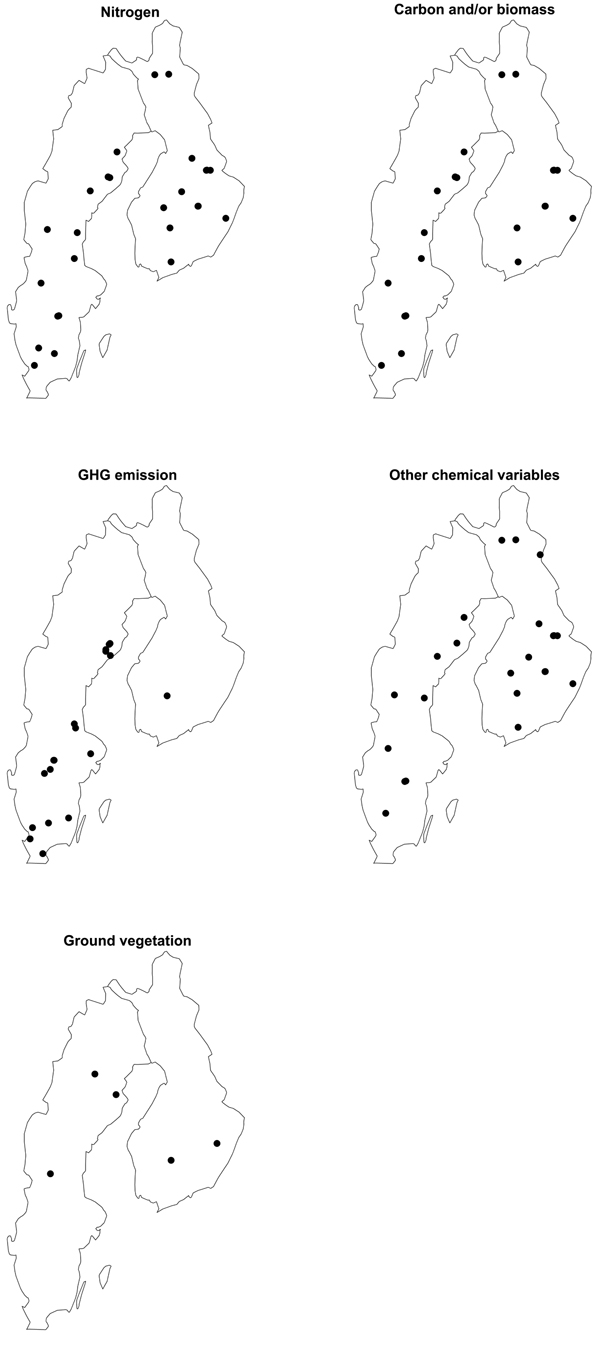
Fig. 2. Location of study sites in Sweden and Finland where environmental impact of MSP on nitrogen (n = 27), carbon and/or biomass (n = 21), greenhouse gas (GHG) emissions (n = 18), other chemical variables (n = 23) and ground vegetation species and/or cover (n = 5) have been investigated (NB: there was insufficient site information for one site where N, C and /or biomass and other chemical variables were studied). Sites surveyed in synoptic studies are not shown on the maps. The maps were produced by Per Westerfelt.
The environmental impacts of MSP on N, C and/or biomass, GHG emission, and other chemical variables were investigated at 28, 22, 18, and 24 sites, respectively. For N, C and/or biomass, and other chemical variables, these impacts encompassed any type of effect, for example on decomposition, soil-water and soil chemistry, and biomass store. Effects on the cover and/or composition of ground vegetation were studied at five sites and in three synoptic surveys. Disc trenching and ploughing have been studied most frequently (Table 1). Studies of the disturbed surface area proportion by different MSP practice were not included in Table 1, as a comprehensive compilation of these has already been presented by Sikström et al. (2020).
Some paired catchment studies, which investigate sites subject to clearcutting followed by MSP and, sometimes, subsequent drainage, have been undertaken in Sweden and Finland (Ahtiainen 1992; Rosén et al. 1996; Ahtiainen and Huttunen 1999; Porvari et al. 2003; Schelker et al. 2012; Eklöf et al. 2014; Palviainen et al. 2014). These studies show mainly the combined effect of clearcutting and MSP, and generally offer limited scope to conclusively isolate the impact of MSP, due to the interdependence between forestry measures performed within a short window of time and difficulties in separating the impact of MSP from those related to factors such as post-harvest variability in weather (cf. Schelker et al. 2012; Eklöf et al. 2014; Salmivaara et al. 2023). This review puts less focus on paired catchment studies due to the difficulty of isolating the effects of MSP from other factors.
4 Physical soil disturbance
The MSP practices reviewed here involve at least one of the following soil disturbances: 1) exposure of mineral soil by removing the organic soil-surface layer (patch scarification), or inverting the upper soil or creating mounds or ridges/tilts with adjacent pits or furrows/trenches (mounding, disc trenching, and ploughing), 2) burial of the organic soil-surface layer below mineral soil (mounding, inverting, disc trenching and ploughing), or 3) mixing organic and mineral soil (which depends on both MSP practice and execution) (Sutton 1993; Sikström et al. 2020). All of these involve redistributing soil material vertically and/or horizontally, and mixing it to some extent. In addition, driving MSP machinery across a site during the process can cause soil compaction and rutting (Cambi et al. 2015).
The physical soil disturbance caused by MSP can be described by the proportion of disturbed surface area, soil depth affected, soil volumes displaced and soil surface microtopography (also referred to as the soil surface roughness), all of which are interrelated (Fig. 3). The “disturbance intensity” caused by MSP referred to in this review is a subjective judgement of the level of disturbance taking into account both the proportion of the area disturbed and the soil depth affected.

Fig. 3. A clearcut site subjected to disc trenching on which various metrics of soil disturbance are illustrated schematically. Oriented roughness refers to a systematic variation in topography caused by, for example, tillage. Photo E. Ring.
4.1 Disturbed surface area proportion
Based on collated data from field studies, Sikström et al. (2020) proposed that 55–65% of the soil surface area is disturbed during ploughing, 45–55% during disc trenching, and 30–40% during mounding, patch scarification, and inverting. The reported disturbed proportion not only depends on the MSP practice used but also on on-site execution (for example, row spacing), equipment (for example, size of the MSP unit), and inventory method (Sikström et al. 2020). In addition, site characteristics such as soil texture, microtopography, stoniness, and soil wetness may affect the disturbed area proportion. In a study of spot mounding in Finland using continuously advancing mounders, the number of suitable planting spots created was influenced by the depth of the humus layer, stoniness, slope inclination, presence of fresh logging residue, and soil texture (Saksa et al. 2018). The total area of disturbance may extend beyond the area covered by the MSP microsites, as MSP may disrupt or move coarse roots, stumps, and stones, thereby affecting nearby areas.
4.2 Affected soil depth
The soil depth and soil layers affected by the MSP practices reviewed range from removing the organic layer by patch scarification to creating plough furrows reaching approximately half a meter into the mineral soil (0.5–0.9 m for deep ploughing) (Örlander et al. 1990). The displaced soil volume depends on the MSP practice used and number of planting spots required, but also on how the practice is executed, particularly the depth and width of the furrows and pits. The soil depth affected and the associated redistribution of soil change the conditions for soil evolution in the MSP microsites (cf. Šamonil et al. 2018). In podzols, Al and Fe accumulate in the Bs horizon during soil formation (Giesler et al. 2000). Exposing the soil from the Bs horizon by inverting it onto the soil surface through tilt ploughing was found to mobilise Al in the ploughed soil profiles at the Karkkila (K1) and Kuorevesi (K2) sites in Finland (Tanskanen et al. 2004). A soil study performed at the same sites further supported that Al mobilisation had occurred and also suggested that Al had migrated and been retained in organic matter further down in the soil (Tanskanen and Ilvesniemi 2004). The effects were most pronounced at Karkkila. Additional studies at Karkkila indicated strong weathering of the topsoil on the plough tilt during the first 17 years after ploughing, and element depletion of the top soil in the trenches (Lindroos et al. 2016; Lindroos and Ilvesniemi 2023).
4.3 Soil surface microtopography
The soil surface microtopography describes the small-scale irregularities of the soil surface (Thomsen et al. 2015). Although there is significant variability in how individual MSP practices are executed, patch scarification and inverting are likely to have the least impact of the practices reviewed on soil microtopography (cf. Örlander et al. 1990; Sikström et al. 2020). Changes to soil surface microtopography may affect the accumulation of litter and snow, erosion, and sediment transport, among other things (Neff 1980; Alcázar et al. 2002; Lindroos et al. 2016). In cold climates, consecutive snowmelt periods combined with increased snow accumulation was proposed as an explanation for the long-term effects on soil chemistry that have been measured below plough trenches (Sutinen et al. 2010; Sutinen et al. 2019). Disc trenching and ploughing cause oriented surface roughness, i.e., a systematic variation in topography that can be caused, for example, by tillage (see categorisation by Takken (2000) as presented in Thomsen et al. 2015) (Fig. 3). The oriented roughness caused by disc trenching and ploughing may increase soil erosion and sediment transport to nearby surface water bodies (Alcázar et al. 2002), with detrimental effects on aquatic life if insufficient protection zones are applied (Ahtiainen and Huttunen 1999; Österling et al. 2010).
The alteration to the soil surface microtopography caused by MSP changes over time because of erosion, subsidence, litter accumulation and revegetation (cf. Kubin and Kemppainen 1994; Palviainen et al. 2007; Valtera and Schaetzl 2017). At a site on till soil in Finland, mound heights were observed to decrease over the first 2–3 years after MSP (Heiskanen et al. 2013). At a site on till soil in Sweden, six and a half years after disc trenching, the mean height of the ridges and mean depth of the furrows tended to have decreased (Fig. 4). At two sites in northern Finland, traces of ploughing were visible on a 2 m by 2 m digital elevation model (interpolated from LiDAR data) 25 years after MSP operations (Sutinen et al. 2019).
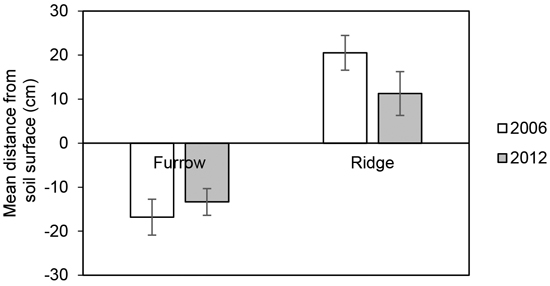
Fig. 4. Mean distance from the undisturbed soil surface to the bottom of the furrow and top of the ridge, respectively, about two months after disc trenching in 2006 (data from Ring et al. 2013) and in 2012 six and a half years later. The measurements were carried out at the Hagfors field site in Sweden (n2006 = 27, n2012 = 54). The error bars show ±1 standard deviation.
4.4 Conclusions and research needs
The physical soil disturbance intensity related to the MSP practices reviewed varies considerably, from intermittently removing small patches of the organic surface layer to creating continuous deep trenches in the mineral soil with adjacent ridges of organic matter and mineral soil. Today, however, the most intense practice, i.e., ploughing, is only carried out on small areas in Finland. Disturbance intensity relates to both the MSP practice used and its on-site execution, and possibly also site conditions such as the character of the remaining tree-root system and stoniness. The character and intensity of disturbance determines subsequent environmental impacts, on-site accessibility, and possibly people´s appreciation of the forest (Gundersen et al. 2016). In a long-term perspective that encompasses several rotations, repeated MSP at high disturbance intensity is likely to alter the upper soil substantially across the site, with possible implications for future soil evolution. Further studies of current MSP practices and their long-term effects on element mobilisation and distributions are needed.
MSP could potentially mitigate soil compaction of the upper soil caused by transportation during previous forestry operations, particularly because soil compaction due to transportation is greatest near the soil surface (Cambi et al. 2015). In contrast to logging trails used for multiple passes with heavy loads, MSP using pulling machinery involves a single pass across large parts of the site-prepared area, the significance of which remains unknown. However, the favourable effect of MSP on regeneration is well established (Sikström et al. 2020).
