John Alexander Pulgarin Diaz  ,
Markus Melin,
Tiina Ylioja,
Päivi Lyytikäinen-Saarenmaa,
Heli Peltola,
Olli-Pekka Tikkanen
,
Markus Melin,
Tiina Ylioja,
Päivi Lyytikäinen-Saarenmaa,
Heli Peltola,
Olli-Pekka Tikkanen
Relationship between stand and landscape attributes and Ips typographus salvage loggings in Finland
Pulgarin Diaz J. A., Melin M., Ylioja T., Lyytikäinen-Saarenmaa P., Peltola H., Tikkanen O.-P. (2024). Relationship between stand and landscape attributes and Ips typographus salvage loggings in Finland. Silva Fennica vol. 58 no. 3 article id 23069. https://doi.org/10.14214/sf.23069
Highlights
- In Finland, the European spruce bark beetle (SBB) prefers mature stands (high age and mean diameter at breast hight), herb-rich heath forest sites and semi-coarse or coarse heath forest soil type, as well as a short distance to the closest wind damage from the previous-year, SBB damage from previous-year and particularly to clear-cuts
- These stand types should be prioritised for monitoring SBB damage.
Abstract
Disturbances caused by the European spruce bark beetle (SBB; Ips typographus L.) on Norway spruce (Picea abies (L.) H. Karst.), have increased immensely across Central and Northern Europe, and are expected to increase further as a result of climate change. While this trend has been noted in Finland, so far limited research has been published. To support proper SBB risk management in Finland, we compared stand properties between salvage loggings due to SBB damage during 2012–2020 (4691 cases) and spruce stands free of SBB damage. Also, we explored the role of landscape attributes as drivers of SBB damage. We considered the forest stand attributes of site fertility class, stand development class, soil type, stand mean diameter at breast height and mean stand age. Considered forest landscape attributes were the distance from SBB-damaged stands to the closest clear-cut, to previous-year SBB-damaged stands and to the previous-year wind-damaged stand. We used nationwide forest logging and forest stock data, and analysed forest stand attributes using chi-squared and Mann-Whitney U tests and landscape attributes using generalised linear mixed models. Based on our findings, the SBB didn’t damage stands randomly, but prevailed in mature stands (high age and high mean diameter at breast height), in herb-rich heath forest site types and in semi-coarse or coarse heath forest soil soils. We found correlation between the landscape variables and the number of salvage loggings, with a higher number of loggings due to SBB damage close to clear-cuts. Our results help to find risk areas of SBB damage.
Keywords
Picea abies;
wind damage;
clear-cuts;
European spruce bark beetle;
stand attributes
-
Pulgarin Diaz,
School of Forest Sciences, University of Eastern Finland, P.O. Box 111, FI-80101 Joensuu, Finland
 https://orcid.org/0000-0003-0554-8254
E-mail
alexander.pulgarin.diaz@uef.fi
https://orcid.org/0000-0003-0554-8254
E-mail
alexander.pulgarin.diaz@uef.fi
-
Melin,
Natural Resources Institute Finland (Luke), P.O. Box 68, FI-80101 Joensuu, Finland
https://orcid.org/0000-0001-7290-9203
E-mail
markus.melin@luke.fi
-
Ylioja,
Natural Resources Institute Finland (Luke), Latokartanonkaari 9, FI-0079 Helsinki, Finland
https://orcid.org/0000-0002-8840-7504
E-mail
tiina.ylioja@luke.fi
-
Lyytikäinen-Saarenmaa,
School of Forest Sciences, University of Eastern Finland, P.O. Box 111, FI-80101 Joensuu, Finland
https://orcid.org/0000-0003-1884-3084
E-mail
paivi.lyytikainen-saarenmaa@ef.fi
- Peltola, School of Forest Sciences, University of Eastern Finland, P.O. Box 111, FI-80101 Joensuu, Finland E-mail heli.peltola@uef.fi
-
Tikkanen,
School of Forest Sciences, University of Eastern Finland, P.O. Box 111, FI-80101 Joensuu, Finland
https://orcid.org/0000-0002-3875-2772
E-mail
olli-pekka.tikkanen@uef.fi
Received 3 November 2023 Accepted 19 March 2024 Published 17 April 2024
Views 45169
Available at https://doi.org/10.14214/sf.23069 | Download PDF

Supplementary Files
1 Introduction
Insect pests are becoming increasingly prominent due to range expansion as a result of climate change (Battisti and Larsson 2015). This increase also holds for the European spruce bark beetle, Ips typographus L. (SBB; Coleoptera: Curculionidae), which is the most damaging forest insect to Norway spruce (Picea abies (L.) H. Karst.; hereafter referred to as spruce) (Venäläinen et al. 2020; de Groot et al. 2023). The amount of timber damaged due to SBB has greatly increased in Europe over recent decades and the high risks areas are expected to increase under climate change (e.g. Hlásny et al. 2019; Venäläinen et al. 2020). In addition, spruce planted outside of its natural range, on sites with lower soil-water holding capacity and an increased growing stock have made spruce forests more prone to SBB outbreaks (e.g. Hlásny et al. 2019).
While the unprecedented damage caused by SBB to date has occurred mainly in Central and Eastern Europe, conditions favourable to outbreaks have become more prevalent also in Northern Europe due to increasing temperatures and occurrence of droughts (Jönsson et al. 2012; Battisti and Larsson 2015; Tikkanen and Lehtonen 2023). In Sweden, SBB damage is predicted to worsen under warmer conditions (Jönsson et al. 2012), as the probability of high population levels (Jönsson et al. 2007) and frequency of outbreak occurrence increases (Müller et al. 2022; Kärvemo et al. 2023). The future far-northern distribution limit of SBB outbreaks in Sweden is expected to border western Finnish Lapland (68.8°N) under both low emissions and high emissions climate scenarios for 2070 (Hof and Svahlin 2016). In Finland, the frequency of SBB outbreaks have been increasing since the 2010s (Blomqvist et al. 2018; Hantula et al. 2023; Tikkanen and Lehtonen 2023). Although the scale of damage in Finland remains minor compared to that of Central and Eastern Europe (Hantula et al. 2023), local concerns about increasing SBB disturbances and timber losses have been raised, especially in Southern and Central Finland. These growing concerns show the need to improve forest resilience through monitoring and predicting future SBB damage risks and further locally adapted risk management practices.
The population dynamics of economically impactful bark beetles (such as SBB in Eurasia and Dendroctonus ponderosae Hopkins [Coleoptera: Curculionidae] in North America) have been characterised as having 2–4 phases (Kunegel-Lion and Lewis 2020), according to the insect population size. The two most common are the endemic phase when the population density is low and the epidemic phase when the population density is high, with transition phases between. For SBB, epidemics densities are promoted by favourable weather and presence of susceptible trees in vast areas, often in the vicinity of previous-years damaged areas (Wermelinger 2004; Hlásny et al. 2021). Epidemics are also favoured by drought at large scales, which makes trees less ready to defend against insect attacks (Müller et al. 2022). During epidemics, beetles search for new hosts over short distances – within 0–500 m (Kautz et al. 2011; Kärvemo et al. 2014), similar to what happens in large-scale windthrow events (Hroššo et al. 2020). Once population levels are high enough, they can keep elevated levels by themselves, given favourable weather and presence of host trees. At this moment, all trees regardless of their size and vitality are suitable for attack (Hroššo et al. 2020; Hlásny et al. 2021).
