Henrik Lindberg  ,
Tuomas Aakala,
Ilkka Vanha-Majamaa
,
Tuomas Aakala,
Ilkka Vanha-Majamaa
Ignition probability and fuel consumption of boreal ground vegetation fuels – an experimental study in Finland
Lindberg H., Aakala T., Vanha-Majamaa I. (2024). Ignition probability and fuel consumption of boreal ground vegetation fuels – an experimental study in Finland. Silva Fennica vol. 58 no. 3 article id 23064. https://doi.org/10.14214/sf.23064
Highlights
- Ignition probability and mass loss rates clearly differed in experimental burnings among four common circumboreal moss and lichen species
- Cladonia rangiferina was the most flammable, Dicranum spp. the least flammable and Pleurozium schreberi and Hylocomium splendens intermediate.
Abstract
In boreal forests fires often ignite and spread within the dominant moss and lichen cover of the ground layer vegetation, which thus greatly influences fire hazard. We used an experimental set-up in greenhouse conditions to study the differences in how (1) fuel moisture and (2) wind velocity influence the ignition probability and fuel consumption among four common circumboreal ground vegetation fuels, Pleurozium schreberi (Willd. ex Brid.) Mitt., Hylocomium splendens Schimp., Dicranum spp. and Cladonia rangiferina (L.) F. H. Wigg. Our results show that the reindeer lichen C. rangiferina was clearly the most flammable species, with high ignition probability even at high moisture contents and low wind velocities. Of the mosses, Dicranum was the least flammable, with low ignition probability and mass loss at low wind velocities regardless of moisture content. P. schreberi and H. splendens behaved somewhat similarly with wind velocities quickly increasing the initially low ignition probability and mass loss observed in the absence of wind. However, especially for mass loss, among-species differences tended to disappear with stronger winds. The observed differences can be explained by the different structures and growth forms of the studied species and open a potential avenue for improving forest fire risk predictions.
Keywords
forest fires;
ground vegetation;
prescribed burning;
flammability;
fuel moisture content;
mass loss rate
-
Lindberg,
Häme University of Applied Sciences, School of Bioeconomy, Visamäentie 35 A, P.O. Box 230, FI-13100 Hämeenlinna, Finland
 https://orcid.org/0000-0001-9045-2673
E-mail
henrik.lindberg@hamk.fi
https://orcid.org/0000-0001-9045-2673
E-mail
henrik.lindberg@hamk.fi
-
Aakala,
University of Eastern Finland, School of Forest Sciences, P.O. Box 111, FI-80101 Joensuu, Finland
https://orcid.org/0000-0003-0160-6410
E-mail
tuomas.aakala@uef.fi
- Vanha-Majamaa, Natural Resources Institute Finland (Luke), Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail ilkka.vanha-majamaa@luke.fi
Received 24 October 2023 Accepted 22 April 2024 Published 30 April 2024
Views 42832
Available at https://doi.org/10.14214/sf.23064 | Download PDF

1 Introduction
Until recent years, the forest fire hazard in Fennoscandia has been low compared to many other parts of the boreal zone, but recent large fires in Sweden (Gustafsson et al. 2019; Granström 2020) have forced a re-evaluation of the risk and significance of forest fires also in Fennoscandia. At the same time, the effective fire suppression has led to a decline of fire-associated biodiversity (Granström 2001), hence conservation of burned areas and increasing prescribed burning are considered necessary to safeguard fire-related habitats and species.
Both the potential risk of increasing impact of forest fires in the boreal zone and the need to maintain fire-related habitats require developing fire management methods. The flammability characteristics of fuels are key factors in predictions of fire ignition and spread (Ryan 2002). The fuel moisture content (FMC) is known to play a pivotal role in the ignition, fire behavior and fuel consumption during a fire, and can be considered as one of the most important inputs in forest fire prediction models (Keane 2015). Although somewhat less studied, wind velocity also affects the flammability of forest fuels by transporting oxygen and heat, as well as tilting the flame that further enhances heat transfer (Varner et al. 2015).
In many parts of the boreal zone, a continuous moss and lichen cover consisting mostly of a few circumboreal species forms the surface part of duff layer, where forest fires typically ignite and spread (Schimmel and Granström 1997; Tanskanen et al. 2005). The feather moss (Pleurozium schreberi (Willd. ex Brid.) Mitt.) is the most abundant in many areas of white (Picea glauca (Moench) Voss) and black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.) dominated forests of North America, Norway spruce (Picea abies (L.) H. Karst.), Scots pine (Pinus sylvestris L.), and larch (Larix) dominated forests in boreal Eurasia (Tesky 1992; Babintseva and Titova 1996; Mäkipää 2000; Ivanova et al. 2020). Pleurozium schreberi often shares dominance with the stairstep moss (Hylocomium splendens Schimp.), and fork mosses (Dicranum), that have in recent decades increased in abundance in Fennoscandia (Mäkipää and Heikkinen 2003). The reindeer lichens (Cladonia) are abundant in drier and xeric sites (Nousiainen 2000).