From an environmental perspective, any disturbance beyond the level required to facilitate efficient tree regeneration is undesirable. Planting spots of about 0.25 m2 have been found to be sufficient for successful regeneration (Örlander et al. 1990). Two thousand planting spots of this size per hectare would cover 5% of the area. Hence, there is potential to reduce the disturbed area proportion associated with most current MSP practices (Sikström et al. 2020). A further concern is to execute MSP in ways that avoid increasing erosion and soil transport to surface water bodies. To avoid increasing sediment, nutrient, C, and Hg exports to nearby streams and lakes, more knowledge about the widths of protection zones required for different MSP practices would be valuable. In areas with a shallow groundwater table, for example groundwater discharge areas beside surface water bodies, pits and furrows may extend below the groundwater table, and furrows may then act as drainage channels, potentially increasing hydrological connectivity and sediment and element exports to adjacent surface water bodies (Eklöf et al. 2016). Such unintended drainage could also impair the ecological value of wet forest patches which have been left for conservation purposes.
5 Nitrogen
In conifer forests growing on mineral soils in Sweden and Finland, the largest N pool is typically found in the soil with a substantial part in the humus layer (Merilä et al. 2014; Sponseller et al. 2016). Redistributing soil horizontally and vertically through MSP changes the spatial distribution of soil N (and other elements), and affects soil temperature and moisture conditions and vegetation development within the microsites created (Örlander et al. 1990; Palviainen et al. 2007; Piirainen et al. 2015). These changes could potentially affect soil N cycling and N pools as well as N concentrations in and exports to soil water, groundwater, and streams.
5.1 Microsite scale
Decomposition (i.e., mass loss) and N release rates were higher for green Norway spruce and Scots pine needles placed underneath mounds, disc trenched ridges, and plough tilts than for needles placed on the non-prepared A00 layer, over the first three or four years after MSP (Johansson 1994). Higher decomposition rates were also reported for two types of Norway spruce needle litter when placed underneath disc trenched ridges than on the non-prepared forest floor, but only one type of needle litter (collected from two-month-old logging residue) showed a net release of N at the end of the study (Lundmark-Thelin and Johansson 1997). At a site in east-central Sweden, higher decomposition rates of brown needle litter from Scots pine were found following incubation within a double humus layer covered by mineral soil or in a mixture of humus and mineral soil (Mjöfors et al. 2015). Here the N content of the needle litter tended to increase during the first year of incubation (cf. Lundmark-Thelin and Johansson 1997), particularly when incubated within a double humus layer covered by mineral soil (100% increase after one year). Mjöfors et al. (2015) conclude that the initial increase of the needle litter N content results from N being transported into the litter from external sources driven by the N requirements of the decomposers. The same study also looked at Scots pine root litter with a diameter of c. 6 mm: in this case there were few significant differences in remaining N content between the studied microsite types (Mjöfors et al. 2015).
At two sites in Finland, a higher store of NH4-N was found in the upper organic layer of mounds in the second year after mounding than in non-prepared soil (Nieminen et al. 2012). After disc trenching, higher NH4-N concentrations (Piirainen et al. 2007) were reported below ridges than below furrows in soil water collected in the B horizon of podzolised soil using zero-tension lysimeters. The ridges were recolonised by field-layer vegetation more rapidly than the furrows were, and five years after disc trenching the N content of the mosses, field layer, and roots growing on the ridges was 40 kg ha–1 compared with 16 kg ha–1 in the furrows (Palviainen et al. 2007). Recolonisation by ground vegetation and low N deposition was found to mitigate N leaching (Piirainen et al. 2007).
In a study at two other sites on podzolised soil in Finland, Smolander and Heiskanen (2007) drew the overall conclusion that mounding and inverting did not significantly affect the net N mineralisation rate in the humus layer (per kg organic matter) over the first three years. However, in the first growing season after MSP, the mean net N mineralisation rate was higher in the humus layer of the mounds or inverted spots than in intermediate areas with non-prepared soil, and in the second season net nitrification increased both for mounds and inverted spots but returned to low levels in the following season (Smolander and Heiskanen 2007).
5.2 Groundwater quality
Studies of groundwater quality following MSP are rare. At the Pahalouhi and Hautala sites located on sand or sandy till in central Finland groundwater samples were collected after clearcutting and clearcutting followed by ploughing, respectively, using 4–6 m long tubes with holes in the lowermost 1.5 m (Kubin 1998). The study started before clearcutting in 1985 and covered 12 years. At Pahalouhi, visual comparison of the graphs for clearcutting and waste wood left on site vs. clearcutting and ploughing seemed to show moderate elevations in the mean NO3-N concentration of groundwater (roughly 0.1–0.2 mg l–1 in magnitude) for some years after ploughing. At Hautala, the temporal patterns of the NO3-N concentrations in groundwater tended to differ between the two treatments. The lag period before NO3-N concentrations started to rise was longer after clearcutting and ploughing. The maximum mean NO3-N concentrations reported for the two treatments were <0.8 mg l–1 for both sites. In another study, Mannerkoski et al. (2005) reported that the groundwater NO3-N (and Cl-) concentrations in groundwater wells located in upland mineral soils increased after clearcutting and disc trenching, peaking at 1.3 mg l–1 for NO3-N. The authors concluded that these increases were below the levels that could present a health risk.
5.3 Impact of previous fertilisation
Forest fertilisation with N generally increases the N store of the organic top-soil layer and reduces the C to N ratio of the humus layer (Mälkönen 1990; Nohrstedt 1990; Ring et al. 2011). Concerns about the fertiliser-induced increase in the soil N store leading to post-harvest N leaching were already being raised in the 1970s by Tamm et al. (1974). Since MSP affects the upper soil layers, the physical alterations to the upper soil could potentially affect N concentrations in soil water and N leaching following MSP. However, in three harvested fertilisation experiments in Sweden (Hagfors, Nissafors and Billingsjön) with subsequent MSP, no significant statistical interaction between previous N fertilisation and MSP was detected in terms of N concentrations in soil water collected at c. 0.5 m soil depth (Ring et al. 2013, 2018). The total N fertilisation rates studied were 450 and 900 kg ha–1 at Hagfors, 360 kg ha–1 at Nissafors, and 360–1800 kg ha–1 at Billingsjön. Previous fertilisation tended to increase the N store of the organic layer (i.e., the fermentation and humus layers, FH) at all three sites (Nohrstedt 1990; Ring et al. 2011).
In the Billingsjön fertilisation experiment, located on a low-productivity site in central Sweden, disc trenching simulated by digging furrows and ridges with a spade was introduced as a sub-plot treatment two years after clearcutting (Nohrstedt 2000). A soil study conducted five years after MSP indicated that the area-integrated soil stores of inorganic N for simulated disc trenching were not significantly different from those in non-prepared soil (Nohrstedt 2000). Soil-water samples were collected from c. 0.5 m soil depth using suction cups with the intention of capturing the combined effect of all microsites associated with disc trenching. The effect of simulated disc trenching on the soil-water NO3-N concentrations started to develop after about five years, with lower NO3-N concentrations in soil water after simulated disc trenching than in non-prepared soil, and this effect persisted for about nine years (Ring et al. 2018). Higher N uptake by vegetation following MSP, as indicated by the higher biomass of the juvenile Scots pine trees, was given as a possible explanation.
The Hagfors and Nissafors sites were similar in terms of experimental design and site quality (5.9 and 5.5 m3 ha–1 yr–1, respectively) (Ring et al. 2011, 2013). Nitrogen fertilisation was the main-plot treatment and MSP the sub-plot treatment (each main plot was divided in two sub-plots). The sites were disc trenched within two months after whole-tree harvesting and soil-water samples were collected at 40–45 cm depth (from the original soil surface) below ridges, furrows, and zones between furrows, respectively, in disc-trenched sub-plots, and in non-prepared sub-plots. For both sites, no significant overall effect of previous fertilisation on the soil-water NO3-N concentration was found during the first four or five seasons after harvesting and disc trenching, but a significant effect of microsite was noted (Ring et al. 2013). At Hagfors, the mean NO3-N concentration below ridges (0.28 mg l–1) was significantly higher than below furrows (0.069 mg l–1), while at Nissafors the mean NO3-N + NO2-N concentration below the ridges (3.0 mg l–1) was significantly higher than on all the other microsites (≤0.49 mg l–1), non-prepared soil included. The rise in NO3-N concentration tended to be more pronounced below the ridges at Nissafors. At Nissafors, the generally warmer and wetter weather (on an annual basis), c. 60% higher dry mass of the FH-layer, and lower C to N ratio (32 compared with 39) than at Hagfors (Ring et al. 2011, 2013) may have contributed to the higher NO3-N concentrations. Other important factors that could influence the concentration, but which are unknown in this case, include the re-vegetation rate above the soil-water samplers, how well the spatial variation in concentration was captured, and the observation that 23% of the samples at Nissafors were missing, probably because the well-drained sandy soil rendered sampling difficult.
In modelling the N leaching from 0.5 m soil depth at the Hagfors site over the first six years, including just the unfertilised control and the treatment using 450 kg ha–1 of N, higher mean inorganic N leaching was reported after disc trenching in both cases, but more so with the N treatment than for the unfertilised control (Rappe George et al. 2017). For the 450 kg ha–1 regime, the annual mean leaching rate of inorganic N without disc trenching was estimated as 2.3 kg ha–1 yr–1 (range 0.8–6.9 kg ha–1 yr–1) and 6.0 kg ha–1 yr–1 (range 1.8–17 kg ha–1 yr–1) with disc trenching. For the unfertilised control, the annual mean N leaching rate without disc trenching was 3.1 kg ha–1 yr–1 (range 1.4–23 kg ha–1 yr–1) and 4.6 kg ha–1 yr–1 (range 1.9–13 kg ha–1 yr–1) with disc trenching. The modelled leaching after disc trenching represented an area with ridges covering 37%, furrows 41%, and areas between two furrows 21%.
In a harvested fertilisation experiment on podzolised soil in Finland, higher concentrations of NO3- and total N were indicated in soil water collected below mounds (using zero-tension lysimeters) than in non-prepared soil, despite little effect on the net formation of NH4-N and (NO2 + NO3)-N and nitrification in the buried humus layers of the mounds (Smolander et al. 2000). The elevated NO3- and total N concentrations recorded in soil water were largely attributed to the doubled amount of organic matter in the mounds which, at the same rate of mineralisation and nitrification, could provide more N. The NH4-N and NO3-N concentrations in soil water decreased over the first two years after mounding, probably because of uptake by ground vegetation according to Smolander et al. (2000). The measurements were carried out on four main plots including an unfertilised control plot and three plots which had been fertilised with a total of 6 tonnes ha–1 of lime, 860 kg ha–1 of N, and the lime and N applications combined, respectively, over the 35 years before clearcutting. The logging residue was left evenly distributed on the plots and each plot was divided into two halves, one of which was mounded and the other of which was left non-prepared.
5.4 Ecosystem N stores
MSP has been found to change the distribution of N between different aboveground vegetation compartments and soil layers. In three conifer plantations on mineral soil in Sweden, the N stores in the trees, roots, field layer, and soil were assessed ten years after either stump harvesting followed by deep soil cultivation (inverting to a soil depth of 0.5–0.6 m on 50% or 100% of the plots), or patch scarification (Nordborg et al. 2006). Patch scarification meant that planting spots with bare mineral soil were created by scraping off 0.4 m × 0.4 m patches of the humus layer, affecting c. 5% of the area. The total N store in the trees, roots, field layer, and soil did not differ significantly between the two MSP treatments, nor did the total N store in trees, field layer, and roots. However, the N store in the trees was higher after deep soil cultivation than patch scarification, while the field layer N store was lower. In the soil (humus layer down to 0.6 m depth in the mineral soil), the total N store was not significantly different, but the stores of individual soil layers varied (Nordborg et al. 2006). At two of these sites, an additional assessment was performed 22 and 24 years after MSP treatment, respectively (Egnell et al. 2015). At those points, no significant difference between the two treatments was detected in the total ecosystem N (and C) stocks (tree biomass, litter, field vegetation, humus layer, and 0–70 cm of the mineral soil).