High population density of natural enemies, low availability of breeding material and unfavourable weather can cause the SBB population to return to the endemic phase (Wermelinger 2004; Hlásny et al. 2021). During the endemic phase, SBB mainly attack weakened trees, windthrows or older trees with low vigour (Louis et al. 2015), causing low stand mortality. As a result, beetles must disperse over long distances (>1000 m) to find suitable hosts (Netherer et al. 2019; Hlásny et al. 2021).
The minimum effective thermal sum for development from egg to pupae is approximately 334.2 degree days above a lower threshold of 8.3 °C (Wermelinger and Seifert 1998). These requirements already exist within Finland under current climate. Climate warming is likely to increase the probability of two annual generations or at least the occurrence of sister broods (Jönsson et al. 2007; Marini et al. 2017; Bentz et al. 2019). In addition, local conditions may become favourable due to clear-cut edges that create warmer microclimatic in comparison to the forest interior, further enhancing SBB breeding (Lindman et al. 2023). Given that clearcut is a common practice in Finnish forestry, we need to study its role as a factor promoting SBB damage.
Sanitary and salvage logging are the most common forest management approaches to control SBB populations and prevent further damage (e.g. Hlásny et al. 2019, 2021 and references within). Only a small fraction of the research considering these techniques covers the boreal forest, especially within Finland, where research on the stand and landscape attributes contributing to SBB damage has been limited to date.
To support the development of proper risk analysis and forest management, we aimed to explore the properties of spruce stands where salvage loggings due to SBB damage took place and of spruce stands free from SBB damage. This setting allowed us to explore, for the first time, forest-use declaration data and contrast it with the forest stock data, to determine if certain characteristics predispose spruce stands to SBB attack. We then explored the role of landscape attributes in increasing the occurrence of SBB damage. All these settings will help us to better understand the characteristics that predispose a spruce stand to SBB damage and communicate our results in a practical way. We were also interested in determining the advantages and disadvantages these datasets have for such analysis. We expected the properties of damaged and non-damaged spruce stands to differ, and that distance from the closest clear-cut, previous-year SBB damage and previous-year wind damage affects the number of new SBB-damaged stands at national-level scale. Our study was based on nationwide open forest data and forest-use declarations between 2012–2020.
2 Methods
2.1 Study area
Finnish forestry land spans 26.6 M ha, of which 77% is productive forest. Of the productive forest, 64% is dominated by Pinus sylvestris L. and 25% by spruce (Korhonen et al. 2021). In South Finland, productive forest covers 73% of the land (11.2 M ha), with 33% dominated by spruce. The study area (Fig. 1) covered the southern half of Finland between 60°N–64°N, which is predominately in the southern boreal vegetation zone, accompanied by marginal hemiboreal forests in the southwest and middle boreal forests in the upland watersheds of Central Finland (Korhonen et al. 2021). During the reference period of 1991–2020 (current climate), the mean annual temperature ranged between 6 °C in southern Finland (Helsinki, latitude 60.2°N) and 2.6 °C in the north (Kajaani, latitude 64.2°N), while the annual total precipitation ranged between 680 mm in the south and 585 mm in the north (Finnish Meteorological Institute 2022). Southern Finland has mostly flat lowlands, increasing up to 200 m a.s.l. in a north-eastwards direction (Nygren 2011). The forests in the study area are coniferous-dominated and are primarily managed forests with an even-aged structure (Korhonen et al. 2021).
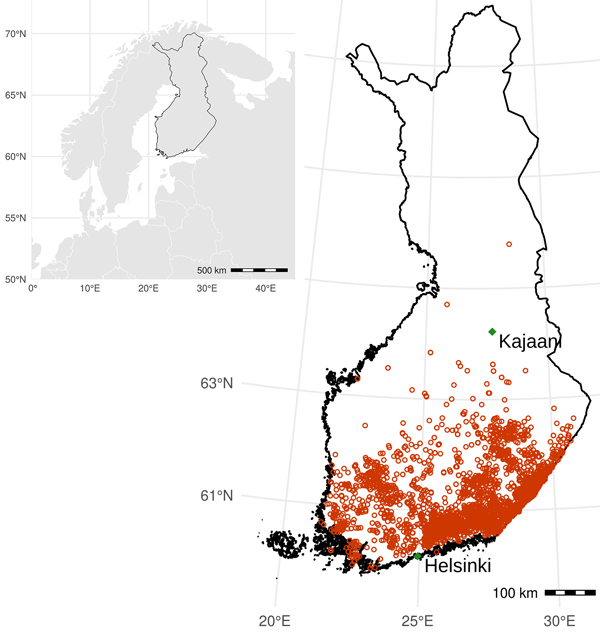
Fig. 1. Study area: Southern half of Finland from 60°N–64°N, and distribution of salvage logging operations due to Ips typographus damage (red circles) during 2012–2020, according to forest-use declaration data.
2.2 Forest datasets
For assessing the relationships between forest structure and SBB damage, we used two nationwide geo-datasets (GeoPackage file format) provided by the Finnish Forest Centre (Fig. 2): First, the forest stock dataset contained information about the structure and attributes of privately owned forest stands (Metsäkeskus 2021b). This supplied information on spruce stands that were free of SBB damage (non-damaged stands, hereafter referred to as NDS) and helped to estimate the location of clear-cuts. And second, the forest-use declaration dataset contained information about salvage logging operations resulting from SBB damage (SBBD) or wind damage (WD) (Metsäkeskus 2021a). These datasets use the forest stand as the smallest management unit, meaning that the area inside the stand polygon is uniform in terms of its structure and management history.
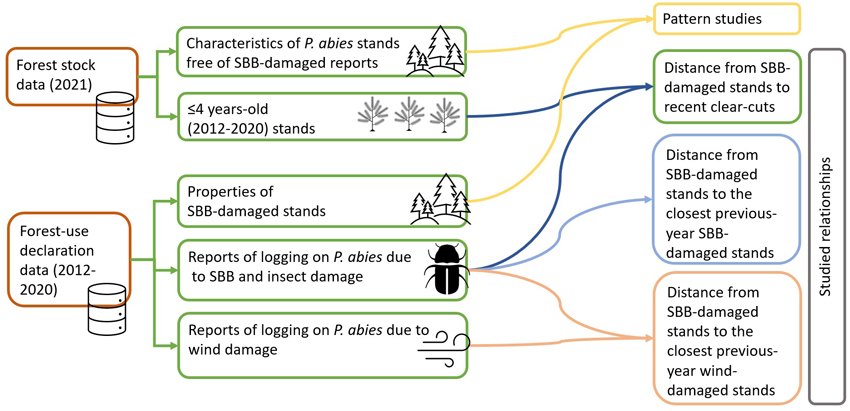
Fig. 2. Methodological workflow for studying the relationship between forest stand and landscape attributes and Ips typographus (SBB) damage on Picea abies stands in Finland (stands ≤4 years old were used as a proxy for recent clear-cuts).
The forest stock spatial dataset (vector, polygon) included information from all privately owned Finnish forests collected through both remote sensing and fieldwork (Metsäkeskus 2021a). We downloaded the data on 18 November 2021, chose ‘data originated during inventory’, stands (tree-stratum in the dataset) dominated by spruce, but excluded drained peatlands and northern Finnish regions (Lapland and Kainuu, >64°N) because SBB damage hasn’t been reported in these areas. The final dataset included an area of 3 629 695 ha distributed across 3 109 706 stands. As a proxy for recent clear-cuts assigned to each analysed year, we selected stands less than or equal to four years old using the date of the last updated age. This setting helped us evaluate the effect that clear-cuts have on new SBB damage up to four years after clearcutting. The analysis was done using the package Simple Features ‘sf’ version 1.0-5 (Pebesma 2018) in RStudio 022.07.0+548.