Although mosses and lichens are living organisms, they can be considered to behave similarly to fine, dead fuels, since they have no internal water holding or transporting organs, and thus dry rapidly (Tanskanen et al. 2006). Yet, these species can vary greatly in the amount of moisture they contain, because of their tissue material, stem structure and growth form (Tanskanen et al. 2006; Lindberg et al. 2021). Compared to mosses, reindeer lichens are known to have lower bulk density, lower surface-to-volume ratio, and lower heat content (Munger 2008). Of the moss species of our study Dicranum forms denser growth including thick tomentum (Peterson and Mayo 1975) which is shown in higher bulk density when compared with H. splendens and P. schreberi (Grau-Andrés et al. 2022).
These properties also influence air flow and can be expected to modify the influence of wind on the ignition and spread of fires. Forest floors with different moss and lichen community composition are thus known to have different drying rates (Ivanova et al. 2020; Lindberg et al. 2021), affecting their flammability. It is possible that the species also differ in their flammability even with similar FMC values and wind conditions because of their structural differences. In this experimental study we examined the impact of FMC and wind velocity on the ignition probability and fuel consumption of above-mentioned common moss and lichen species. Since the study was done in controlled greenhouse conditions our aim was to find out possible general patterns and relative differences between the studied materials. Thus, the focus of the study was not to define threshold values directly applicable in field conditions.
2 Materials and methods
2.1 Sampling and experimental set-up
Undisturbed samples of four common circumboreal moss and lichen species of mature mineral-soil forests were collected from a mature Scots pine-dominated semi-xeric Vaccinium-type forest stand of approximately one hectare in size in Evo state forest in Hämeenlinna, Southern Finland (61°12´N, 25°05´E). The stand was 55 years old, with a standing volume of 180 m3 ha–1, and average tree height of 19.5 m. Stand structure was typical to a Finnish managed stand with an evenly aged structure and practically absent understory, as a result of several pre-commercial and commercial thinnings. The stand had a characteristic raw humus layer of ~4 cm which is typical to semi-xeric sites in Finland and a continuous ground layer where moss and lichen species dominance varies in patches.
The four collected species were those that are in general the most abundant in mesic and semi-xeric forests in Finland: Dicranum spp., mainly D. polysetum Sw. ex anon, with possible marginal amounts of other Dicranum species, P. schreberi, H. splendens and Cladonia rangiferina (L.) F. H. Wigg. We randomly sampled monospecific patches of each target species from uniform moss-lichen carpet, so they represented similar substrates and micro-locations. The samples were separated from the raw humus layer carefully by hand and thus included only the loose living growth (height 5–10 cm) of species, retained their original growth structure and contained no raw humus nor litter.
Samples were placed in aluminum trays (110 mm × 84 mm × 41mm) and were oven-dried in 105 °C temperature for 18 hours to measure their dry mass. They were then fully moistened with 100 milliliters of water, which equals a rainfall of approximately 10 millimeters and wets the moss or lichen layer thoroughly. After this they were dried to target dry weight FMC values of 11% (tolerance ± 0.9%), 25% (± 0.9%), 43% (±1.1%), 67% (±2.0%) and 100% (±2.0%). The used FMC values were chosen so that they covered a wide range of moisture contents, to ensure that the shape of the response and differences among the species were captured.
Immediately after the targeted FMC were reached the samples were placed on a table in a greenhouse, where three different wind velocities (0, 1, 2 m s–1) were created by a fan. Since this was an experimental study, we used rather low velocities to analyze the sensitivity of ignition to even small changes in wind velocity. For each combination of FMC value, wind velocity and species, at least five replicate samples were targeted (Table 1). However, as several samples were contaminated by mold, the targeted burning of five replicates could not be achieved in all cases (Table 1). Especially for Dicranum, experiments with 25% and 43% FMC values and 0m s–1 wind velocity were reduced because of this. (Table 1). The air humidity and temperature could not be fully controlled but the ignition tests were timed on circumstances when air relative humidity was observed to be between 40% and 60% and temperature over 20 °C. The ignition potential of similar fuels has been previously studied by match ignition trials in field experiments in Canada (Lawson et al. 1993), Sweden (Granström and Schimmel 1998) and Finland (Tanskanen et al. 2005). In our study we adapted similar point-ignition source method, yet instead of matches the samples were ignited by keeping a long tube open-flame gas igniter for 30 seconds in the center of the lower quarter (relative to wind direction). After the potential burning was completed, the percentage of burned area was determined and samples were dried and weighed to calculate the mass loss.