Five years after disc trenching of experimental plots at the Hagfors fertilisation experiment in Sweden, the summed biomass of seedlings and ground vegetation was similar to that on the non-prepared plots (Johansson et al. 2013). However, the distribution of biomass between seedlings and ground vegetation differed, with higher seedling biomass and lower ground vegetation biomass on disc trenched plots than on non-prepared plots. The total N content of the aboveground biomass was approximately 20% lower on the disc trenched subplots, due to the lower N concentration of the planted seedlings than the ground vegetation. Seedling growth was not significantly affected by previous fertilisation of the harvested stand but did increase after disc trenching (Johansson et al. 2013).
About 65 years after site preparation, the N store in the field layer and down to 1 m in the mineral soil was 16 and 19% lower on two plots that had been spade inverted (Andersforsheden site) or hoed (Rosinedalsheden site) than in the non-prepared control plots (unreplicated study design) (Örlander et al. 1996). At the Tällvattensmon site within the same study 23–24 years after ploughing, the N store in the field vegetation and down to 0.25 m in the mineral soil was 30% lower than on the control plot. Örlander et al. (1996) concluded that tree production was not negatively affected, despite these reductions in the N store. All three sites were located in northern Sweden and site preparation disturbed almost the entire surface area down to a soil depth of about 10–20 cm.
5.5 Conclusions and research needs
The relatively few studies on needle decomposition, soil N transformation, and soil-water chemistry mainly after mounding and disc trenching in recharge areas, indicate that MSP affects N cycling at the microsite scale during the first few years after treatment with elevated rates or concentrations in mounds and ridges. Higher NO3- or NH4-N concentrations in soil water have been recorded below mounds and ridges than below furrows or non-prepared soil. This effect seems to be temporary, and the decline appears to be driven by uptake by the emerging ground vegetation. Post-treatment N mineralisation, weather, and the length of the fallow period may also contribute, but this has been little studied. Management of the previous forest stand, for example, N fertilisation and whole-tree harvesting, could also potentially affect N cycling after MSP, but no overall conclusions can be drawn from the few studies that have been carried out. There are gaps in knowledge regarding the impact of MSP on groundwater quality and the contribution that MSP makes to stream N exports over that caused by the preceding clearcutting. The impact of streamside management such as delineation of non-prepared protection zones with and without forest cover also needs further investigation.
MSP redistributes the soil N store, but the effect depends on both MSP practice and disturbance intensity. In addition, MSP may affect the allocation of N in the biomass . Long-term impacts on the soil N store have been investigated after intensive MSP at a few sites (with unreplicated treatments) and the results suggest a reduction (Örlander et al. 1996). If current MSP practices affect the soil N store over the longer term remains unknown. When studying the impact of MSP on ecosystem N (and C) stores, all significant N (and C) stores within the system and fluxes at a given time need to be monitored to capture possible translocations as pointed out by Nordborg et al. (2006) and Piirainen et al. (2015).
6 Carbon
6.1 Microsite scale
All MSP practices redistribute soil C horizontally and vertically by removing or burying the organic surface layer or mixing it with mineral soil. This changes the conditions for C cycling. Needle litter studies covering 2–4 years indicate that the needle decomposition rate (i.e., the mass or C loss) increases when the litter is buried in mounds or ridges or in mixtures of humus and mineral soil, compared with litter placed on the soil surface (Johansson 1994; Lundmark-Thelin and Johansson 1997; Mjöfors et al. 2015). The effects of mounding and inverting on the C (and N) transformations in the humus layer were studied over two and a half years at two mesic forest sites in Finland with podzolic soils (Smolander and Heiskanen 2007). At one of the sites, the rate of C mineralisation in the buried humus layer was significantly lower for inverting than for mounding but the amounts of microbial C and N tended to be higher. Smolander and Heiskanen (2007) concluded that this indicates a deficiency in easily mineralisable C sources in the inverted patches. At the other site, no significant difference in the C mineralisation rate of the humus layer was detected.
Carbon (and N) transformations were investigated over three years after mounding in a harvested fertilisation experiment in south-east Finland (Smolander et al. 2000, see also 5.3). Mounding decreased C mineralisation in the humus layer compared with non-prepared soil but had little effect on microbial biomass C. The authors propose that higher ground vegetation density and species composition in the non-prepared plots may have favoured soil microbial activity. They also propose that higher immobilisation of nutrients from decomposing logging residue buried within the mounds than for the residue left on top of the non-prepared soil could have contributed to decreased microbial activity in the surrounding humus layers. Moreover, microbial biomass C and C mineralisation tended to be higher in the rhizosphere of birch seedlings planted in the mineral soil of the mounds than in corresponding bulk soil (Smolander et al. 2000). The explanation offered was that root exudates and root litter from the seedlings provided substrates for the microbial population.
In undisturbed soil profiles, the input of organic matter into subsoils occurs mainly as root litter and root exudates, dissolved organic matter, and/or bioturbation as summarised by Rumpel and Kögel-Knabner (2011). MSP could potentially affect all of these processes. Higher total C or dissolved organic C concentrations in soil water collected below ridges than below furrows in the B horizon of podzolised soils were reported during the first few years after disc trenching in Sweden, in a harvested fertilisation experiment, and in Finland (Piirainen et al. 2007; Ring et al. 2013). The overall soil-water C concentration tended to be about five to ten times higher in the study in Finland than that in Sweden (where the mean concentration of total C for all microsites was 2–4 mg l–1 for the entire period). One explanation could be that different methods for soil-water sampling were used: zero tension lysimeters in Finland and suction cups in Sweden.
6.2 Ecosystem C stores
Twenty-five years after mounding, disc trenching, and ploughing at three sites in Sweden, the ecosystem C store was larger for site-prepared treatments than non-prepared controls because of a higher C store in the tree biomass (Mjöfors et al. 2017). Ten years after establishing three conifer plantations on forest land, the C stores were investigated on plots subjected to either stump harvesting followed by deep soil cultivation, or patch scarification (Nordborg et al. 2006, see also 5.4). The total ecosystem C stores did not differ significantly between deep soil cultivation and patch scarification, but the tree C store was higher, and the field-layer C store lower, after deep soil cultivation than after patch scarification. Furthermore, the total C stores in the humus layer and down to 0.6 m in the mineral soil were not significantly different for the two MSP practices, but the C stores of individual soil layers differed (Nordborg et al. 2006).
Long-term effects of MSP on soil C stores were suggested at three sites in northern Sweden (all with unreplicated designs and intensive MSP, see 5.4) (Örlander et al. 1996). At the Tällvattensmon site, a 41% decrease of the C store in the field and ground vegetation, organic debris, humus, and mineral soil down to 0.25 m depth was reported 23–24 years after ploughing. At the Rosinedalsheden and Andersforsheden sites, reductions of 6 and 21% of the C stored in the field and ground vegetation, organic debris, humus, and mineral soil down to 1 m depth were found about 65 years after hoeing and spade inversion, respectively (Örlander et al. 1996).
6.3 Conclusions and research needs
There are few studies of how MSP affects soil C cycling and stores, particularly over the longer term. Decomposition of needle litter appears to increase when buried under or mixed with mineral soil, while reduced or similar mineralisation has been reported for buried organic matter (humus), suggesting that “fresh” and “old” C-sources respond differently when buried (Mäkipää et al. 2023). Long-term studies in Denmark and Germany indicate that burying organic matter deep in the mineral soil may increase its stability (Alcántara et al. 2017). However, Mayer et al. (2020) identify a gap in knowledge regarding the decomposition rate of surface organic matter when buried deep down in the mineral soil by site preparation. They further state that the potential mechanisms of stabilisation depend on the MSP practice, and forest and soil type. This calls for further studies, at different forest sites as proposed by Mäkipää et al. (2023), and of different MSP practices. Improved understanding of decomposition of buried organic matter, including the interactions between plants and soil (Smolander et al. 2000; Mäkipää et al. 2023), could help shed light on the long-term effects of MSP on soil organic matter.
C concentrations in soil water have been found to be higher initially below ridges (disc trenching) than below furrows (Piirainen et al. 2007; Ring et al. 2013). If the leaching rates of dissolved organic C are affected, as reported by (Piirainen et al. 2007), this could potentially influence the downward transportation of elements associated with organic matter. Better understanding of the contribution of MSP to C export to nearby surface water bodies is needed (as for N). Furthermore, the consequences of executing MSP in streamside discharge areas need further investigation.
MSP increases the tree biomass C store (Nordborg et al. 2006; Mjöfors et al. 2017; Hjelm et al. 2019; Sikström et al. 2020) and may increase the total ecosystem C store (Mjöfors et al. 2017). MSP can affect the distribution of C among aboveground C compartments as well as vertically in the soil without affecting the total C store, at least in the short term (10–20 years) (Nordborg et al. 2006; Egnell et al. 2015). However, more studies are needed on the long-term effects of MSP on total ecosystem C (and N) stocks, particularly for current MSP practices.
7 Greenhouse gas emissions
7.1 Microsite scale
In situ measurements of the CO2 flux following MSP have been conducted at boreal and hemi-boreal forest sites in Sweden and Finland, using closed static chambers or a portable soil-respiration system. Measurements were performed up to three years after MSP and showed that CO2 emissions from MSP microsites with exposed mineral soil were significantly lower than from non-prepared soil (Pumpanen et al. 2004; Strömgren and Mjöfors 2012; Mjöfors et al. 2015; Strömgren et al. 2017). In the case of mounds (usually of mineral soil over humus), increased (first two years; Pumpanen et al. 2004), more or less unaffected (Strömgren and Mjöfors 2012; Mjöfors et al. 2015), and decreased (first year) CO2 emissions have all been detected (Strömgren et al. 2017). From microsites with mixed soil, Mjöfors et al. (2015) reported increased CO2 emissions during the first month in the first year and a reduction during the second year, whereas Strömgren et al. (2017) reported a reduction during the first year and unaffected emissions in the second year. The effects mentioned above were all compared with clearcutting after which logging residues were extracted. In the study by Pumpanen et al. (2004), non-prepared controls with and without logging residues were present. Where residues were present the annual CO2 emissions during years 1, 2 and 3 were 78%, 45% and 47% higher, respectively, than where there were no residues. Hence, the effect of the MSP microsites depended on the chosen control treatment. However, the directions of the differences and conclusions reported were the same, regardless of whether or not the control was covered by residues (Pumpanen et al. 2004).
At three sites with mesic to mesic-moist soils in Sweden, CH4 and N2O was monitored over two years by taking in situ air samples (Strömgren et al. 2016). The C/N in humus was 21–27 and 14–20 in mineral soil at 10–15 cm soil depth. Here, a non-replicated plot with disc trenching (one site) and mounding (two sites) was compared with a non-prepared control plot from which logging residues were removed. In addition, samples from wheel ruts were collected at all three sites. At one site with mounding, N2O emissions were highest from the non-prepared soil and mounds (double humus with mineral soil on top) and, compared with these two microsites, emissions were significantly lower from wheel ruts, while emissions from exposed mineral soil pits fell somewhere in between (but were not statistically significantly different). At this site, there was a significant relationship between soil NO3-N and NH4-N availability and N2O emissions at the different microsites. N2O emissions at the other site with mounding were generally low and not significantly different between microsite types. At the third site (disc trenching), exposed mineral soil in furrows yielded significantly higher emissions than non-prepared soil and wheel ruts, but not higher than those from ridges. The authors highlight the importance of available nitrogen fractions in the soil for N2O emission, and that the emerging ground vegetation in clearcuttings acts as a competitor for N and may thereby counteract emission. At all three aforementioned sites, CH4 uptake occurred at all mounding and disc trenching microsites, while CH4 was emitted from wheel ruts (Strömgren et al. 2016). In general, all the CH4 fluxes were low except for the emissions from wheel ruts at the most fertile site.