The forest-use declaration spatial dataset (vector, polygon) reported logging operations intentions conducted in privately owned Finnish forests (Metsäkeskus 2021b). We downloaded the geo-dataset on 18 October 2021 and selected stands having spruce as the main tree species and SBB damage or ‘insect damage’ as the reason for the salvage logging during 2012–2020. We included ‘insect damage’ as there are only a few cases in which insects other than SBB, such as the bark beetle Pityogenes spp., could cause damage to spruce stands that would require salvage logging. Also, in the earlier years of forest-use data collection, reports SBB damage was sometimes registered as ‘insect damage’. This dataset described polygons (stands) that were harvested due to SBB damage, but it didn’t contain other information about e.g. the extent of damage (dead wood versus infested wood) or the SBB population size. We also selected stands having spruce as the main tree species and where salvage logging had taken place due to wind damage (WD).
Because the year when the damage occurred wasn’t explicit in the forest-use declaration dataset, we assigned the most likely year of damage occurrence, following the lead by Netherer et al. (2019) and Müller et al. (2022). For this, we assigned the year of the report (declaration) as the year of damage if the report occurred from June–December. If the report month was from January–May, we assigned the previous year as the year of damage, because in January–May SBB is mainly hibernating in Finland (Annila 1969) meaning damage reported from January–May must therefore originated in the previous year. Similarly, we assigned the year of wind damage to correspond to the swarming of SBB. For this, we assigned the wind damage salvage loggings reported from September-December to the next year, as in Finland the SBB swarms mainly from May–August, meaning that blown down trees after August are available for breeding in the next year.
2.3 Data analysis
To test if the occurrence of logging operations due to SBB damage is influenced by the occurrence of previous-year SBB damage, we measured autocorrelation. For this, we used the time series of logging operations due to SBB damage and the correlogram (Cowpertwait and Metcalfe 2009) produced by The R Stats Package v. 4.2.2 (R Core Team 2023).
Further, to study the relationship between stand attributes and the occurrence of SBB damage (or salvage loggings), we compared spruce dominated stands with and without SBB damage in terms of five stand attributes – site fertility class, stand development class, soil type, stand mean diameter at breast height (Dmean) and stand mean age (Table 1). We used Chi-squared tests (α = 0.05) to compare the distribution of the two groups of stands (SBB and NDS) in terms of site fertility class, stand development class and soil type. We used the Mann-Whitney U non-parametric tests (α = 0.05) to compare Dmean and stand age between spruce dominated stands with and without SBB damage. For this last comparison, we selected stands with a Dmean >15 cm and age >25 years old, which threshold values are fairly close to the developed stand class (Table 1), as SBB didn’t attack younger stands (see 3.2 section). We ran these comparisons in the Statsr package v.0.3.0 in RStudio 2022.07.0+548 (Rundel et al. 2021).
| Table 1. Stand and landscape attributes studied with relation to Ips typographus (SBB) damage in Finland. The presented variables were in both datasets.* | ||
| Category | Attributes | Description |
| Stand | Site fertility class | In decreasing order of fertility: • Herb-rich forest (most fertile, high water holding capacity) • Herb-rich heath forest • Mesic heath forest • Sub-xeric heath forest • Xeric heath forest (barren, low fertility, low water holding capacity) |
| Stand development class | • Young stand: height <7 m, overstory of seed or shade trees • Developing stand: height >7 m, age >11 years, Dmean = 8–16 cm • Developed stand: age >25 years, Dmean >16 cm. • Mature stand: mature enough for regenerative cutting • Shelter-tree stand: natural spruce regeneration protected by a cover of trees | |
| Soil type | • Coarse deposited soil • Coarse moraine soil • Fine-grained deposited soil • Fine-grained heath forest soil • Moraine soil and fine-grained particles • Mull soil • Outcrop stone field • Peat soil • Sedge peat • Semi-coarse or coarse heath forest soil • Sphagnum peat sensitive to erosion • Stony coarse moraine • Stony fine-grained heath forest soil • Stony semi-coarse or coarse heath forest soil | |
| Mean diameter at breast-height | Dmean for each analysed stand (cm) | |
| Mean age | Mean age for each analysed stand (years) | |
| Landscape | Distance to the closest recent clear-cut | Recent clear-cut stands (or stands 0–4 years old) were selected from the forest stock dataset, using the date of the last updated age for the analysed years (2012–2020) |
| Distance to the closest previous-year SBB-damaged stand (SBBDy–1) | Selected from the forest-use declaration dataset in relation to the year of occurrence of the current SBBD. | |
| Distance to the closest previous-year wind-damaged stand (WDy–1) | Selected from the forest-use declaration dataset in relation to the year of occurrence of the current SBBD. | |
| * A complete list of the soil types, site fertility and stand development classes present in Finland is available in Äijälä et al. (2019). Dmean = stand mean diameter at breast height. SBBD = SBB-damaged stand. | ||
Site fertility class in Finland (Table 1) refer to a site’s potential for wood production, which is affected by the interaction of all site factors, such as climate, soil vegetation and topography (Nygren 2011). Also, in Finnish mineral soils, site fertility is the combination of nutrient availability and water holding capacity which correlate strongly. In the Finnish soil fertility classification (Table 1), lower site fertility means lower water-holding capacity and vice versa, i.e. sites with poor water-holding capacity also have fewer available nutrients. The stand development class (Table 1) describes the silvicultural and wood production development stage of a stand at a certain time; it is determined based on the age, structure and previous forest treatments (Äijälä et al. 2019). For details on the site fertility and soil type classifications, see Korhonen et al. (2021).
We assessed three landscape attributes to determine their impact on the number of SBB damage (or salvage loggings): Euclidean distance from the centroid of the SBB-damaged stand to the closest clear-cut, to the closest previous-year SBB-damaged stand (SBBDy–1) and to the closest previous-year wind-damaged stand (WDy–1, Table 1). The Euclidean distances to these objects were determined by forming Euclidean distance rasters from them and by extracting the value of the raster to the centroid of the SBB-damaged stands. The rasters had a cell size of 100 m, meaning that the distances followed a 100 m classification (Fig. 3). The used cell sizes and ranges also agree with the distance resolution used in other SBB studies (Kärvemo et al. 2014; Müller et al. 2022) and the distances flown by the SBB (Hinze and John 2019).
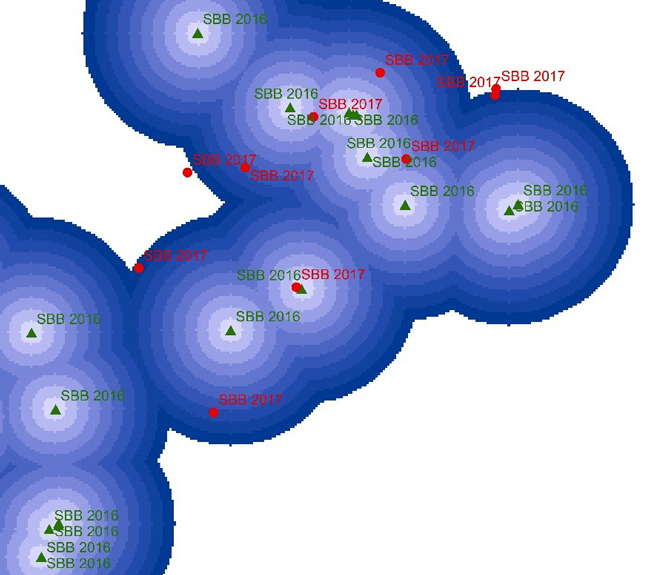
Fig. 3. Illustration on the measurement of the distance from the centroid of each Ips typographus-damaged stand (SBBD; in red) to the closest previous-year SBBD (SBBDy–1; in green). The distance buffers (concentric circles) in 0–1500 m range and 100 m cell size are drawn around the centroid point of SBBD2016 (in green) and centroid points of SBBD2017 (in red) stands. The distance from SBBD2017 to SBBD2016 was measured based on the cell where SBBD2017 occurred. The measurement of distance for the other two landscape attributes (clear-cuts and previous-year wind-damaged stands [WDy–1]) followed the same procedure.