| Table 1. The number of ignition tests performed in the study by species, moisture content and wind velocity. Total number of tests: 293, location of the study: Evo state forest, Hämeenlinna, Finland. | |||||
| Wind velocity 0 m s–1 | |||||
| Moisture content | 11% | 25% | 43% | 67% | 100% |
| Species | |||||
| Pleurozium schreberi | 6 | 5 | 5 | 5 | 5 |
| Dicranum spp. | 5 | -^ | 2 | 5 | 5 |
| Hylocomium splendens | 5 | 4 | 5 | 5 | 5 |
| Cladonia rangiferina | 5 | 5 | 5 | 5 | 5 |
| Wind velocity 1 m s–1 | |||||
| Pleurozium schreberi | 5 | 4 | 5 | 5 | 4 |
| Dicranum spp. | 5 | 4 | 3 | 7 | 4 |
| Hylocomium splendens | 5 | 7 | 7 | 5 | 4 |
| Cladonia rangiferina | 3 | 5 | 6 | 6 | 6 |
| Wind velocity 2 m s–1 | |||||
| Pleurozium schreberi | 6 | 5 | 4 | 6 | 5 |
| Dicranum spp. | 5 | 8 | 4 | 4 | 5 |
| Hylocomium splendens | 5 | 5 | 5 | 5 | 5 |
| Cladonia rangiferina | 6 | 5 | 5 | 4 | 4 |
| ^) All samples in this group were contaminated by mold and omitted from analysis | |||||
Flammability is most often understood as the general ability of a fuel to ignite and combust in the absence of ignition source (Varner et al. 2015), so in our study all samples with successful ignitions were considered flammable. In this study no separation to smoldering and flaming combustions was made (see Larjavaara 2005).
2.2 Data analysis
Ignition was considered successful when ≥ 90% of the surface area was burned by flaming combustion and unsuccessful when < 90% was burned. The 90% threshold was chosen because in some cases small, unburned patches were left in the corners of the test trays. Ignition probability was then modelled as a function of FMC and wind speed (as a factorial variable), using logistic regression modelling. Models were fit separately for each species.
Fuel consumption was similarly modelled as a function of FMC and wind, separately for each species. Exploratory analyses showed that different-shaped non-linear relationships between FMC and fuel consumption were common, and thus generalized additive models (GAMs) were used to allow for differences in the shape of the response. Fuel consumption was modelled as a smooth function of FMC, with wind as a factorial variable. For strictly positive data (fuel consumed), a Gaussian error distribution with a log-link was used. The spline smoothers were constrained to be monotonically decreasing to prevent the biologically unrealistic increase in fuel consumption with moisture content, which occurred in few cases. This was visually judged to originate from high variation in fuel consumed at low wind speeds. Statistical analyses were conducted in R (version 3.5), using the package scam for the GAM models (Pya 2018).
3 Results
We identified clear differences in both ignition probability (Fig. 1, Table 2) and fuel consumption (Fig. 2, Table 3) among the species. The lichen C. rangiferina was the most flammable fuel with 100% ignition probability up to second highest moisture content tested (67%). Even in the 100% FMC, ignition probability stayed clearly over 50% (Fig. 1). Opposite to the mosses, C. rangiferina’s ignition probability was also almost independent of wind velocity, as response was shown only with the highest FMC values (Fig. 1).