7.2 Area-integrated assessments
Area-integrated CO2 emissions for different MSP practices have been determined by considering the area proportions of different MSP microsites and their specific CO2 emission rates. Using this approach for a site in east central Sweden with a glacial sand podzol, where different microsites were created by an excavator, Mjöfors et al. (2015) estimated that C emissions were 7% and 10% lower in the first and second year, respectively, after patch scarification than on non-prepared soil, and 11% and 17% lower after disc trenching. In a similar study by Strömgren et al. (2017), 11% lower CO2 emissions were reported following patch scarification or mounding, and disc trenching tended to reduce emissions by 9% during the first year after treatment. During the second year, emissions were similar to those from non-prepared soil. The study by Strömgren et al. (2017) was based on measurements at 14 sites located on a north-south gradient across Sweden, representing different climates and site properties. Although there was no non-prepared control in the study by Strömgren and Mjöfors (2012), it can be concluded that mounding did not increase CO2 emission on an area-integrated basis as the emissions from non-prepared microsites were higher than or similar to those from disturbed microsites throughout the two-year study period. This was also the case for disc trenching but the area-integrated CO2 emissions from disc-trenched plots were about 10% higher than those for mounding in the second year.
The area-integrated N2O and CH4 emissions in the study by Strömgren et al. (2016) indicate that mounding and disc trenching had little or no influence on the emission of CH4 and N2O from the soil during the first two years after MSP. The authors further pointed out that the global warming potentials of CH4 and N2O were small at these sites, i.e., 0.0–0.2% for CH4 and 0.1–8% for N2O, compared to the potential of the CO2 emissions (Strömgren et al. 2016).
7.3 Conclusions and research needs
The area-integrated emissions reported from studies in Sweden indicate that patch scarification, mounding, and disc trenching do not significantly affect, or can even reduce, CO2-emission from the soil over the first one or two years, when compared with non-prepared soil (Strömgren and Mjöfors 2012; Mjöfors et al. 2015; Strömgren et al. 2017). Similarly, CH4 and N2O were not significantly affected by disc trenching or mounding at three mesic to mesic-moist sites (Strömgren et al. 2016). Hence, in the shorter term, MSP does not seem to affect the fluxes of CO2, CH4, and N2O emitted from soils on upland boreal and hemi-boreal forest sites to any great extent. At the microsite level, however, differences in CO2 emissions between microsites are apparent in the short term. Lower CO2 emissions have consistently been reported for bare mineral soil than for non-prepared soil, while varying and short-lived effects have been reported for mounds (e.g. Pumpanen et al. 2004; Strömgren et al. 2017). This variation in effects might be caused by different chemical quality of soil organic matter (SOM), site conditions (climate) and soil properties (clay content, soil moisture, pH, nutrient status) (Jandl et al. 2007), weather conditions, and MSP execution (affected soil volume, level of soil mixing etc.), all of which may affect decomposition of SOM in the mounds.
Investigations at sites with different characteristics (e.g., site productivity, soil moisture) are needed, particularly to study the fluxes of CH4 and N2O. It is important that future studies include all three gases (CO2, CH4 and N2O) in order to be able to evaluate the global warming potential of MSP.
8 Other chemical variables
8.1 Microsite scale
Higher release rates of Ca (and to some extent Mg and P) from Scots pine and Norway spruce needles were found when the needles were buried below mounds, plough tilts, and ridges than when they were placed on top of non-prepared soil (Johansson 1994; Lundmark-Thelin and Johansson 1997). For K, only minor differences between non-prepared soil and prepared soil were reported.
Creating different types of MSP microsites, exposes new soil surfaces and soil layers which can affect leaching (cf. Hildebrand 1990; Tanskanen et al. 2004; Lindroos et al. 2016; Lindroos and Ilvesniemi 2023). After disc trenching in the Kangasvaara catchment in Finland, the concentrations of Mg, Al, and Fe were significantly higher, and pH lower, below ridges than in furrows, in leachates collected using zero-tension lysimeters in the upper B horizon during the main part of a five-year long study (Piirainen et al. 2009). Higher concentrations below ridges than furrows were also noted for Ca during years four and five, and for K during year two only. The concentrations below ridges were also often higher than those on non-prepared microsites. The higher leaching rates (and concentrations) below the ridges were attributed to the release of elements from dead ground vegetation, logging residue, and the double humus layer (Piirainen et al. 2009). No significant differences between microsites were found for the SO42- concentration or the estimated annual water flux (Piirainen et al. 2007).
At the Hagfors fertilisation experiment in Sweden, significant effects of disc trenching (see 5.3) after whole-tree harvest were detected on electrical conductivity and concentrations of Na+, K+, Mg2+, Ca2+ and Cl− (also for NO3-N, total N, and total C, see sections 5 and 6) in soil water collected at 40–45 cm depth over a five year period (Ring et al. 2013). The highest mean concentrations were generally found below ridges and the lowest below furrows. Mean electrical conductivity and the mean concentrations of Na+, K+, Cl−, total N, and total C were lower below furrows than in non-prepared soil. There was no interaction between fertilisation and disc trenching except for Mn.
In a study in northernmost Finland 12 years after tilt or shoulder ploughing, soil samples were collected from 0–20 cm depth in the mineral soil on non-prepared microsites, trenches, and tilts or shoulders (Sutinen et al. 2019). Significant differences between at least two of these microsites in median concentrations were indicated for 14 elements (C, N, P, S, Na, K, Ca, Al, Fe, Mn, Ba, Cr, Sr and Zn) when extracted with ammonium acetate and 21 elements (Ba, Be, Ca, Co, Cr, Cu, K, La, Li, Mg, Mn, Na, Ni, P, S, Sc, Sr, Ti, V, Y and Zn) when extracted with Aqua Regia. The element concentration in trenches, following one or both ploughing practices, was significantly lower than on non-prepared microsites for C, N, P, S, Na, K, Mg, Ca, and Al. One proposed explanation for the lower concentrations in trenches was enhanced leaching during snowmelt periods, caused by the removal of the organic layer and accumulation of snow in the trenches (Sutinen et al. 2019). This leaching can also decrease the electrical conductivity of the soil solution over the longer term (Sutinen et al. 2006, 2010). Forty-six years after disc trenching of a burnt forest site in northern Finland, mineral soil samples from furrows showed lower electrical conductivity and lower concentrations of Ca, Mg, Na, Fe (extracted with ammonium acetate), and higher concentrations of Cu, Mn, and Zn than samples from non-prepared soil (Närhi et al. 2013). The mineral soil was sampled down to a depth of 20 cm but the E horizon was removed.
In a study of soil chemistry performed 17 years after ploughing, chemical weathering, higher percolation rates, and dissolved organics were presented as possible explanations for the more rapid depletion of some elements (MgO, CaO, Sr, and V) in the upper part of the plough trenches (consisting of the B horizon of a podzol) than in the corresponding horizons of non-prepared soil (Lindroos et al. 2016). The investigated soil contained minerals with high weathering rates and the dissolved organics originated from the new organic layer and litter accumulation in the furrows. In the plough tilts, changes in concentration were mainly due to mixing of the original horizons and this tended to mask any depletion caused by weathering, according to Lindroos et al. (2016).
The nutrient stores of the field-layer vegetation five years after disc trenching were investigated at the Kangasvaara site in Finland and the Hagfors site in Sweden. At Kangasvaara, the field-layer stores of K, Ca, and P (and N) were similar for vegetation growing on ridges and in non-prepared soil but lower for vegetation growing in furrows (Palviainen et al. 2007). Five years after disc trenching (and whole-tree harvesting) at Hagfors, the ground-vegetation stores of N, Ca, Cu, K, Mg, Mn, P, S, and Zn were lower in disc-trenched subplots than in non-prepared subplots, because of lower ground vegetation biomass in the disc-trenched subplots (Johansson et al. 2013).
8.2 Conclusions and research needs
Elevated cation concentrations in soil water below ridges and mounds may result from elevated soil-water NO3- concentrations. Exposing new mineral soil surfaces by MSP and, initially, low vegetation uptake may also affect cation concentrations. Intense percolation may deplete soil element stores. In harsh climates, snow accumulation in plough trenches combined with a distinct snowmelt period was proposed as an explanation for the decreased stores of some elements in the soil (Sutinen et al. 2019). In milder climates with no or little snow accumulation, the rainfall regime, atmospheric deposition, and litter accumulation are probably important factors in any changes in soil element stores.
Studies into how soil chemistry, other than N and C, and soil evolution are affected by other MSP practices than ploughing are scarce. More information on trace metal mobility following MSP is needed, since trace metals like Hg, Cd, and Pb which originate from anthropogenic deposition have accumulated in the soil, particularly the mor layer (for podzols), in north European forests (Alriksson 2001; Johansson et al. 2001; Bringmark et al. 2013). In summarising studies of how various forestry operations affect Hg mobilisation from soils to aquatic environments, Eklöf et al. (2016) found substantial variation between sites in the effects on total Hg and methylmercury, particularly for methylmercury which is bioavailable. There was little information on how MSP alone affects Hg mobilisation. Eklöf et al. (2016) state that the mobilisation of Hg from the soil caused by forestry activities results either from induced changes in hydrology, i.e., soil moisture, runoff, and groundwater levels, and/or the rate of net Hg methylation.
9 Ground vegetation
Reducing vegetation competition to promote the desired crop is one explicit reason for carrying out MSP (Örlander et al. 1990). However, this can have unwanted side effects, for example, impacts on reindeer lichen forage. In a field experiment located in a Scots pine-heath type boreal forest in northern Sweden, the influence of MSP on reindeer (Rangifer tarandus tarandus L.) grazing opportunities was evaluated (Roturier and Bergsten 2006). MSP was undertaken using a HuMinMix scarification unit mounted on the rear of a forwarder. This unit consists of a rotating metal wheel with teeth which are pushed a few cm into the mineral soil and creates a mixture of milled humus and mineral soil (HuMinMix) in continuous tracks. Mounds can also be created at desired intervals. In the study, five epigeic lichen species which are important as reindeer forage in this forest type (four Cladina spp. and one Cladonia sp.) were monitored. The MSP treatments disturbed 14% to 28% of the surface area. Six growing seasons after MSP, the lichen cover and volume were 10–20% lower in MSP plots than in non-prepared soil (area-weighted averages).
In a survey study in northern Sweden, Roturier et al. (2011) found that reindeer lichen (four Cladonia spp.) cover and biomass can be completely re-established about a decade after MSP using HuMinMix (disturbing 15% of the soil surface), while they estimated that several decades, possibly more than half a century, might be needed after disc trenching (40–60% soil surface disturbance). The survey encompassed 17 clearcuttings of former Scots pine forests, of which 11 sites were surveyed 1–15 years after disc trenching and six sites 2 and 8–9 years after MSP using HuMinMix. The faster re-establishment of lichen cover and biomass after HuMinMix was explained by the more favourable substrate created by HuMinMix. This substrate includes fragments of lichen thalli that more easily attach to the mixture of humus and mineral soil than to the bare mineral soil in furrows (cf. Roturier et al. 2007). Furthermore, some Cladonia species are not pioneer species and dependent on pioneers such as crustose lichens and bryophyte species to first colonise bare mineral soil (cf. Roturier et al. 2011). However, bare mineral soil can favour recolonisation of lichens from intact lichen mats surrounding bare mineral soil (Roturier et al. 2007). The general conclusion drawn by Roturier et al. (2011) was that both the degree of initial disturbance caused by MSP and the re-establishment rate of reindeer lichens must be considered when estimating the effect of MSP on lichen loss for reindeer grazing over time.
Tonteri et al. (2022) analysed how lichen cover responds to forest management, site, and stand variables, and to co-existing plants, in different boreal vegetation zones in Finland. The study was based on a survey of understory vegetation cover in 1721 permanent sample plots on mineral soil on productive forest land. The plots were established by the Finnish National Forest Inventory. The cover of “all ground lichens” showed a significant response to MSP (undertaken in connection with regeneration cutting over the previous 25 years). On ploughed sites, lichen cover was about a third of that in non-prepared sites, whereas on sites with less intensive soil treatment, including disc trenching, scalping, and mounding, lichen cover was higher than on non-prepared sites. Similar results were reported for the cover of “reindeer lichens” (i.e., four Cladina spp.) which was analysed for a subset of the sample plots, namely plots on nutrient poor sub-xerix and xeric sites (n = 610).