To quantify the potential effect of the landscape attributes (independent variables) on the number of stands damaged by SBB (dependent variable), we used generalised linear mixed models (GLMM). The fixed-effect variables were the distance class (in 100 m intervals, from 0to 1500 m) for each measured landscape attribute (clear-cut, SBBDy–1 & WDy–1) and their interaction, while the year of damage (2012–2020) was set as the random-effect variable. We used a negative binomial probability distribution model for count data (α = 0.05), log link function, E(yi) = μi and Var(yi) = E(yi) × (1 + E(yi) / θ), where θ is the dispersion parameter (Hardin and Hilbe 2007; Brooks et al. 2017; Mehtätalo and Lappi 2020). During the model development, we noticed that, depending on the landscape attribute, the dependent variable became more dispersed compared to the mean. We controlled this variability by modelling the dispersion parameter dependent on the landscape attributes, setting log(θ) = b0 + b1 × (landscape attribute). For modelling, we used the glmmTMB package (Brooks et al. 2017). We checked for overdispersion by plotting Pearson residuals against the predicted values (Mehtätalo and Lappi 2020) and in the dispersion statistic, Pearson X2 divided by ‘residuals degrees of freedom’ (Hardin and Hilbe 2007; Hilbe 2009; Mehtätalo and Lappi 2020).
3 Results
3.1 Wind and spruce bark beetle damage
The total number of salvage logging operations during the study period (2012–2020) due to WD was 20 486 and 4653 due to SBB damage. The annual number of logging events (Fig. 4) showed one clear peak for each damaging agent – in 2013 for SBB and in 2014 for WD – followed by a decreasing trend. After 2014, the annual number of WD salvage loggings was higher than that of SBB salve loggings.
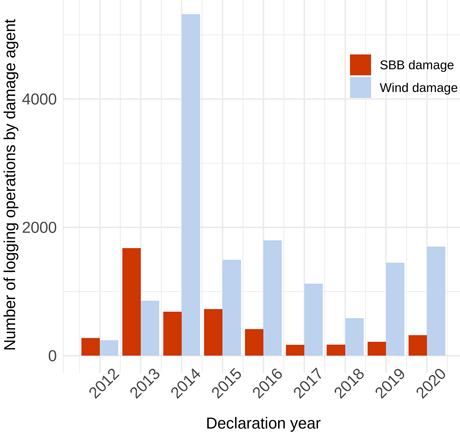
Fig. 4. Number of reported logging operations on Picea abies stands due to Ips typographus (SBB) and wind damage, across Finland during 2012–2020.
Much of the SBB damage concentrated in Southeastern Finland, close to the Russian border (Fig. 1). The two northernmost SBBDs were in the Northern Ostrobothnia region, but most were located between 60–62°N (Southern Finland). Autocorrelation in time for the number of SBB salvage loggings was absent (p > 0.05; Fig. 5).
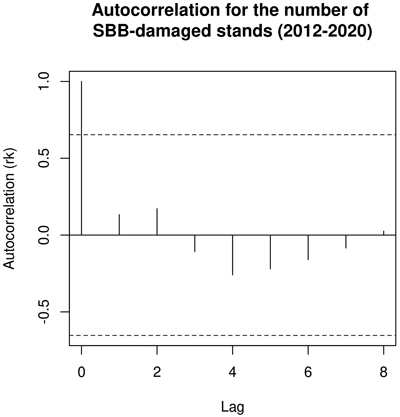
Fig. 5. Correlogram for the number of logging operations due to Ips typographus damage in Finland during 2012–2020.
3.2 Forest-stand attributes of SBB-damaged stands
When considering forest-stand attributes, we found that SBB didn’t damage stands randomly. That is, the structure of the SBB damaged stands differed from what was available in the overall landscape (NDS). If SBB would randomly select stands from all available spruce stands we would expect a similar distribution between SBBD and NDS, but this wasn’t the case for all the studied attributes.
Approximately 95% of SBBD occurred in either mesic heath forests (63%) or herb-rich heath forests (32.4%, Fig. 6A; see Supplementary file S1 for details). In NDS these proportions were 63.6% and 28.2%, respectively. We found that herb-rich heath forest was slightly overrepresented in SBBD when compared to NDS, and the sub-xeric heath forest class was underrepresented. The sub-xeric heath forest class was the greatest contributor to the statistically significant difference between site fertility classes of NDS and SBB-damaged stands.
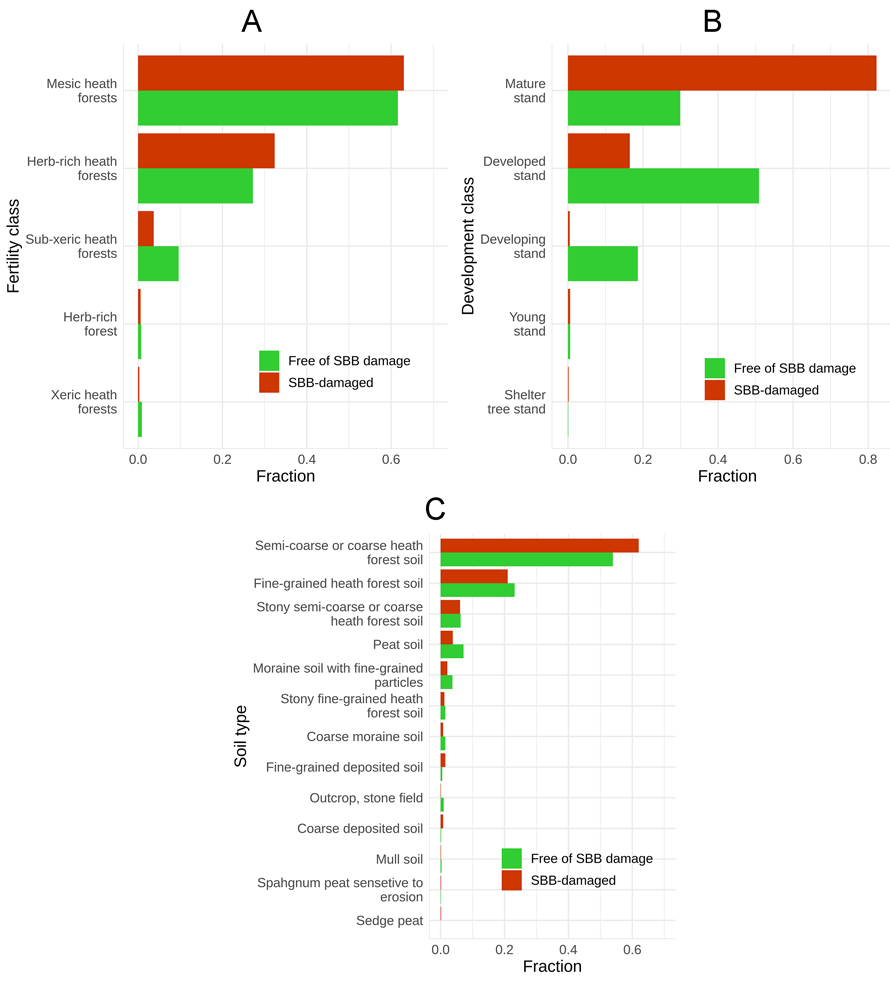
Fig. 6. Distribution of Picea abies stands damaged by Ips typographus (SBB) and free of SBB in Finland during 2012–2020, by (A) site fertility class, (B) stand development class and (C) soil type. See Supplementary file S1 for details.