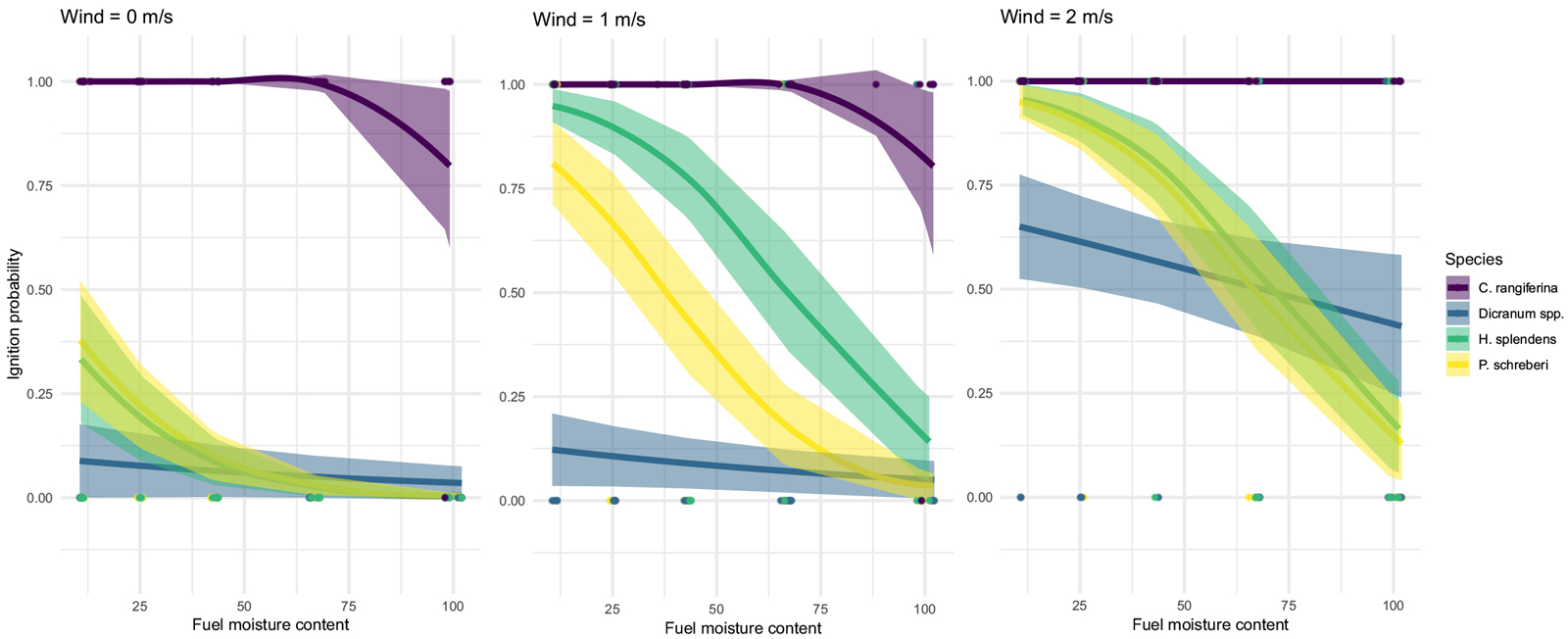
Fig. 1. Ignition probabilities of the studied species by wind velocity and fuel moisture content (%, dry weight basis) of used logistic regression models. Shaded areas show the standard errors of each fitted model and dots show the original measured observations. View larger in new window/tab.
| Table 2. Coefficients of the logistic regression model (ignition probability analysis). | ||||
| Species | Variable | Estimate | Std. error | p |
| Cladonia rangiferina | Intercept | 14.08 | 13.22 | 0.28 |
| Moisture% | –0.13 | 0.14 | 0.34 | |
| Wind 1 m s–1 | 0.35 | 1.59 | 0.83 | |
| Wind 2 m s–1 | 18.26 | 4715.29 | 1 | |
| Dicranum spp. | Intercept | 2.22 | 1.14 | 0.05 |
| Moisture% | –0.01 | 0.01 | 0.30 | |
| Wind 1 m s–1 | 0.36 | 1.27 | 0.78 | |
| Wind 2 m s–1 | 2.95 | 1.11 | 0.01** | |
| Hylocomium splendens | Intercept | –0.14 | 0.75 | 0.86 |
| Moisture% | –0.05 | 0.01 | <0.001*** | |
| Wind 1 m s–1 | 3.60 | 0.96 | <0.001*** | |
| Wind 2 m s–1 | 3.78 | 0.99 | <0.001*** | |
| Pleurozium schreberi | Intercept | 0.06 | 0.68 | 0.92 |
| Moisture% | –0.05 | 0.01 | <0.001*** | |
| Wind 1 m s–1 | 1.95 | 0.81 | 0.016** | |
| Wind 2 m s–1 | 3.45 | 0.92 | <0.001*** | |
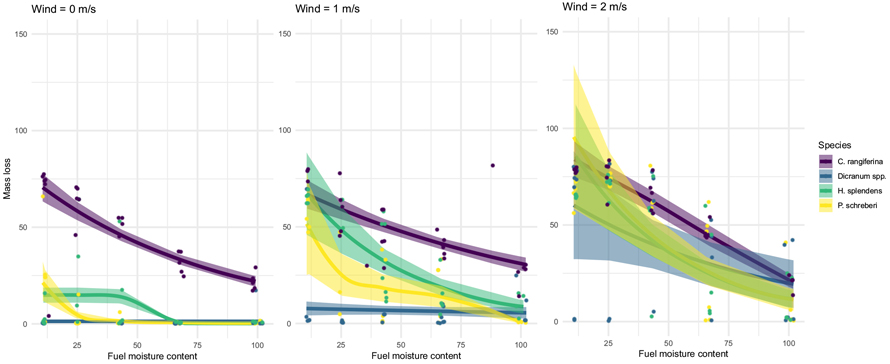
Fig. 2. Mass losses (%, dry weight) of the studied species by wind velocity and fuel moisture content (%, dry weight basis) of used generalized additive models. Shaded areas show the standard errors of each fitted model and dots show the original measured observations. View larger in new window/tab.