Based on preliminary results from a chronosequence study of disc trenched and ploughed clearcuttings in northern Sweden (63–68° N), covering 1–20 years after harvest, Eriksson and Raunistola (1990) suggest that it takes more than 20 years for the bare mineral soil in ploughed areas to be re-covered by ground vegetation and that this time span is shorter after disc trenching. The disturbed surface area proportions were 78–87% for ploughing and 44–52% for disc trenching. Compared with similar mature forest stands which were uncut, observed changes in the ground vegetation after MSP (and clearcutting) included less cover of dwarf shrubs and greater cover of grasses and herbs. Of individual species, Empetrum hermaphroditum, Vaccinium myrtillus and mosses such as Pleurozium schreberi (Bird) Mitt. and Hylocomium splendens (Hedw.) B.S.G. decreased after ploughing, whereas Deschampsia flexuosa L. and pioneer mosses such as Polytrichum piliferum (Hedw.) and Ceratodon purpureus increased. Eriksson and Raunistola (1990) concluded that 20 years is too short a period to determine any differences in the composition of ground vegetation in mature new stands compared with previous harvested mature stands.
In an investigation in northern Sweden, Norberg et al. (1997) used small plots (0.6 m × 0.6 m) to study patch site preparation (and steaming) in a clear-cut site classified as a Myrtillus-type (Cajander 1949). The forest floor vegetation was dominated by V. myrtillus and the feather mosses P. schreberi and H. splendens. Four years after treatment, the total cover of both field- and bottom-layer vegetation was significantly lower in patch-treated than non-prepared plots. In the field layer, the cover of V. myrtillus, Vaccinium vitis-idaea and D. flexuosa was lower in patches, whereas the cover of Betula spp. was somewhat higher. In the bottom layer, recolonisation was fast in patch plots, primarily due to the establishment of mosses such as Polytrichum juniperinum (Hedw.) and P. piliferum, whereas P. schreberi and H. splendens, along with Dicranum spp., were more prevalent on the non-prepared plots.
Bergstedt et al. (2008) investigated ground vegetation response to harvesting (four intensities) and MSP in a field experiment 14 years after treatment, in four blocks in west-central Sweden. The low productivity site (site quality 2.0–2.5 m3 ha–1 yr–1) was located on two slopes where soil moisture conditions varied from xeric through mesic to hydric. To evaluate the effect of MSP, plots subjected to clearcutting and MSP (disc trenching in two blocks and ploughing in two blocks) and planting (treatment “Conventional”) were compared with non-prepared plots that were almost clearcut and planted (treatment “High”; 0–19% tree retention by volume; tree diameter <0.08 m left on site). All vascular plants (except for trees >1 m in height), mosses and lichens were recorded and the effect of MSP, i.e., “Conventional” compared with “High”, was analysed for a total of 28 species and groups of species. Bergstedt et al. (2008) state that their study showed that V. myrtillus was more sensitive to cutting intensity than to MSP, whereas V. vitis-idaea was more or less unaffected by cutting intensity but strongly negatively affected by MSP. E. nigrum was also disadvantaged by MSP whereas P. sylvestris, Betula spp., and D. flexuosa were promoted by MSP. Of the mosses, the authors concluded that Polytrichum spp. were promoted by MSP whereas both P. schreberi and H. splendens were negatively affected. Finally, of the lichens, Cladina arbuscula was negatively affected by MSP but other Cladina spp., such as Cladina rangifera, seemed indifferent to both cutting intensity and MSP. The authors concluded that 14 years after treatment they were unable to detect any difference in species richness between any of the treatments evaluated. They also suggested that MSP had a profound and long-lasting effect on the ground-layer flora, possibly explained by burial of part of the flora under heaps of the disrupted vegetation layer together with humus and soil from the scarified area, and that the ground vegetation succession after MSP starts from a patchy area with vegetation-covered areas intermingled with bare mineral soil. Fourteen years after MSP, bare mineral soil patches were still present at the site investigated.
In eastern Finland, the development of ground vegetation on ridges, in furrows, and on non-prepared soil was investigated from two to five years after disc trenching of a clearcutting (Kangasvaara site) (Palviainen et al. 2007). The results indicated that field layer vegetation and mosses responded differently to disc trenching, and that the biomass development on ridges, in furrows, and on non-prepared soil proceeded at different rates. Colonisation of mosses was rapid in furrows and slow on ridges. Hence, at the end of the study period, the biomass of mosses on ridges remained significantly lower than that on non-prepared soil and furrows. The field layer biomass on non-prepared soil was significantly higher than on ridges and in furrows throughout the study. Furthermore, the field layer biomass was significantly higher on ridges than in furrows at the end of the study. The total ground vegetation biomass of ridges and non-prepared soil became similar in the fourth year after MSP, whereas it remained significantly lower in furrows throughout the study period and was about one third of that on the other two microsites in the fifth year.
Vanha-Majamaa et al. (2017) studied the response of understory vegetation cover to five different felling treatments, of which three included disc trenching, up to ten years after treatment. Species´ extinction and colonisation patterns were also examined. The study was conducted in mature Norway spruce dominated forests in southern Finland and the inventory included tree saplings (up to 1.8 m in height), other vascular plants, mosses, liverworts, and lichens. Although the effect of MSP could not be entirely separated from the effect of harvesting alone, the authors conclude that, in addition to harvest intensity, MSP influences understory vegetation on the levels of the whole vegetation community, different species groups, and individual species. In general, on the community scale, treatments involving disc trenching changed the vegetation composition distinctly, due to mechanical soil disturbance and the colonisation of pioneer species. This effect could still be observed ten years after treatment. An immediate reduction in species richness was seen following all treatments, but the recovery was rather fast following most of the treatments for most species groups. However, bryophytes, particularly liverworts, had not recovered ten years after treatment. The authors considered that the MSP was largely responsible for the decline of late successional understory species, affecting both their cover and species number. At the same time, the species number increased after disc trenching as it created favourable microhabitats for early successional, disturbance-dependent pioneer species and ruderal species. Furthermore, they concluded that, while the dynamics of vascular plants depended on harvest intensity, the dynamics of non-vascular species also depended on MSP. They found no clear evidence that the intensive treatments tested had caused vascular plant loss. Regarding changes in species groups, herbs seemed to be the most resilient group. Of the dwarf shrubs, the results suggested that the decrease in V. myrtillus cover was largely caused by the disc trenching. The Rubus idaeus cover increased, particularly in the treatments with MSP. The cover of grasses was also highest in treatments with MSP, D. flexuosa being the most abundant species, although grass cover after ten years was not significantly different to that pre-treatment. The moss P. schreberi seemed to be disadvantaged by disc trenching.
9.1 Conclusions and research needs
Unequivocally, MSP initially reduces ground vegetation surface cover and biomass. Species composition is also affected by MSP and can be long-lasting, probably longer than the 10–20 years that are documented in the available studies. It is difficult to generalise how MSP affects ground species cover and abundance because it depends on the composition of the original plant community, MSP intensity, and the establishment rate of different species. However, a common effect on species cover seems to be a decline in late succession understory species. At the same time, MSP creates microhabitats that favour the establishment of pioneer and ruderal species (Bergstedt et al. 2008; Vanha-Majamaa et al. 2017). Moreover, non-prepared strips and patches provide refuge for remnant communities of forest species (Newmaster et al. 2007). In the studies by Bergstedt et al. (2008) and Vanha-Majamaa et al. (2017), species richness does not seem to have been affected by MSP 10–14 years after MSP.
For individual species and species groups in boreal forests, the cover of some feather mosses, e.g., P. schreberi and H. splendens, as well as Dicranum spp. and dwarf shrubs (V. myrtillus L., V. vitis-idaea L. and E. hermaphroditum) generally decreases after MSP, whereas pioneer mosses, such as Polytricum spp. and Ceratodon purpureus, become abundant in furrows, while grasses and herbs (mainly D. flexuosa and Epilobium angustifolium L.) can grow vigorously on ridges. Roturier et al. (2011) observed both increased and decreased lichen species numbers 10–15 years after treatment depending on the MSP method used. For reindeer forage lichens, it seems possible to both reduce the disturbed lichen area and to increase the re-establishment rate by using an appropriate site preparation technique (Roturier and Bergsten 2006; Roturier et al. 2011).
A methodological issue in some studies is the difficulty to strictly distinguish the effect of MSP on ground vegetation from that of harvesting (Bergstedt et al. 2008; Vanha-Majamaa et al. 2017). A challenge to interpreting the results of the survey study by Tonteri et al. (2022) is that the occurrence of lichens prior to MSP being carried out probably affected which MSP practice was selected, with less intensive MSP practices being used on sites with high initial lichen cover and ploughing on sites with low cover. In addition, there is some uncertainty about which types of sites were included as the non-prepared controls.
More long-term studies (>15 years) on changes in ground vegetation cover, in terms of both abundance and composition, after MSP are needed in order to establish whether or not the observed changes are reversible. From a reindeer-forage perspective, it is important to further study the re-establishment of reindeer lichens after different MSP practices. In future studies on ground vegetation, it is essential to separate the effects of harvesting and MSP.
10 Discussion
Considering the large areas annually subjected to MSP in Sweden and Finland (Fig. 1), rather limited research on the environmental impacts of MSP has been undertaken (Table 1). In some cases, the study design renders it difficult to isolate the effect of MSP from other forest management practices, usually clearcutting. Thus, it is difficult to draw far-reaching general conclusions for most of the variables analysed in this review.
Given the nature and purpose of MSP, i.e., to physically impact the soil to promote regeneration, it inevitably alters the soil. As presented in this review, such alterations include soil surface microtopography, soil profile and element redistributions, and exposure of mineral soil. These changes may, in turn, affect vegetation cover and composition, erosion, leaching, and exports of elements and sediment to surface water bodies. It is vital to remember that MSP represents a wide range of disturbance intensities (see section 4) and that the environmental impact also depends on its execution and on-site conditions.
In light of increasing concerns about climate change, the potential impacts of forestry on forest C sequestration and soil C store are highly pertinent. In earlier reviews, the common view offered is that MSP generally increases decomposition and loss of soil C, reduces the soil C store, and that the reduction is related to the degree of disturbance (Johnson 1992; de Wit and Kvindesland 1999; Jandl et al. 2007). This view is contradicted by the more recently reported in situ measurements of soil CO2 emissions after MSP in Sweden (Mjöfors et al. 2015; Strömgren et al. 2017). There are several possible explanations for the reported differences. In the review by Johnson (1992), the studies on site preparation referred to were mainly performed in climatic regions outside the focus area of this review, and other site-preparation practices were also included (e.g., burning, litter raking). Johnson (1992) also points out a methodological problem, namely, that often it is not possible to separate the soil C loss caused by displacement from that lost by decomposition. Another methodological problem is to detect small storage changes in large soil stores such as soil C. In this context, measurement of CO2 fluxes from the soil is a more sensitive method than soil sampling. However, both methods include uncertainties related to sampling highly variable soil C stores across fine spatial scales (Haukenes et al. 2022). Moreover, there is more knowledge about the effects of forest management practices, including MSP, on C in the organic surface layer than there is for stable C pools in the mineral soil (Jandl et al. 2007). In summary, MSP by patch scarification, mounding, or disc trenching, undertaken at clearcuttings in boreal and hemi-boreal forests, does not seem to affect the loss of C to the atmosphere significantly, while the amount of C leached from the soil after these MSP practices remains relatively unknown. In addition, the same practices generally increase above-ground biomass production (Nordborg et al. 2006; Mjöfors et al. 2017; Hjelm et al. 2019; Sikström et al. 2020). Increased above-ground biomass production can contribute to increased accumulation of stable plant-derived C in the soil, although the rate of soil C accumulation is low compared to the aboveground accumulation rate in a forest ecosystem and the stabilisation process is slow (Jandl et al. 2007). Thus, as a single operation, MSP by patch scarification, mounding or disc trenching seems to contribute to increasing the ecosystem C store by contributing to high forest growth while avoiding significant C loss (Mjöfors et al. 2017). Nonetheless, most current MSP practices generally cause unjustified soil disturbance and need be developed further to cause only the disturbance necessary to achieve successful regeneration.