Concerning the stand development class of the stands, most SBBD (98.7%) occurred in mature stands (82.2%) and developed stands (16.5%; Fig. 6B; see Suppl. file S1 for details). While in NDS these proportions were 29.9% for mature stands and 50.9% for developed stands. The SBBD were overrepresented in the mature stand class and underrepresented in the developed stand class compared to NDS (Fig. 6). Mature stand and developed stand classes contributed the most to the statistically significant difference between SBBD and NDS, being mature stand as the main contributor (Table 2).
| Table 2. Statistical results comparing Picea abies stands damaged by Ips typographus (SBB) and stands free of SBB-damage in Finland during 2012–2020, based on Chi-squared (X2; α = 0.05) and Mann-Whitney U tests (U; α = 0.05). | |||||
| Attributes | Statistic value | df * | Number of stands | p-value | |
| SBB-damaged | Non SBB-damaged | ||||
| Site fertility class | X2 = 109.6 | 4 | 4650 | 2 994 536 | <0.001 |
| Stand development class | X2 = 5661.9 | 4 | 4265 | 2 441 024 | <0.001 |
| Soil type | X2 = 562.7 | 13 | 4518 | 3 106 289 | <0.001 |
| Mean diameter at breast height >15 cm | U = 5 137 488 938 | 3116 | 2 165 405 | <0.001 | |
| Mean age >25 years | U = 5 128 003 | 3202 | 2 431 519 | <0.001 | |
| * df = degrees of freedom. | |||||
For the soil type, most SBBD stands were in the soil class semi-coarse or coarse heath forest soil (62%) and in fine-grained heath forest soil (21%, Fig. 6C; see Suppl. S1 for details). In NDS these proportions were 53.9% in semi-coarse or coarse heath forest soil and 23.2% were in fine-grained heath forest soil. Fascinatingly, SBBD was overrepresented in semi-coarse or coarse heath forest soil, while the sedge peat class contributed the most to the statistically significant difference between the two types of stands. However, the soil type classification in the used data has significant limitations, which are discussed later.
When considering forest age, we found that the age distribution of SBBD and NDS differed (p < 0.001; Table 2), SBB damage occurred in general in older stands (Fig. 7A). The age distribution for SBB-damaged stands peaked at 80 years, the same occurred roughly 20 years earlier for NDS. Nearly all (97.3%) of the SBB damage occurred in forests aged between 44–119 years, whereas this age class comprised ca. 30% of the non-damaged stands.
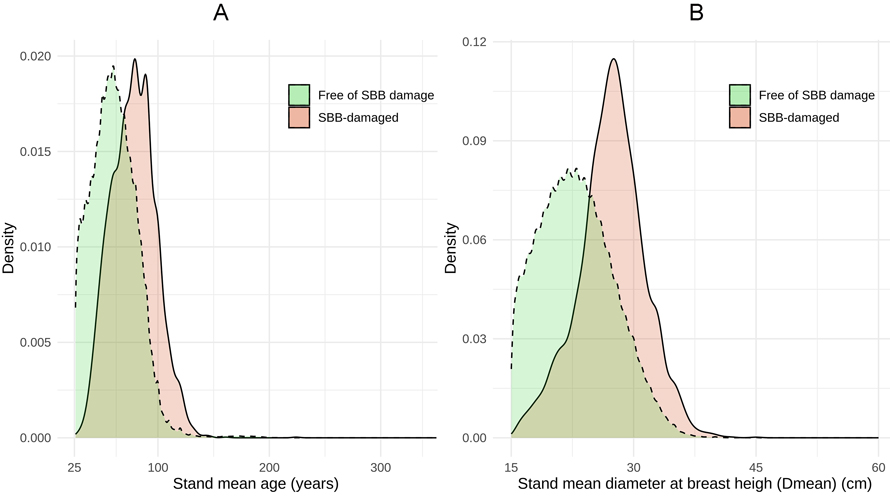
Fig. 7. Density distribution of Picea abies stands damaged by Ips typographus (SBB) and free of SBBD damage in Finland during 2012–2020, in terms of (A) mean age (>25 years) and (B) mean diameter at breast height (Dmean >15 cm) of the stand.
The Dmean distribution in SBBD and NDS differed as well (p < 0.001; Table 2, Fig. 7B). The SBB damage was most common in stands with Dmean between 20–35 cm, showing a peak at 27.5 cm.
3.3 Effect of landscape attributes
We found a statistically significant effect of the distance from SBB-damaged stands to clear-cuts, WDy–1 and SBBDy–1 on the annual number of SBBD (Table 3). This means that SBB damage was more common near clear-cuts and similar effects – though not as strong – were also seen near previous year’s wind and SBB damage. Our model shows a low discrepancy between observations and the model results, being homogeneous for the used variables. Also, the landscape attributes were good predictors of the dispersion model (p = 0.046 for SBBy–1 and p = 0.016 for WDy–1; Table 4). The dispersion model shows that there is higher dispersion concerning the mean of SBB damage generated by WDy–1 than for SBBy–1, and these two variables have higher dispersion than the generated by clear-cuts (Table 4). It is also possible to see that variability in Fig. 8, where the 95% Wald-type confidence intervals for the predicted average count are much narrower for WDy–1 and SBBy–1 than for clear-cuts, concerning their mean.
| Table 3. Analysis of deviance (Type III Wald Chi-square tests) of the effects of landscape attributes on the number of Ips typographus-damaged stands (SBBD) in Finland. See methods for model details. | |||
| Variable | Chisq | Df | Pr(>Chisq) |
| Intercept | 57.423 | 1 | <0.001 |
| Distance class | 301.784 | 14 | <0.001 |
| Landscape attribute | 59.566 | 2 | <0.001 |
| Landscape attribute × Distance class | 68.783 | 28 | <0.001 |
| Table 4. Dispersion model for the generalised linear mixed model with a negative binomial probability distribution for the response variable. See methods for model details. | ||||
| Predictor* | Estimate | Std. Error | z-value | Pr(>|z|) |
| Intercept | 6.156 | 2.545 | 2.418 | 0.016 |
| Landscape attribute SBBy–1 | –5.131 | 2.570 | –1.996 | 0.046 |
| Landscape attribute WDy–1 | –6.193 | 2.560 | –2.419 | 0.016 |
| * The landscape attribute clear-cut was the reference category. It is redundant, as it is a linear combination of the remaining parameters. | ||||
Our model shows also that the number of SBBD was the highest at a distance around 450 and 600 m from the closest clear-cut (Fig. 8). The reader should note that the biggest contributions of WDy–1 and SBBDy–1 were at longer distances and that the interaction between landscape attributes and distance class was statistically significant (p < 0.001, Table 3).
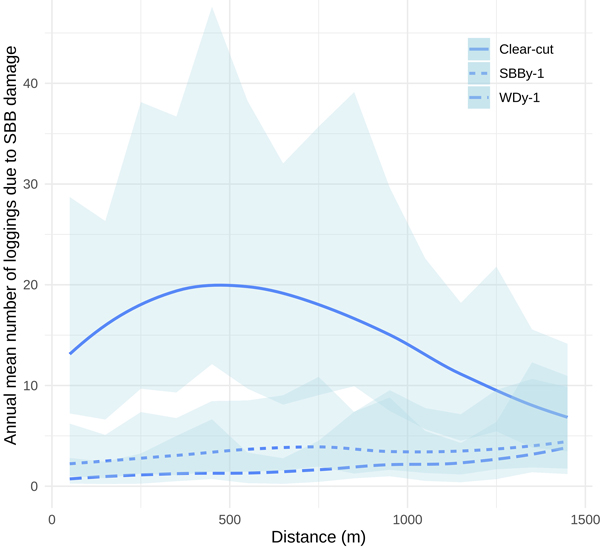
Fig. 8. Effect of distance from clear-cuts, previous-year Ips typographus damage (SBBy–1) and previous-year wind damage (WDy–1) on the mean number of SBB-damaged stands based on the generalised linear mixed model developed in this study. The shaded area represents the 95% Wald-type confidence intervals for the predicted average count.