| Table 3. Parametric coefficients and significances of smoother terms of generalized additive model (mass loss analysis). Edf: efficient degrees of freedom. | |||||
| Species | Variable/Smooth term | Estimate | Std.error | p | edf |
| Cladonia rangiferina | Parametric coefficients | ||||
| Intercept | 4.43 | 0.12 | <0.001*** | ||
| Wind 1 m s–1 | –0.11 | 0.18 | 0.54 | ||
| Wind 2 m s–1 | 0.11 | 0.21 | 0.60 | ||
| Smooth terms | |||||
| s(moisture): wind 0 m s–1 | <0.001*** | 1.03 | |||
| s(moisture): wind 1 m s–1 | <0.001*** | 0.98 | |||
| s(moisture): wind 2 m s–1 | <0.001*** | 1.80 | |||
| Dicranum spp. | Parametric coefficients | ||||
| Intercept | 0.27 | 0.38 | 0.48 | ||
| Wind 1 m s–1 | 1.83 | 0.67 | 0.008** | ||
| Wind 2 m s–1 | 3.99 | 0.68 | <0.001*** | ||
| Smooth terms | |||||
| s(moisture): wind 0 m s–1 | ̶̶ | 0 | |||
| s(moisture): wind 1 m s–1 | 0.66 | 0.65 | |||
| s(moisture): wind 2 m s–1 | 0.21 | 1.11 | |||
| Hylocomium splendens | Parametric coefficients | ||||
| Intercept | 2.70 | 0.26 | <0.001*** | ||
| Wind 1 m s–1 | 1.80 | 0.47 | <0.001*** | ||
| Wind 2 m s–1 | 2.06 | 0.47 | <0.001*** | ||
| Smooth terms | |||||
| s(moisture): wind 0 m s–1 | <0.001*** | 1.00 | |||
| s(moisture): wind 1 m s–1 | <0.001*** | 0.93 | |||
| s(moisture): wind 2 m s–1 | 0.001** | 1.11 | |||
| Pleurozium schreberi | Parametric coefficients | ||||
| Intercept | 4.81 | 1.44 | 0.001** | ||
| Wind 1 m s–1 | 0.04 | 2.44 | 0.99 | ||
| Wind 2 m s–1 | 0.05 | 1.52 | 0.97 | ||
| Smooth terms | |||||
| s(moisture): wind 0 m s–1 | <0.001*** | 2.38 | |||
| s(moisture): wind 1 m s–1 | <0.001*** | 3.20 | |||
| s(moisture): wind 2 m s–1 | 0.05* | 1.24 | |||
For all three moss species, the absence of wind led to low ignition probability. Even with the driest treatment, the ignition probability remained below 50%. Among moss species, Dicranum was clearly the least flammable (Fig. 1).
Increasing wind had a significant effect on the ignition probability of all studied mosses. P. schreberi and H. splendens showed higher ignition probability already with the 1 m s–1 wind velocity, since over 50% of ignitions were predicted for FMC values under ~40% for P. schreberi, and under ~65% for H. splendens, whereas Dicranum stayed well below the 50% probability even with lowest FMC values. In the highest wind velocity (2 m s–1), all moss species ignited with 50% probability in FMC values lower than ~70%, but P. schreberi and H. splendens showed similarly higher probabilities than Dicranum, especially with lower FMC values (Fig. 1). Compared to C. rangiferina the moss species showed clearly more stochastic ignition behaviour.
In mass loss curves (Fig. 2), homologous behavior among species was apparent. Mass loss curves for C. rangiferina were almost linear, with a minor impact of wind velocity showing that fuel consumption was mainly dependent on FMC. Wind affected the mass loss of moss species clearly and in similar pattern as with ignition probabilities: H. splendens and P. schreberi reached higher mass losses already with 1 m s–1 velocity. Dicranum`s mass loss increased only at the highest wind velocity (2 m s–1, Fig. 2). It is notable that with 2 m s–1 velocity the shapes and values of mass loss curves among species were starting to resemble each other, yet among the moss species the mass loss values were slightly lower and deviation higher than for C. rangiferina (Fig. 2).