10.1 Research needs
As previously mentioned, there is limited research on the environmental effects of MSP, particularly regarding the longer-term effects (>5–10 years) of the MSP practices commonly used today, and the influence of climate and site productivity (cf. Table 1, Fig. 2). Multiple investigations have been performed at some sites, providing a more comprehensive picture of impact at these sites, but additional studies are needed for various MSP practices and their execution, to shed light on the influence of factors that vary geographically, namely climate, site productivity, and soil type and geochemistry. Site productivity is likely to affect the impact of MSP given the differences in, for example, soil N store and organic surface layer C to N ratio (Högberg et al. 2021), but how this is manifested is unclear and needs further research.
More field studies could also help distinguish the effects which relate to study design and methods (e.g., Weihermüller et al. 2007) and identify the interaction with other forestry measures performed soon before or after MSP, for example leaving forest buffers, harvesting logging residue or stumps, drainage activities, and wood-ash application (cf. Mayer et al. 2020). With few available studies, detailed comparisons of different studies are constrained by variations in study design and methods. For example, soil-water chemistry was investigated after disc trenching at both the Kangasvaara and Hagfors sites, but the logging residue was left on site at Kangasvaara and harvested at Hagfors, and different techniques for soil-water sampling were used (Piirainen et al. 2007; Ring et al. 2013). Unlike the Kangasvaara site, the experimental design of the Hagfors site also included previous N fertilisation.
Although knowledge has improved over recent decades, there is still little evidence about how the effects of forestry activities in upland areas (groundwater recharge areas) on soil-water chemistry are transferred to adjacent surface water bodies, and the role of streamside groundwater discharge areas. The riparian zone has a strong impact on stream-water chemistry (Bishop et al. 2004; Ledesma et al. 2013, 2018) and has been found to be more important than upland areas for the N and total organic C chemistry of boreal streams (Fölster 2000; Blackburn et al. 2017). Soil type (organic soils), superficial groundwater flow, and groundwater table fluctuations have been identified as key factors for N and total organic C export from the riparian zone (Fölster 2000; Grabs et al. 2012; Blackburn et al. 2017). Hence, clearcutting could potentially increase N and total organic C exports to adjacent streams and lakes as it causes elevation of the groundwater table and expanded streamside discharge areas due to decreased evapotranspiration (cf. Lundin 1979; Bishop et al. 2004; Schelker et al. 2012; Ledesma et al. 2018; Wei et al. 2022), disregarding any effect on denitrification. The reduction in evapotranspiration by clearcutting ought to exceed the effect of subsequent MSP by far. At the Balsjö catchments in northern Sweden, Eklöf et al. (2014) did not detect an additional effect of disc trenching on discharge compared with merely clearcutting. Thus, over the short term, the elevation of the groundwater table in streamside areas is probably mainly caused by clearcutting. However, since MSP improves regeneration success and initial tree growth (Sikström et al. 2020), it could potentially reduce nutrient exports to nearby surface water bodies over the longer term by shortening the period over which the elevated groundwater table and increased runoff persists, as also proposed by Eklöf et al. (2014), but this calls for more research. Further, leaving wide (30–50 m) forest buffers next to surface water may reduce excessive nutrient and sediment exports (Ahtiainen and Huttunen 1999), but little is known about the efficacy of current forest buffers in Sweden and Finland (Kuglerová et al. 2020; Ring et al. 2022) in mitigating nutrient and sediment loads to surface water. More knowledge on the role of streamside areas and their management could help determine the protection zones required to safeguard stream-water quality.
10.2 Suggestions for operational forestry
The environmental impact caused by MSP can be reduced through careful planning and skilful execution. The prime concerns are to prevent elevated exports of sediments, nutrients, dissolved organics, Hg, and other trace metals. In addition, consideration and adaptation to reindeer husbandry is important. Although beyond the scope of this review, it is also important to safeguard other values, for example retained trees and tree groups left for biodiversity and ecological purposes, social values, and historical remains. Topography, hillslope inclination, soil erodibility, climate, presence of surface water bodies, and location of groundwater discharge areas (and their expansion after clearcutting) must be considered when planning and executing MSP (Carling et al. 2001; Borrelli and Schütt 2014; Sikström and Hökkä 2016). Special attention should be paid when considering MSP on highly erosive soils and hillslopes with high inclination (cf. Aust and Blinn 2004). During the planning stage, careful examination of the type of disturbance required for successful regeneration must be assessed for individual sites, an appropriate MSP practice selected, and sufficient protection zones next to sensitive areas and surface water bodies delineated. The execution must be adapted to on-site conditions, for example by adjusting the ground pressure of discs during disc trenching, and directing furrows to avoid erosion. Systematic follow-ups and good communication among the staff involved could help improve procedures and implementation.
10.3 Concluding remarks
Valuing the benefits of MSP against undesired environmental impact is complicated as indicated in this review, due to gaps in knowledge, the wide range in MSP practices, their disturbance intensity and execution, and the different types of impact rendering comparisons difficult. The beneficial effects of MSP are improved regeneration success and possibly reduced usage of pesticides against pine weevil (H. abietis), since planting seedlings in areas with pure mineral soil reduces pine weevil attacks (Wallertz et al. 2018). The potential undesired effects of MSP, according to current knowledge, include increased erosion and sediment transport, disturbed soil profiles, elevated nutrient leaching and trace metal exports, and long-term effects on ground vegetation cover and species composition. However, it is difficult to compare the desired and undesired impacts of MSP in general, since the undesired impacts relate to the disturbance intensity and consideration measures taken.
The most common MSP practices currently used in Sweden and Finland, disc trenching and mounding, generally disturb considerably higher surface area proportions than is required for regeneration success. Continuous efforts to develop regeneration and MSP practices and technology are important to limiting soil disturbance intensity to the level necessary for successful regeneration. Finally, the values of preserving intact soil profiles should be assessed.
Authors´ contributions
Both authors (E. Ring and U. Sikström) contributed substantially to the conception of the research question, design of the work, analysis, writing and critical revision of previous versions of this article.
Acknowledgements
Sees-editing Ltd. (https://www.sees-editing.co.uk/) contributed with linguistic refinement of an earlier version of the manuscript. We thank Knut-Erland Berglund for contributing to producing the maps in Fig. 2.
Funding
This study was funded by a grant (no. 2018-634) from Skogssällskapet and Skogforsk´s framework agreement funding.
References
Ahtiainen M (1992) The effects of forest clear-cutting and scarification on the water quality of small brooks. Hydrobiologia 243–244: 465–473. https://doi.org/10.1007/BF00007064.
Ahtiainen M, Huttunen P (1999) Long-term effects of forestry managements on water quality and loading in brooks. Boreal Environ Res 4: 101–114.
Alcántara V, Don A, Vesterdal L, Well R, Nieder R (2017) Stability of buried carbon in deep-ploughed forest and cropland soils – implications for carbon stocks. Sci Rep 7, article id 5511. https://doi.org/10.1038/s41598-017-05501-y.
Alcázar J, Woodard PM, Rothwell RL (2002) Soil disturbance and the potential for erosion after mechanical site preparation. North J Appl For 19: 5–13. https://doi.org/10.1093/njaf/19.1.5.
Alriksson A (2001) Regional variability of Cd, Hg, Pb and C concentrations in different horizons of Swedish forest soils. Water Air Soil Pollut Focus 1: 325–341. https://doi.org/10.1023/A:1017589114345.
Aust WM, Blinn CR (2004) Forestry best management practices for timber harvesting and site preparation in the eastern United States: an overview of water quality and productivity research during the past 20 years (1982–2002). Water Air Soil Pollut Focus 4: 5–36. https://doi.org/10.1023/B:WAFO.0000012828.33069.f6.
Bergstedt J, Hagner M, Milberg P (2008) Effects on vegetation composition of a modified forest harvesting and propagation method compared with clear-cutting, scarification and planting. Appl Veg Sci 11: 159–168. https://doi.org/10.3170/2007-7-18343.
Bishop K, Seibert J, Köhler S, Laudon H (2004) Resolving the Double Paradox of rapidly mobilized old water with highly variable responses in runoff chemistry. Hydrol Process 18: 185–189. https://doi.org/10.1002/hyp.5209.
Blackburn M, Ledesma JLJ, Näsholm T, Laudon H, Sponseller RA (2017) Evaluating hillslope and riparian contributions to dissolved nitrogen (N) export from a boreal forest catchment. J Geophys Res-Biogeo 122: 324–339. https://doi.org/10.1002/2016JG003535.
Borrelli P, Schütt B (2014) Assessment of soil erosion sensitivity and post-timber-harvesting erosion response in a mountain environment of central Italy. Geomorphology 204: 412–424. https://doi.org/10.1016/j.geomorph.2013.08.022.
Bringmark L, Lundin L, Augustaitis A, Beudert B, Dieffenbach-Fries H, Dirnböck T, Grabner M-T, Hutchins M, Kram P, Lyulko I, Ruoho-Airola T, Vana M (2013) Trace metal budgets for forested catchments in Europe—Pb, Cd, Hg, Cu and Zn. Water Air Soil Poll 224: 1–14. https://doi.org/10.1007/s11270-013-1502-8.
Cajander AK (1949) Forest types and their significance. Acta For Fenn 56: 1–71. https://doi.org/10.14214/aff.7396.
Cambi M, Certini G, Neri F, Marchi E (2015) The impact of heavy traffic on forest soils: a review. Forest Ecol Manag 338: 124–138. https://doi.org/10.1016/j.foreco.2014.11.022.
Carling PA, Irvine BJ, Hill A, Wood M (2001) Reducing sediment inputs to Scottish streams: a review of the efficacy of soil conservation practices in upland forestry. Sci Total Environ 265: 209–227. https://doi.org/10.1016/S0048-9697(00)00660-4.
de Wit HA, Kvindesland S (1999) Carbon stocks in Norwegian forest soils and effects of forest management on carbon storage. Rapport fra skogforskningen Supplement 14, Norsk institutt for skogforskning. http://hdl.handle.net/11250/2591727.
Egnell G, Jurevics A, Peichl M (2015) Negative effects of stem and stump harvest and deep soil cultivation on the soil carbon and nitrogen pools are mitigated by enhanced tree growth. Forest Ecol Manag 338: 57–67. https://doi.org/10.1016/j.foreco.2014.11.006.
Eklöf K, Schelker J, Sørensen R, Meili M, Laudon H, von Brömssen C, Bishop K (2014) Impact of forestry on total and methyl-mercury in surface waters: Distinguishing effects of logging and site preparation. Environ Sci Technol 48: 4690–4698. https://doi.org/10.1021/es404879p.
Eklöf K, Lidskog R, Bishop K (2016) Managing Swedish forestry’s impact on mercury in fish: defining the impact and mitigation measures. Ambio 45: 163–174. https://doi.org/10.1007/s13280-015-0752-7.
Eriksson O, Raunistola T (1990) Impact of soil scarification on reindeer pastures. Rangifer 10: 99–106. https://doi.org/10.7557/2.10.3.837.
Fölster J (2000) The near-stream zone is a source of nitrogen in a Swedish forested catchment. J Environ Qual 29: 883–893. https://doi.org/10.2134/jeq2000.00472425002900030026x.
Giesler R, Ilvesniemi H, Nyberg L, van Hees P, Starr M, Bishop K, Kareinen T, Lundström US (2000) Distribution and mobilization of Al, Fe and Si in three podzolic soil profiles in relation to the humus layer. Geoderma 94: 249–263. https://doi.org/10.1016/S0016-7061(99)00057-9.
Grabs T, Bishop K, Laudon H, Lyon SW, Seibert J (2012) Riparian zone hydrology and soil water total organic carbon (TOC): implications for spatial variability and upscaling of lateral riparian TOC exports. Biogeosciences 9: 3901–3916. https://doi.org/10.5194/bg-9-3901-2012.
Gundersen V, Clarke N, Dramstad W, Fjellstad W (2016) Effects of bioenergy extraction on visual preferences in boreal forests: a review of surveys from Finland, Sweden and Norway. Scand J Forest Res 31: 323–334. https://doi.org/10.1080/02827581.2015.1099725.
Hagner S (1962) Naturlig föryngring under skärm. [Natural regeneration in shelterwoods]. Medd Stat Skogsforskningsinst 52: 1–263.