4 Discussion
Analysing the available data across Finland revealed spatial patterns in SBB damage. We found that SBBD was prevalent in mature stands, sites with above average fertility (herb-rich heath forest) and soils with medium or low water holding capacity (semi-coarse or coarse heath forest), as well as in stands with higher Dmean and of older age. In addition, the distance to the studied landscape attributes correlated with the annual number of SBB salvage logging operations. These results allow us to provide practical suggestions for SBB-damage management measures in Finnish forests (see section 4.3).
4.1 Forest stand attributes of SBBD
Our analysis of site fertility class, stand development class and soil type showed that spruce stands, SBB attacked and NDS, were mainly in two classes of each of these variables, probably due to forest management. The most common classes for fertility were mesic heath forest and herb-rich heat forest, for development were mature stand and developed stand; and for soil type were semi-coarse or coarse heath forest soil and fain-grained heath forest soil.
In our study, SBB damage occurred predominantly in the site fertility classes that were most common in the landscape, mesic heath forests and herb-rich heath forests. The SBB damage was underrepresented in sub-xeric and xeric heath forest (dry and nutrient-poor) and overrepresented in semi-coarse or coarse heath forest soil. Soils such as fine-grained heath forest soil, moraine soil with fine-grained particles and peat soils, which have good water-holding capacity, presented fewer SBB. As a comparison, in Austrian Federal Forests, SBB damage was less prevalent in soils with limited water-holding capacity which leads to chronic stress in spruce stands (Netherer et al. 2019). This was also observed in Norway and Germany (Christiansen and Bakke 1988). It is important to note that soils having limited water holding capacity could favour SBB attacks as a result of poor defensive capacity (Netherer et al. 2015), but when there is an epidemic phase, also trees with good defensive traits are attacked (Hlásny et al. 2021). Our analysis of the effect of soil type, however, is limited as the soil types aren’t rigorously estimated during inventories. Thus, no exact data about the detailed sub-soil types exists even though the stands are classified based on it.
Concerning the stand development class, the overrepresentation of mature stands in SBBD indicates that older stands attract more SBB, as indicated in previous research (Eriksson et al. 2007). Also, we found that stands with higher mean Dmean (>19.9 cm) and mean age (>44.8 years) were more damaged by SBB – a finding also supported by previous research (Göthlin et al. 2000; Hroššo et al. 2020; Müller et al. 2022). The main reason for this is that trees with higher diameter at breast height (DBH) and age, have larger phloem and thicker bark, providing SBB larvae with better conditions for development (Wermelinger 2004; Kärvemo et al. 2014).
SBB usually attack trees with DBH of 20–25 cm, but during epidemics, they may attack smaller trees as well (Hlásny et al. 2021). This is illustrated also in the High Tatra Mountains (Slovakia-Poland border) where damage started at DBH ≈14.3 cm in the incipient epidemic phase (Mezei et al. 2014). If SBB population levels are high (epidemic phase), high and low-DBH trees are attacked, but during the low population phase, trees having a lower capacity to defend against attacks are preferred, as described in the introduction (Mezei et al. 2014; Hlásny et al. 2021).
4.2 Landscape attributes affecting new SBB damage
From previous research, it is known that the distance to clear-cuts, SBBy–1 and WDy–1 can increase the number of new SBB damage, as they create ideal conditions for the SBB population development (Kärvemo et al. 2014; Müller et al. 2022). However, in Finland the removal of wind-felled and SBB-infested spruces is required by law if their amount exceeds 10 m3 per hectare (Finlex 2013). Thus, the joint effect of the cases when the wind-felled and SBB-attacked trees were removed before the SBB adult emergence could be the cause for WDy–1 and SBBDy–1 injecting high variation into the model. Finally, damage intensity (i.e. the volume of damaged timber and/or affected area) may also correlate with these variables as previously shown (Kautz et al. 2011; Hlásny et al. 2021), but this data isn’t available in the Finnish salvage logging data.
In our study, the main landscape-related driver of SBB damage at the country-level was clear-cuts, which included not only spruce, but all species, leading to a stunning result. The effect of clear-cuts on number of salvage loggings (SBBD) was greater than the combined effect of WDy–1 and SBB y–1. In Sweden, it has been suggested that if the number of clear-cuts increases, new SBB damage will appear in the nearby area, around 200–500 m depending on the study (Kärvemo et al. 2014; Müller et al. 2022). Usually, new clear-cut edges create ideal conditions for SBB damage (Peltonen 1999; Lindman et al. 2023), as in such edges microclimate is more favourable for reproduction of SBB, spruce trees in these edges are also more susceptible to wind damage, and receive more solar radiation which could increase their bark temperature making them prone to drought stress.
Clear-cuts could have influenced in new SBB damage up to 1500 m, probably facilitated by Finland’s flat topography, which create massive differences in microclimates in more mountainous regions (Netherer et al. 2019; Hlásny et al. 2021). Wind-felled and weakened trees, common in clear-cut edges, are a resource that sustains the SBB population, providing breeding material to facilitate reproduction, population growth and development (Louis et al. 2015; Hroššo et al. 2020).
Earlier studies have shown high numbers of SBBD surrounding previous years’ WD areas (Eriksson et al. 2007; Økland et al. 2016); also two years old WD has been reported as an important driver, although most attacks occur during the first year after WD (Hroššo et al. 2020). We expected to find similar patterns in our nationwide approach. Apparently, more than 20 fallen spruce trees (approximately 10 m3 if the trees were mature) are needed to initiate substantial SBB damage in our study area (Eriksson et al. 2007). If any WD of this scale happened, the trees were assumed to be removed in less than one year – as required by Finnish forest legislation (Finlex 2013), before the beetles use it for reproduction. Then, longer feedbacks (more than one year) are unlikely. This missing link between SBB damage and proximity to WDy–1 suggests that other factors were driving SBB damage and can’t be completely analysed with the data that we used.
We expected to see a strong influence of SBBy–1 on new SBB damage, but according to our model, the influence of this attribute is low. This was also confirmed by the absence of temporal autocorrelation, meaning that, for all Finland, the number of SBB salvage loggings in one year didn’t affect their number in the following years. We expected to see similar results as previous research in Sweden, which indicated that SBB from previous years in less than 125 m increases the probability of new damage, with a weak signal up to 750 m (Müller et al. 2022). Similarly, other research has indicated a maximum distance of influence of 213 m (Stereńczak et al. 2019) and 500 m (Kautz et al. 2011). The difference between our results and the mentioned research may again be due to the Finnish forest management regulations that require damaged trees to be removed in less than one year (Finlex 2013). As a result, the influence of SBB damage in subsequent years may be very low.
It is important to note that our data comes only from managed forests and our results can’t be generalised to protected areas. Though not considered in this study, what occurs in protected Finnish forests could also affect the managed forests. Some WD and SBBD in nearby protected forests, where damaged timber isn’t removed, could also predispose the commercial forest we studied to SBB damage, but data on this isn’t available.