4 Discussion
Our results are consistent with previous studies characterizing Cladonia as one of the most flammable forest fuels due to its high surface-to-volume ratio, gelatinous thallus, lack of rhizoids and loose growth form (Pech 1991; Munger 2008; Ivanova et al. 2020). This also possibly explains why the wind velocity had a minor impact on the ignition probability of C. rangiferina, since compared to mosses, the airy stem structure transfers heat and oxygen more rapidly after ignition.
The studied mosses showed different ignition probabilities and clear responses to wind velocity. Dicranum was the least flammable with practically no ignitions with wind velocities of 0and 1 m s–1 but showed a notable increase in ignition probability with 2 m s–1 wind velocity. This is probably explained by the compact, mattress-like growth form and dense tomentum of D. polysetum, which hinders the transfer of oxygen and heat more efficiently than the looser P. schreberi and H. splendens, which showed increasing ignition probabilities already with 1 m s–1 wind velocity. Although comparable experiments with these species are lacking, studies with other fine surface fuels support the impact of wind on flaming ignition (Dimitrakopoulos et al. 2010; Marino et al. 2010; Masinda et al. 2020). In our study increasing wind velocity increased the ignition probability in general and lowered the differences in flammability. Thus, with increasing wind speeds, species identity becomes irrelevant. It must be noted that since we examined the studied materials by their entire living growth the moisture gradient within the growth was not taken into account. The surface layer of mosses is known to dry more rapidly than the denser lower parts, thus possibly enabling ignitions of the surface layer even if the FMC of the entire growth would be too high for ignition.
The correlation with FMC and mass loss of mosses and lichens in fires has been reported in several studies (Hille and den Ouden 2005; Hille and Stephens 2005; Ivanova et al. 2020). Earlier studies have considered FMC values below 30% to result in complete consumption and values of 30–120% in partial reduction (Hille and Stephens 2005). In their laboratory experiment Blauw et al. (2015) reported mass loss rates of >80% and ~60% with respective FMCs of 10% and 30%. In our study, mass loss of C. rangiferina was in line with these principles, as the reduction was ~75% with the lowest FMC (11%) and decreased almost linearly to ~25% with the highest FMC value (100%).
Our results showed low ignition probabilities in the absence of wind for all moss species, regardless of moisture conditions. Even when considering differences in how ignitions are defined, the ignition probabilities were clearly lower than the near 30% moisture content considered as ignition threshold for dead fuels (Keane 2015). In laboratory tests Blauw et al (2015) found both 10% and 30% FMC samples of P. schreberi ignitable. In different field tests, earlier studies have reported clearly higher (e.g., 25–35%; Granström and Schimmel 1998; Tanskanen et al. 2005) FMC values below which ignitions increase, whereas Ivanova et al. (2020) present a range of 10–40% for mosses and 25–40% for lichens based on studies in Russia. The low ignition probabilities in our experiment are likely partially explained by the rather high relative humidity conditions on which the test burnings were performed as FMC of lichens and mosses are influenced rapidly by the atmospheric relative humidity (Norum 1982; Pech 1989). In our experiment the relative humidity varied between 40% and 60%, whereas in their laboratory study Blauw et al. (2015) maintained a clearly lower air humidity of 20–40%. In our experiment we studied flaming combustion with open-flame ignition tests, so the results cannot be adapted to smouldering combustion, which can occur in clearly higher FMC values (Frandsen 1997).
5 Conclusions
The studied species differed notably in their ignition probability and mass loss rates, in a manner that was consistent with differences in their morphology and compactness. Also, wind velocity had a clear impact on the ignition probabilities of mosses and on mass loss of all studied species. Importantly, increasing wind speed seems to even out among-species differences relative to ignition or mass loss at a given moisture content. The results indicate potential for improving fire weather indices by providing more species-specific information as well as helping to predict the burn depth e.g. in prescribed burnings to better achieve the targeted goals.
Major changes in fire regimes in the European boreal region are often explained by transitions from spruce-dominated to pine-dominated forests (and vice versa) (Tryterud 2003; Ohlson et al. 2011), which can be partially due to the lichen/moss ratio. Our results support this hypothesis since there were major differences in the ignition probabilities of C. rangiferina and mosses which, combined to their different moisture behavior result in notable differences in their flammability. Finally, it must be emphasized that the results of this experimental study are not directly applicable to field conditions but form a basis for further developing models for ignitions and fuel consumption, based on moisture content, wind, and species characteristics.