Haukenes VL, Åsgård L, Asplund J, Nybakken L, Rolstad J, Storaunet KO, Ohlson M (2022) Spatial variation of surface soil carbon in a boreal forest – the role of historical fires, contemporary vegetation, and hydro-topography. Scand J Forest Res 37: 287–294. https://doi.org/10.1080/02827581.2022.2104364.
Heiskanen J, Saksa T, Luoranen J (2013) Soil preparation method affects outplanting success of Norway spruce container seedlings on till soils susceptible to frost heave. Silva Fenn 47, article id 893. https://doi.org/10.14214/sf.893.
Hildebrand EE (1990) The spatial heterogeneity of chemical properties in acid forest soils and its importance for tree nutrition. In: Zöttl HW, Hüttl RF (eds) Management of nutrition in forests under stress. Proceedings of the International Symposium, Freiburg, Germany, 18–21 September 1989. Springer Netherlands, Dordrecht, pp 183–191.
Hjelm K, Nilsson U, Johansson U, Nordin P (2019) Effects of mechanical site preparation and slash removal on long-term productivity of conifer plantations in Sweden. Can J Forest Res 49: 1311–1319. https://doi.org/10.1139/cjfr-2019-0081.
Högberg P, Wellbrock N, Högberg MN, Mikaelsson H, Stendahl J (2021) Large differences in plant nitrogen supply in German and Swedish forests – implications for management. Forest Ecol Manag 482, article id 118899. https://doi.org/10.1016/j.foreco.2020.118899.
Jandl R, Lindner M, Vesterdal L, Bauwens B, Baritz R, Hagedorn F, Johnson DW, Minkkinen K, Byrne KA (2007) How strongly can forest management influence soil carbon sequestration? Geoderma 137: 253–268. https://doi.org/10.1016/j.geoderma.2006.09.003.
Johansson K, Bergbäck B, Tyler G (2001) Impact of atmospheric long range transport of lead, mercury and cadmium on the Swedish forest environment. Water Air Soil Pollut Focus 1: 279–297. https://doi.org/10.1023/A:1017528826641.
Johansson K, Ring E, Högbom L (2013) Effects of pre-harvest fertilization and subsequent soil scarification on the growth of planted Pinus sylvestris seedlings and ground vegetation after clear-felling. Silva Fenn 47, article id 1016. https://doi.org/10.14214/sf.1016.
Johansson MB (1994) The influence of soil scarification on the turn‐over rate of slash needles and nutrient release. Scand J Forest Res 9: 170–179. https://doi.org/10.1080/02827589409382828.
Johnson DW (1992) Nitrogen retention in forest soils. J Environ Qual 21: 1–12. https://doi.org/10.2134/jeq1992.00472425002100010001x.
Karlsson C, Örlander G (2000) Soil scarification shortly before a rich seed fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scand J Forest Res 15: 256–266. https://doi.org/10.1080/028275800750015073.
Kubin E (1995) The effect of clear cutting, waste wood collecting and site preparation on the nutrient leaching to groundwater. In: Nilsson LO, Hüttl RF, Johansson UT (eds), Nutrient uptake and cycling in forest ecosystems. Proceedings of the CEC/IUFRO Symposium Nutrient uptake and cycling in forest ecosystems, Halmstad, Sweden, 7–10 June 1993. Springer Netherlands, Dordrecht, pp 661–670.
Kubin E (1998) Leaching of nitrate nitrogen into the groundwater after clear felling and site preparation. Boreal Environ Res 3: 3–8.
Kubin E, Kemppainen L (1994) Effect of soil preparation of boreal spruce forest on air and soil temperature conditions in forest regeneration areas. Acta For Fenn 244. https://doi.org/10.14214/aff.7506.
Kuglerová L, Jyväsjärvi J, Ruffing C, Muotka T, Jonsson A, Andersson E, Richardson JS (2020) Cutting edge: a comparison of contemporary practices of riparian buffer retention around small streams in Canada, Finland, and Sweden. Water Resour Res 56, article id e2019WR026381. https://doi.org/10.1029/2019WR026381.
Ledesma JLJ, Grabs T, Futter MN, Bishop KH, Laudon H, Köhler SJ (2013) Riparian zone control on base cation concentration in boreal streams. Biogeosciences 10: 3849–3868. https://doi.org/10.5194/bg-10-3849-2013.
Ledesma JLJ, Futter MN, Blackburn M, Lidman F, Grabs T, Sponseller RA, Laudon H, Bishop KH, Köhler SJ (2018) Towards an improved conceptualization of riparian zones in boreal forest headwaters. Ecosystems 21: 297–315. https://doi.org/10.1007/s10021-017-0149-5.
Lehto J (1956) Tutkimuksia männyn luontaisesta uudistumisesta Etelä-Suomen kangasmailla. [Studies on the natural reproduction of Scots pine on the upland soils of southern Finland]. Acta For Fenn 66. https://doi.org/10.14214/aff.7472.
Lindroos A-J, Ilvesniemi H (2023) Weathering rates of Ca and Mg related to granitic and gabbro mineralogy in boreal forest soils and the effect of mechanical soil disturbance on weathering release. Silva Fenn 57, article id 10648. https://doi.org/10.14214/sf.10648.
Lindroos A-J, Derome K, Piispanen J, Ilvesniemi H (2016) Geochemical changes in podzolic forest soil 17 years after deep tilling. Boreal Environ Res 21: 504–512. http://hdl.handle.net/10138/225809.
Lundin L (1979) Kalhuggningens inverkan på markvattenhalt och grundvattennivå. [The effect of clearcutting on soil moisture and groundwater level]. Rapporter i skogsekologi och skoglig marklära 36, Swedish University of Agricultural Sciences, Uppsala.
Lundmark-Thelin A, Johansson M-B (1997) Influence of mechanical site preparation on decomposition and nutrient dynamics of Norway spruce (Picea abies (L.) Karst.) needle litter and slash needles. Forest Ecol Manag 96: 101–110. https://doi.org/10.1016/S0378-1127(97)00040-6.
Mäkipää R, Abramoff R, Adamczyk B, Baldy V, Biryol C, Bosela M, Casals P, Curiel Yuste J, Dondini M, Filipek S, Garcia-Pausas J, Gros R, Gömöryová E, Hashimoto S, Hassegawa M, Immonen P, Laiho R, Li H, Li Q, Luyssaert S, Menival C, Mori T, Naudts K, Santonja M, Smolander A, Toriyama J, Tupek B, Ubeda X, Johannes Verkerk P, Lehtonen A (2023) How does management affect soil C sequestration and greenhouse gas fluxes in boreal and temperate forests? – A review. Forest Ecol Manag 529, article id 120637. https://doi.org/10.1016/j.foreco.2022.120637.
Mälkönen E (1990) Estimation of nitrogen saturation on the basis of long-term fertilization experiments. Plant Soil 128: 75–82. https://doi.org/10.1007/BF00009398.
Mannerkoski H, Finér L, Piirainen S, Starr M (2005) Effect of clear-cutting and site preparation on the level and quality of groundwater in some headwater catchments in eastern Finland. Forest Ecol Manag 220: 107–117. https://doi.org/10.1016/j.foreco.2005.08.008.
Mayer M, Prescott CE, Abaker WEA, Augusto L, Cécillon L, Ferreira GWD, James J, Jandl R, Katzensteiner K, Laclau J-P, Laganière J, Nouvellon Y, Paré D, Stanturf JA, Vanguelova EI, Vesterdal L (2020) Tamm Review: Influence of forest management activities on soil organic carbon stocks: a knowledge synthesis. Forest Ecol Manag 466, article id 118127. https://doi.org/10.1016/j.foreco.2020.118127.
McMinn JW (1985) Whole-tree harvesting affects pine regeneration and hardwood competition. South J Appl For 9: 81–84. https://doi.org/10.1093/sjaf/9.2.81.
Merilä P, Mustajärvi K, Helmisaari H-S, Hilli S, Lindroos A-J, Nieminen TM, Nöjd P, Rautio P, Salemaa M, Ukonmaanaho L (2014) Above- and below-ground N stocks in coniferous boreal forests in Finland: implications for sustainability of more intensive biomass utilization. Forest Ecol Manag 311: 17–28. https://doi.org/10.1016/j.foreco.2013.06.029.
Mjöfors K, Strömgren M, Nohrstedt H-Ö, Gärdenäs A (2015) Impact of site-preparation on soil-surface CO2 fluxes and litter decomposition in a clear-cut in Sweden. Silva Fenn 49, article id 1403. https://doi.org/10.14214/sf.1403.
Mjöfors K, Strömgren M, Nohrstedt H-Ö, Johansson M-B, Gärdenäs AI (2017) Indications that site preparation increases forest ecosystem carbon stocks in the long term. Scand J Forest Res 32: 717–725. https://doi.org/10.1080/02827581.2017.1293152.
Närhi P, Gustavsson N, Sutinen M-L, Mikkola K, Sutinen R (2013) Long-term effect of site preparation on soil quality in Tuntsa, Finnish Lapland. Geoderma 192: 1–6. https://doi.org/10.1016/j.geoderma.2012.07.007.
Neff EL (1980) Snow trapping by contour furrows in Southeastern Montana. J Range Manage 33: 221–223. https://doi.org/10.2307/3898290.
Newmaster SG, Parker WC, Bell FW, Paterson JM (2007) Effects of forest floor disturbances by mechanical site preparation on floristic diversity in a central Ontario clearcut. Forest Ecol Manag 246: 196–207. https://doi.org/10.1016/j.foreco.2007.03.058.
Nieminen JK, Räisänen M, Haimi J (2012) Spot mounding and granulated wood ash increase inorganic N availability and alter key components of the soil food web in clear-cut Norway spruce forests. Forest Ecol Manag 263: 24–30. https://doi.org/10.1016/j.foreco.2011.09.023.
Nilsson O, Hjelm K, Nilsson U (2019) Early growth of planted Norway spruce and Scots pine after site preparation in Sweden. Scand J Forest Res 34: 678–688. https://doi.org/10.1080/02827581.2019.1659398.
Nohrstedt H-Ö (1990) Effects of repeated nitrogen fertilization with different doses on soil properties in a Pinus sylvestris stand. Scand J Forest Res 5: 3–15. https://doi.org/10.1080/02827589009382588.
Nohrstedt H-Ö (2000) Effects of soil scarification and previous N fertilisation on pools of inorganic N in soil after clear-felling of a Pinus sylvestris (L.) stand. Silva Fenn 34: 195–204. https://doi.org/10.14214/sf.625.
Norberg G, Jäderlund A, Zackrisson O, Nordfjell T, Wardle DA, Nilsson M-C, Dolling A (1997) Vegetation control by steam treatment in boreal forests: a comparison with burning and soil scarification. Can J Forest Res 27: 2026–2033. https://doi.org/10.1139/x97-183.
Nordborg F, Nilsson U, Örlander G (2003) Effects of different soil treatments on growth and net nitrogen uptake of newly planted Picea abies (L.) Karst. seedlings. Forest Ecol Manag 180: 571–582. https://doi.org/10.1016/S0378-1127(02)00650-3.
Nordborg F, Nilsson U, Gemmel P, Örlander G (2006) Carbon and nitrogen stocks in soil, trees and field vegetation in conifer plantations 10 years after deep soil cultivation and patch scarification. Scand J Forest Res 21: 356–363. https://doi.org/10.1080/02827580600976615.
Örlander G, Gemmel P, Hunt J (1990) Site preparation: a Swedish overview. FRDA Report 105. Ministry of Forests, Research Branch, British Columbia.
Örlander G, Egnell G, Albrektson A (1996) Long-term effects of site preparation on growth in Scots pine. Forest Ecol Manag 86: 27–37. https://doi.org/10.1016/S0378-1127(96)03797-8.
Österling ME, Arvidsson BL, Greenberg LA (2010) Habitat degradation and the decline of the threatened mussel Margaritifera margaritifera: influence of turbidity and sedimentation on the mussel and its host. J Appl Ecol 47: 759–768. https://doi.org/10.1111/j.1365-2664.2010.01827.x.
Palviainen M, Finér L, Laurén A, Mannerkoski H, Piirainen S, Starr M (2007) Development of ground vegetation biomass and nutrient pools in a clear-cut disc-plowed boreal forest. Plant Soil 297: 43–52. https://doi.org/10.1007/s11104-007-9317-2.