4.3 Limitations of the used data
The used datasets supplied a huge quantity of data to study the relationship between forest stands, landscape attributes and SBBD at a national level, not commonly found in SBB studies. Although the datasets were large enough to ensure clear pattern identification, as proven in our results, the data has limitations that prevent a more detailed analysis of the drivers of SBB damage.
The dataset showing the salvage loggings due to SBBD and WD, may have time gaps as the data are notices of intent prior to logging. Also, some of those logging notifications may not have been executed. Either case, the dataset showed the existence of a damage observation prior to the notice. In this, also old unnoticed damage could also be treated as a recent damage. In all these cases, the minimum levels of damage that foresters consider for notifying may be higher than required by law (>10 m3 ha–1). These factors may cause biases in our data, but we can’t clear their effects.
Nevertheless, as the management and compilation of this kind of datasets, such as the forest-use declaration dataset, are overly complex, there is still room for improvement, such as auto-checking the reported information when some vague information is registered. Increasing the types of valuable information supplied by forest owners could allow researchers to develop better risk management solutions.
4.4 Forest health management considerations
To decrease the risk of new SBB damage, the removal of single infested trees is often suggested in Central Europe (Hlásny et al. 2021). To be effective, this sanitation or salvage logging should be done before new SBB adults emerge from colonized spruce trees. However, in Finland these loggings often mean harvesting the entire stand even though only a portion of it has been attacked, creating new clear-cuts that and favouring new SBB damage in the next year. Therefore, information about clear-cuts in neighbouring forests may help to mitigate the risk of new SBB damage.
Our results show that Finnish spruce stands are predisposed to SBB damage, especially in mature stands (high age and Dmean). This suggests that as spruce stands grow, they become closer to the preferred development stage (having high Dmean and age) for SBB attack. Then shortening the rotation period may reduce this predisposition. Also, clear-cuts should be better planned to minimise the exposure of spruce stands with vulnerable conditions to clear-cut-edge effects. Another alternative measure is to implement continuous cover forestry, which may help to avoid the edge effect and decrease a forest’s susceptibility to SBB damage. Yet, not enough experimental research exists on the SBB damage risks at such management.
Promotion of mixed tree species forests is one strategy to reduce SBB damage risk (Huuskonen et al. 2021; de Groot et al. 2023). Pure spruce stands should be particularly avoided in herb-rich heath forest and semi-coarse or coarse heath forest soil, as stands growing in these conditions are more susceptible to SBB damage, based on our and others’ findings (Huuskonen et al. 2021; Müller et al. 2022; de Groot et al. 2023). It is also important to consider that severe storms and droughts create ideal conditions for SBB epidemics and its control by infested-tree removal may become very difficult at the landscape level (Kärvemo et al. 2014), even more difficult if new clear-cuts are created. Therefore, future risk mitigation should consider more diverse forest landscapes.
Implementing all these significant changes to Finnish silviculture and coordinating the related forest management activities will likely be a very difficult task, as there are multiple forest owners with differing objectives (Triviño et al. 2023). Therefore, forest governance has a key role in maintaining good forest health. In addition, state forest administration should emphasise on the quality of forest health information. Currently, the reporting of damage is a forest owner’s responsibility, who may not have enough resources for forest health monitoring, leading to underreporting and incorrect damage agent identification.
5 Conclusions
The enormous nationwide Metsäkeskus (Forest Centre) datasets used in this research helped us to find the forest stand and landscape attributes that are critical for SBB damage occurrence, although these datasets could be improved to study SBB dynamics in boreal forest landscapes. Mature stands, herb-rich heath forest and semi-coarse or coarse heath forest soil, as well as a distance to the closest clear-cuts – no matter the species, wind damage from the previous year and SBB damage from the previous year are critical for new SBBD.
We consider it necessary to find measures to increase forest resilience against SBB damage in Finland, starting with the identification of susceptible stands and avoidance of cultivation of pure spruce stands on herb-rich heath forest and semi-coarse or coarse heath forest soil. We need to pay more attention to forest health management to ensure sustainable ecosystem services and to avoid large-scale damage (Hof and Svahlin 2016; Venäläinen et al. 2020).
Our findings contribute to the identification of SBB stand preferences in Finland and to prioritise places for monitoring and management activities. Also, we see a probable interaction between environmental, stand and population-level factors, which may make the prediction of new SBB attacks really difficult. However, improved data on forest damage could help to overcome these obstacles. Although the management of enormous nationwide datasets is not an easy task, we see that the data we used have great potential for SBB damage risk analysis in Finland.
Declaration of openness of research materials, data, and code
The used datasets are distributed by Metsäkeskus (www.metsakeskus.fi). The statistical code is available upon request.
Authors’ contributions
John Alexander Pulgarin Díaz: Data curation, formal analysis, investigation, methodology, software, visualisation, writing - original draft, writing - review & editing. Markus Melin: Conceptualisation of original idea, investigation, methodology, software, validation, writing - review & editing. Tiina Ylioja: Writing - review & editing. Päivi Lyytikäinen-Saarenmaa: Writing - review & editing. Heli Peltola: Funding acquisition, project administration, writing - review & editing. Olli-Pekka Tikkanen: Conceptualisation of original idea, validation, writing - review & editing.
Acknowledgements
Our thanks to Metsäkeskus (Forest Centre) for capturing, storing, curating and making the data sets used freely available. The computational analyses in RStudio were performed on servers provided by the Bioinformatics Center, University of Eastern Finland, Finland. The authors wish to acknowledge CSC – IT Center for Science, Finland, for computational resources for analysis and to reviewers who provided their valuable input.
Funding
This work was supported by the Research Council of Finland (former Academy of Finland) [grant numbers 337127 and 357906 for UNITE flagship]. We are also grateful to the FORES and LUMET Doctoral Programs at the University of Eastern Finland for their support and to the Finnish forest owners who supplied information to Metsäkeskus, making this research possible. The work for Markus Melin and Tiina Ylioja was partly supported by Luke project JollyForest (Kivametsä), funded by the Regional Council of Keski-Suomi.
References
Äijälä O, Koistinen A, Sved J, Vanhatalo K, Väisänen P (2019) Metsänhoidon suositukset. [Forest management recommendations]. Tapio.
Annila E (1969) Influence of temperature upon the development and voltinism of Ips typographus L. (Coleoptera, Scolytidae). Ann Zool Fenn 6: 161–208.
Battisti A, Larsson S (2015) Climate change and insect pest distribution range. In: Björkman C, Niemela P (eds) Climate change and insect pest. CABI International, Wallingford, pp 1–15. https://doi.org/10.1079/9781780643786.0001.
Bentz BJ, Jönsson AM, Schroeder M, Weed A, Wilcke RAI, Larsson K (2019) Ips typographus and Dendroctonus ponderosae models project thermal suitability for intra- and inter-continental establishment in a changing climate. Front For Glob Chang 2, article id 1. https://doi.org/10.3389/ffgc.2019.00001.
Blomqvist M, Kosunen M, Starr M, Kantola T, Holopainen M, Lyytikäinen-Saarenmaa P (2018) Modelling the predisposition of Norway spruce to Ips typographus L. infestation by means of environmental factors in southern Finland. Eur J For Res 137: 675–691. https://doi.org/10.1007/s10342-018-1133-0.
Brooks ME, Kristensen K, Benthem KJ Van, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Machler M, Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9: 378–400. https://doi.org/10.32614/RJ-2017-066.
Christiansen E, Bakke A (1988) The spruce bark beetle of Eurasia. In: Berryman AA (ed) Dynamics of forest insect populations: patterns, causes, implications. Springer, Boston, MA, pp 479–503. https://doi.org/10.1007/978-1-4899-0789-9_23.