Abbreviations
FMC: Fuel moisture content.
GAM: Generalized additive model.
Data availability statement
The data of this article will be shared on reasonable request to the corresponding author.
Authors’ contribution
HL and I V-M presented the original idea and designed the experiment. HL coordinated the data collecting and data processing. TA performed the statistical analysis. HL prepared the first version of manuscript and all authors participated in finalizing the manuscript.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Acknowledgements
Tomi Hulmi, Antti Kujala and Tuija Toivonen are thanked for their excellent field work and Lammi Biological Station for providing us drying equipment.
Funding
The data collection of study was partially funded by European Union Fifth Framework projects SPREAD and EUFIRELAB.
References
Babintseva RM, Titova YV (1996) Effects of fire on the regeneration of larch forests in the Lake Baikal basin. In: Goldammer JG, Furyaev VV (eds) Fire in ecosystems of boreal Eurasia. Kluwer Academic Publishers, Netherlands, pp 358–365. https://doi.org/10.1007/978-94-015-8737-2_31.
Blauw LG, Wensink N, Bakker L, van Logtestijn RS, Aerts R, Soudzilovskaia NA, Cornelissen JH (2015) Fuel moisture content enhances nonadditive effects of plant mixtures on flammability and fire behavior. Ecol Evol 5: 3830–3841. https://doi.org/10.1002/ece3.1628.
Dimitrakopoulos A, Mitsopoulos I, Gatoulas K (2010) Assessing ignition probability and moisture of extinction in a Mediterranean grass fuel. Int J Wildland Fire 19: 29–34. https://doi.org/10.1071/WF08124.
Frandsen WH (1997) Ignition probability of organic soils. Can J For Res 27: 1471–1477. https://doi.org/10.1139/x97-106.
Granström A (2001) Fire management for biodiversity in the European boreal forest. Scand J Forest Res 16: 62–69. https://doi.org/10.1080/028275801300090627.
Granström A (2020) Brandsommaren 2018: Vad hände, och varför? [Fire summer 2018: What happened and why?]. Myndigheten för Samhällsskydd och Beredskap, Karlstad, Sverige, MSB1496. https://rib.msb.se/filer/pdf/29059.pdf.
Granström A, Schimmel J (1998) Utvärdering av det kanadensiska brandrisksystemet – testbränningar och uttorkningsanalyser. [Assessment of the Canadian forest fire danger rating system for Swedish fuel conditions]. Rescue Service, Karlstad, Sweden, P21–244/98.
Grau-Andrés R, Kardol P, Gundale MJ (2022) Trait coordination in boreal mosses reveals a bryophyte economics spectrum. J Ecol 110: 2493–2506. https://doi.org/10.1111/1365-2745.13965.
Gustafsson L, Berglind M, Granström A, Grelle A, Isacsson G, Kjellander P, Larsson S, Lindh M, Pettersson LB, Strengbom J, Stridh B, Sävström T, Thor G, Wikars L-O, Mikusiński G (2019). Rapid ecological response and intensified knowledge accumulation following a north European mega-fire. Scand J Forest Res 34: 234–253. https://doi.org/10.1080/02827581.2019.1603323.
Hille MG, den Ouden J (2005) Fuel load, humus consumption and humus moisture dynamics in Central European Scots pine stands. Int J Wildland Fire 14: 153–159. https://doi.org/10.1071/WF04026.
Hille MG, Stephens S (2005) Mixed conifer forest duff consumption during prescribed fires: tree crown impacts. Forest Sci 51: 417–424. https://doi.org/10.1093/forestscience/51.5.417.
Ivanova GA, Kukavskaya EA, Ivanov, VA, Conard, SG, McRae DJ (2020) Fuel characteristics, loads and consumption in Scots pine forests of central Siberia. J For Res 31: 2507–2524. https://doi.org/10.1007/s11676-019-01038-0.
Keane R (2015) Wildland fuel fundamentals and applications. Springer Cham. https://doi.org/10.1007/978-3-319-09015-3.
Larjavaara M (2005) Climate and forest fires in Finland – influence of lightning-caused ignitions and fuel moisture. Diss For 5. https://doi.org/10.14214/df.5.
Lawson BD, Armitage, OB, Dalrymple GM (1993) Ignition probabilities for simulated peoplecaused fires in British Columbia’s lodgepole pine and white spruce subalpine fir forests. Proceedings of 12th Conference On Fire & Forest Meteorology, Oct 26–29, 1993, Jekyll Island, Georgia, USA.