Palviainen M, Finér L, Laurén A, Launiainen S, Piirainen S, Mattsson T, Starr M (2014) Nitrogen, phosphorus, carbon, and suspended solids loads from forest clear-cutting and site preparation: long-term paired catchment studies from Eastern Finland. Ambio 43: 218–233. https://doi.org/10.1007/s13280-013-0439-x.
Piirainen S, Finér L, Mannerkoski H, Starr M (2007) Carbon, nitrogen and phosphorus leaching after site preparation at a boreal forest clear-cut area. Forest Ecol Manag 243: 10–18. https://doi.org/10.1016/j.foreco.2007.01.053.
Piirainen S, Finér L, Mannerkoski H, Starr M (2009) Leaching of cations and sulphate after mechanical site preparation at a boreal forest clear-cut area. Geoderma 149: 386–392. https://doi.org/10.1016/j.geoderma.2009.01.003.
Piirainen S, Finér L, Starr M (2015) Changes in forest floor and mineral soil carbon and nitrogen stocks in a boreal forest after clear-cutting and mechanical site preparation. Eur J Soil Sci 66: 735–743. https://doi.org/10.1111/ejss.12264.
Porvari P, Verta M, Munthe J, Haapanen M (2003) Forestry practices increase mercury and methyl mercury output from boreal forest catchments. Environ Sci Technol 37: 2389–2393. https://doi.org/10.1021/es0340174.
Pumpanen J, Westman CJ, Ilvesniemi H (2004) Soil CO2 efflux from a podzolic forest soil before and after forest clear-cutting and site preparation. Boreal Environ Res 9: 199–212.
Rappe George MO, Hansson LJ, Ring E, Jansson PE, Gärdenäs AI (2017) Nitrogen leaching following clear-cutting and soil scarification at a Scots pine site – a modelling study of a fertilization experiment. Forest Ecol Manag 385: 281–294. https://doi.org/10.1016/j.foreco.2016.11.006.
Ring E (1996) Effects of previous N fertilizations on soil-water pH and N concentrations after clear-felling and soil scarification at a Pinus sylvestris site. Scand J Forest Res 11: 7–16. https://doi.org/10.1080/02827589609382907.
Ring E, Jacobson S, Högbom L (2011) Long-term effects of nitrogen fertilization on soil chemistry in three Scots pine stands in Sweden. Can J Forest Res 41: 279–288. https://doi.org/10.1139/X10-208.
Ring E, Högbom L, Jansson G (2013) Effects of previous nitrogen fertilization on soil-solution chemistry after final felling and soil scarification at two nitrogen-limited forest sites. Can J Forest Res 43: 396–404. https://doi.org/10.1139/cjfr-2012-0380.
Ring E, Högbom L, Jacobson S, Jansson G, Nohrstedt H-Ö (2018) Long-term effects on soil-water nitrogen and pH of clearcutting and simulated disc trenching of previously nitrogen-fertilised pine plots. Can J Forest Res 48: 1115–1123. https://doi.org/10.1139/cjfr-2018-0042.
Ring E, Johansson F, von Brömssen C, Bergkvist I (2022) A snapshot of forest buffers near streams, ditches, and lakes on forest land in Sweden – lessons learned. Silva Fenn 56, article id 10676. https://doi.org/10.14214/sf.10676.
Rosén K, Aronson J-A, Eriksson HM (1996) Effects of clear-cutting on streamwater quality in forest catchments in central Sweden. Forest Ecol Manag 83: 237–244. https://doi.org/10.1016/0378-1127(96)03718-8.
Roturier S, Bergsten U (2006) Influence of soil scarification on reindeer foraging and damage to planted Pinus sylvestris seedlings. Scand J Forest Res 21: 209–220. https://doi.org/10.1080/02827580600759441.
Roturier S, Bäcklund S, Sundén M, Bergsten U (2007) Influence of ground substrate on establishment of reindeer lichen after artificial dispersal. Silva Fenn 41: 269–280. https://doi.org/10.14214/sf.296.
Roturier S, Sundén M, Bergsten U (2011) Re-establishment rate of reindeer lichen species following conventional disc trenching and HuMinMix soil preparation in Pinus-lichen clear-cut stands: a survey study in northern Sweden. Scand J Forest Res 26: 90–98. https://doi.org/10.1080/02827581.2010.528019.
Rumpel C, Kögel-Knabner I (2011) Deep soil organic matter – a key but poorly understood component of terrestrial C cycle. Plant Soil 338: 143–158. https://doi.org/10.1007/s11104-010-0391-5.
Saksa T, Miina J, Haatainen H, Kärkkäinen K (2018) Quality of spot mounding performed by continuously advancing mounders. Silva Fenn 52, article id 9933. https://doi.org/10.14214/sf.9933.
Salmivaara A, Leinonen A, Palviainen M, Korhonen N, Launiainen S, Tuomenvirta H, Ukonmaanaho L, Finér L, Laurén A (2023) Exploring the role of weather and forest management on nutrient export in boreal forested catchments using spatially distributed model. Forests 14, article id 89. https://doi.org/10.3390/f14010089.
Šamonil P, Daněk P, Schaetzl RJ, Tejnecký V, Drábek O (2018) Converse pathways of soil evolution caused by tree uprooting: a synthesis from three regions with varying soil formation processes. Catena 161: 122–136. https://doi.org/10.1016/j.catena.2017.09.032.
Schelker J, Eklöf K, Bishop K, Laudon H (2012) Effects of forestry operations on dissolved organic carbon concentrations and export in boreal first-order streams. J Geophys Res-Biogeo 117, article id G01011. https://doi.org/10.1029/2011JG001827.
Sikström U, Hökkä H (2016) Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance. Silva Fenn 50, article id 1416. https://doi.org/10.14214/sf.1416.
Sikström U, Hjelm K, Holt Hanssen K, Saksa T, Wallertz K (2020) Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fenn 54, article id 10172. https://doi.org/10.14214/sf.10172.
Skoklefald S (1995) Spot scarification in a mountainous Scots pine forest in Norway. The Finnish Forest Research Institute Research Papers 567: 85–90. http://urn.fi/URN:ISBN:951-40-1466-9.
Smolander A, Heiskanen J (2007) Soil N and C transformations in two forest clear-cuts during three years after mounding and inverting. Can J Soil Sci 87: 251–258. https://doi.org/10.4141/S06-028.
Smolander A, Paavolainen L, Mälkönen E (2000) C and N transformations in forest soil after mounding for regeneration. Forest Ecol Manag 134: 17–28. https://doi.org/10.1016/S0378-1127(99)00242-X.
Sponseller RA, Gundale MJ, Futter M, Ring E, Nordin A, Nasholm T, Laudon H (2016) Nitrogen dynamics in managed boreal forests: recent advances and future research directions. Ambio 45: 175–187. https://doi.org/10.1007/s13280-015-0755-4.
Strömgren M, Mjöfors K (2012) Soil-CO2 flux after patch scarification, harrowing and stump harvest in a hemi-boreal forest. Scand J Forest Res 27: 754–761. https://doi.org/10.1080/02827581.2012.723741.
Strömgren M, Mjöfors K, Holmström B, Grelle A (2012) Soil CO2 flux during the first years after stump harvesting in two Swedish forests. Silva Fenn 46: 67–79. https://doi.org/10.14214/sf.66.
Strömgren M, Hedwall PO, Olsson BA (2016) Effects of stump harvest and site preparation on N2O and CH4 emissions from boreal forest soils after clear-cutting. Forest Ecol Manag 371: 15–22. https://doi.org/10.1016/j.foreco.2016.03.019.
Strömgren M, Mjöfors K, Olsson BA (2017) Soil-surface CO2 flux during the first 2 years after stump harvesting and site preparation in 14 Swedish forests. Scand J Forest Res 32: 213–221. https://doi.org/10.1080/02827581.2016.1221993.
Sutinen R, Pänttäjä M, Teirilä A, Sutinen M-L (2006) Effect of mechanical site preparation on soil quality in former Norway spruce sites. Geoderma 136: 411–422. https://doi.org/10.1016/j.geoderma.2006.04.018.
Sutinen R, Närhi P, Herva H, Piekkari M, Sutinen M-L (2010) Impact of intensive forest management on soil quality and natural regeneration of Norway spruce. Plant Soil 336: 421–431. https://doi.org/10.1007/s11104-010-0492-1.
Sutinen R, Gustavsson N, Hänninen P, Middleton M, Räisänen ML (2019) Impact of mechanical site preparation on soil properties at clear-cut Norway spruce sites on mafic rocks of the Lapland Greenstone Belt. Soil Till Res 186: 52–63. https://doi.org/10.1016/j.still.2018.10.013.
Sutton RF (1993) Mounding site preparation: a review of European and North American experience. New Forest 7: 151–192. https://doi.org/10.1007/BF00034198.
Swedish Forest Agency (2022) Skogsvårdslagstiftningen, Gällande regler 1 april 2022. [Forestry legislation, Rules in force 1 April 2022].
Swedish National Forest Inventory (2022) Unpublished data. Department of Forest Resource Management, Swedish University of Agricultural Sciences, Umeå. https://www.slu.se/riksskogstaxeringen.
Tamm CO, Holmen H, Popovíc B, Wiklander G (1974) Leaching of plant nutrients from soils as a consequence of forestry operations. Ambio 3: 211–221.
Tanskanen N, Ilvesniemi H (2004) The amount of secondary Al in two ploughed podzolic forest soils. Geoderma 119: 249–260. https://doi.org/10.1016/j.geoderma.2003.08.003.
Tanskanen N, Ilvesniemi H (2007) Spatial distribution of fine roots at ploughed Norway spruce forest sites. Silva Fenn 41: 45–54. https://doi.org/10.14214/sf.306.
Tanskanen N, Kareinen T, Nissinen A, Ilvesniemi H (2004) Soil solution aluminium in disturbed and undisturbed podzolic profiles at two tilt-ploughed forest sites. Boreal Environ Res 9: 347–355.
Thomsen LM, Baartman JEM, Barneveld RJ, Starkloff T, Stolte J (2015) Soil surface roughness: comparing old and new measuring methods and application in a soil erosion model. SOIL 1: 399–410. https://doi.org/10.5194/soil-1-399-2015.
Tonteri T, Hallikainen V, Merila P, Miina J, Rautio P, Salemaa M, Tolvanen A (2022) Response of ground macrolichens to site factors, co-existing plants and forestry in boreal forests. Appl Veg Sci 25, article id e12690. https://doi.org/10.1111/avsc.12690.
Valtera M, Schaetzl RJ (2017) Pit-mound microrelief in forest soils: review of implications for water retention and hydrologic modelling. Forest Ecol Manag 393: 40–51. https://doi.org/10.1016/j.foreco.2017.02.048.
Vanha-Majamaa I, Shorohova E, Kushnevskaya H, Jalonen J (2017) Resilience of understory vegetation after variable retention felling in boreal Norway spruce forests – a ten-year perspective. Forest Ecol Manag 393: 12–28. https://doi.org/10.1016/j.foreco.2017.02.040.
Wallertz K, Björklund N, Hjelm K, Petersson M, Sundblad L-G (2018) Comparison of different site preparation techniques: quality of planting spots, seedling growth and pine weevil damage. New Forest 49: 705–722. https://doi.org/10.1007/s11056-018-9634-8.
Wei X, Giles-Hansen K, Spencer SA, Ge X, Onuchin A, Li Q, Burenina T, Ilintsev A, Hou Y (2022) Forest harvesting and hydrology in boreal forests: under an increased and cumulative disturbance context. Forest Ecol Manag 522, article id 120468. https://doi.org/10.3390/f10111006.
Weihermüller L, Siemens J, Deurer M, Knoblauch S, Rupp H, Göttlein A, Pütz T (2007) In situ soil water extraction: a review. J Environ Qual 36: 1735–1748. https://doi.org/10.2134/jeq2007.0218.
Wennström U, Bergsten U, Nilsson J-E (1999) Mechanizsed microsite preparation and direct seeding of Pinus sylvestris in boreal forests – a way to create desired spacing at low cost. New Forest 18: 179–198. https://doi.org/10.1023/A:1006506431344.
Total of 117 references.