Lindberg H, Aakala T, Vanha-Majamaa I (2021) Moisture content variation of ground vegetation fuels in boreal mesic and sub-xeric mineral soil forests in Finland. Int J Wildland Fire 30: 283–293. https://doi.org/10.1071/WF20085.
Mäkipää R (2000) Pleurozium schreberi. In: Reinikainen A, Mäkipää R, Vanha- Majamaa I, Hotanen J-P (eds) Kasvit muuttuvassa metsäluonnossa. [Changes in the frequency and abundance of forest and mire plants in Finland since 1950]. Tammi, Jyväskylä.
Mäkipää R, Heikkinen J (2003) Large-scale changes in abundance of terricolous bryophytes and macrolichens in Finland. J Veg Sci 14: 497–508. https://doi.org/10.1111/j.1654-1103.2003.tb02176.x.
Marino E, Madrigal J, Guijarro M, Hernando C, Díez C, Fernández Filgueira C (2010) Flammability descriptors of fine dead fuels resulting from two mechanical treatments in shrubland: A comparative laboratory study. Int J Wildland Fire 19: 314–324. https://doi.org/10.1071/WF08123.
Masinda MM, Sun L, Wang G, Hu T (2020) Moisture content thresholds for ignition and rate of fire spread for various dead fuels in northeast forest ecosystems of China. J For Res 32: 1147– 1155. https://doi.org/10.1007/s11676-020-01162-2.
Munger, Gregory T (2008) Cladonia spp. In: Fire effects information system. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory. https://www.fs.usda.gov /database/feis//lichens/claspp/all.html.
Norum RA (1982) Predicting wildfire behavior in black spruce forests in Alaska. U.S. Department of Agriculture, Forest Fire, Pacific Northwest Forest and Range Experiment Station, Portland, OR, Research Note PNW-401. https://doi.org/10.2737/PNW-RN-401.
Nousiainen, H (2000) Cladina In: Reinikainen A, Mäkipää R, Vanha-Majamaa I, Hotanen JP (eds) Kasvit muuttuvassa metsäluonnossa. [Changes in the frequency and abundance of forest and mire plants in Finland since 1950]. Tammi, Jyväskylä.
Ohlson M, Brown KJ, Birks HJB, Grytnes JA, Hornberg G, Niklasson M, Seppä H, Bradshaw RHW (2011) Invasion of Norway spruce diversifies the fire regime in boreal European forests. J Ecol 99: 395–403. https://doi.org/10.1111/j.1365-2745.2010.01780.x.
Péch G (1989) A model to predict the moisture content of reindeer lichen. Forest Sci 35: 1014–1028. https://doi.org/10.1093/forestscience/35.4.1137.
Péch G (1991) Dew on reindeer lichen. Can J For Res 21: 1415–1418. https://doi.org/10.1139/x91-198.
Peterson WL, Mayo JM (1975) Moisture stress and its effect on photosynthesis in Dicranum polysetum. Canad J Bot 53: 2897–2900. https://doi.org/10.1139/b75-318.
Pya N (2018) scam: shape constrained additive models. R package version 1.2-3.
Ryan KC (2002) Dynamic interactions between forest structure and fire behavior in boreal ecosystems. Silva Fenn 36: 13–39. https://doi.org/10.14214/sf.548.
Schimmel J, Granström A (1997) Fuel succession and fire behavior in the Swedish boreal forest. Can J For Res 27: 1207–1216. https://doi.org/10.1139/x97-072.
Tanskanen H, Venäläinen A, Puttonen P, Granström A (2005) Impact of stand structure on surface fire ignition potential in Picea abies and Pinus sylvestris forests in southern Finland. Can J For Res 35: 410–420. https://doi.org/10.1139/X04-188.
Tanskanen H, Granström A, Venäläinen A, Puttonen P (2006) Moisture dynamics of moss dominated surface fuel in relation to the structure of Picea abies and Pinus sylvestris stands. For Ecol Manage 226: 189–198. https://doi.org/10.1016/j.foreco.2006.01.048.
Tesky J (1992) Pleurozium schreberi. In: Fire effects information system. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory. https://www.fs.usda.gov/database/feis/plants/bryophyte/plesch/all.html.
Tryterud E (2003) Forest fire history in Norway: from fire-disturbed pine forests to fire-free spruce forests. Ecography 26: 161–170. https://doi.org/10.1034/j.1600-0587.2003.02942.x.
Varner JM, Kane JM, Kreye JK, Engber E (2015) The flammability of forest and woodland litter: a synthesis. Curr Forestry Rep 1: 91–99. https://doi.org/10.1007/s40725-015-0012-x.
Total of 35 references.