Cowpertwait PS, Metcalfe AV (2009) Introductory time series with R. Springer. https://doi.org/10.1007/978-0-387-88698-5.
de Groot M, Ogris N, Diaci J, Castagneyrol B (2023) When tree diversity does not work: the interacting effects of tree diversity, altitude and amount of spruce on European spruce bark beetle outbreaks. For Ecol Manage 537, article id 120952. https://doi.org/10.1016/j.foreco.2023.120952.
Eriksson M, Neuvonen S, Roininen H (2007) Retention of wind-felled trees and the risk of consequential tree mortality by the European spruce bark beetle Ips typographus in Finland. Scand J For Res 22: 516–523. https://doi.org/10.1080/02827580701800466.
Finlex (2013) Laki metsätuhojen torjunnasta. [Forest Damages Prevention Act]. 1087/2013. Ministry of Justice, Helsinki.
Finnish Meteorological Institute (2022) Ilmastollinen vertailukausi 1991-2020. [Climatic reference period 1991-2020.] https://www.ilmatieteenlaitos.fi/ilmastollinen-vertailukausi. Accessed 18 February 2021.
Göthlin E, Schroeder LM, Lindelöw A (2000) Attacks by Ips typographus and Pityogenes chalcographus on windthrown spruces (Picea abies) during the two years following a storm felling. Scand J For Res 15: 542–549. https://doi.org/10.1080/028275800750173492.
Hantula J, Ahtikoski A, Honkaniemi J, Huitu O, Härkönen M, Kaitera J, Koivula M, Korhonen KT, Lindén A, Lintunen J, Luoranen J, Matala J, Melin M, Nikula A, Peltoniemi M, Piri T, Räsänen T, Sorsa J-A, Strandström M, Uusivuori J, Ylioja T (2023) Metsätuhojen kokonaisvaltainen arviointi : METKOKA-hankkeen loppuraportti. [Comprehensive assessment of forest destruction: final report of the METKOKA project]. Natural resource bioeconomy research 46/2023, Natural Resources Institute Finland (Luke), Helsinki. http://urn.fi/URN:ISBN:978-952-380-688-7.
Hardin J, Hilbe JM (2007) Generalized linear models and extensions. Stata Press.
Hilbe JM (2009) Logistic regression models. CRC press. https://doi.org/10.1201/9781420075779.
Hinze J, John R (2019) Effects of heat on the dispersal performance of Ips typographus. J Appl Entomol 144: 144–151. https://doi.org/10.1111/jen.12718.
Hlásny T, Krokene P, Liebhold A, Montagné-Huck C, Müller J, Qin H, Raffa K, Schelhaas M, Seidl R, Svoboda M, Viiri H (2019) Living with bark beetles: impacts, outlook and management options. From Science to Policy 8, European Forest Institute. https://doi.org/10.36333/fs08.
Hlásny T, König L, Krokene P, Lindner M, Montagné-Huck C, Müller J, Qin H, Raffa KF, Schelhaas MJ, Svoboda M, Viiri H (2021) Bark beetle outbreaks in Europe: state of knowledge and ways forward for management. Curr For Reports 7: 138–165. https://doi.org/10.1007/s40725-021-00142-x.
Hof AR, Svahlin A (2016) The potential effect of climate change on the geographical distribution of insect pest species in the Swedish boreal forest. Scand J For Res 31: 29–39. https://doi.org/10.1080/02827581.2015.1052751.
Hroššo B, Mezei P, Potterf M, Majdák A, Blaženec M, Korolyova N, Jakuš R (2020) Drivers of spruce bark beetle (Ips typographus) infestations on downed trees after severe windthrow. Forests 11, article id 1290. https://doi.org/https://doi.org/10.3390/f11121290.
Huuskonen S, Domisch T, Finér L, Hantula J, Hynynen J, Matala J, Miina J, Neuvonen S, Nevalainen S, Niemistö P, Nikula A (2021) What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For Ecol Manage 479, article id 118558. https://doi.org/10.1016/j.foreco.2020.118558.
Jönsson AM, Harding S, Bärring L, Ravn HP (2007) Impact of climate change on the population dynamics of Ips typographus in southern Sweden. Agric For Meteorol 146: 70–81. https://doi.org/10.1016/j.agrformet.2007.05.006.
Jönsson AM, Schroederb LM, Lagergrena F, Anderbrant O, Smitha B (2012) Guess the impact of Ips typographus – an ecosystem modelling approach for simulating spruce bark beetle outbreaks. Agric For Meteorol 166–167: 188–200. https://doi.org/10.1016/j.agrformet.2012.07.012.
Kärvemo S, Van Boeckel TP, Gilbert M, Grégoire JC, Schroeder M (2014) Large-scale risk mapping of an eruptive bark beetle–importance of forest susceptibility and beetle pressure. For Ecol Manage 318: 158–166. https://doi.org/10.1016/j.foreco.2014.01.025.
Kärvemo S, Huo L, Öhrn P, Lindberg E, Persson H (2023) Different triggers, different stories: bark-beetle infestation patterns after storm and drought-induced outbreaks. For Ecol Manage 545, article id 121255. https://doi.org/10.1016/j.foreco.2023.121255.
Kautz M, Dworschak K, Gruppe A, Schopf R (2011) Quantifying spatio-temporal dispersion of bark beetle infestations in epidemic and non-epidemic conditions. For Ecol Manage 262: 598–608. https://doi.org/10.1016/J.FORECO.2011.04.023.
Korhonen KT, Ahola A, Heikkinen J, Henttonen HM, Hotanen J-P, Ihalainen A, Melin M, Pitkänen J, Räty M, Sirviö M, Strandström M (2021) Forests of Finland 2014–2018 and their development 1921–2018. Silva Fenn 55, article id 10662. https://doi.org/https://doi. org/10.14214/sf.10662.
Kunegel-Lion M, Lewis MA (2020) Factors governing outbreak dynamics in a forest intensively managed for mountain pine beetle. Sci Rep 10, article id 7601. https://doi.org/10.1038/s41598-020-63388-8.
Lindman L, Ranius T, Schroeder M (2023) Regional climate affects habitat preferences and thermal sums required for development of the Eurasian spruce bark beetle, Ips typographus. Ecol Manag 544, article id 121216. https://doi.org/10.1016/j.foreco.2023.121216.
Louis M, Dohet L, Grégoire J (2015) Fallen trees’ last stand against bark beetles. For Ecol Manage 359: 44–50. https://doi.org/10.1016/j.foreco.2015.09.046.
Marini L, Økland B, Jönsson A, Bentz B, Carroll A, Forster B, Gregoire J, Hurling R, Nageleisen L, Netherer S, Ravn HP, Weed A, Schroeder M (2017) Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 40: 1426–1435. https://doi.org/10.1111/ecog.02769.
Mehtätalo L, Lappi J (2020) Biometry for forestry and environmental data: with examples in R. CRC press, Boca Raton. https://doi.org/10.1201/9780429173462.
Metsäkeskus (2021a) Metsänkäyttöilmoitukset. [Description about the forest stand use database with information on harvestings due to damage agents]. https://www.metsakeskus.fi/sites/default/files/document/tietotuotekuvaus-metsankayttoilmoitukset.pdf.
Metsäkeskus (2021b) Metsävarakuviot. [Description about the forest stand database]. https://www.metsakeskus.fi/sites/default/files/document/tietotuotekuvaus-metsavarakuviot.pdf.
Mezei P, Grodzki W, Blaženec M, Škvarenina J, Brandysova V, Jakuš R (2014) Host and site factors affecting tree mortality caused by the spruce bark beetle (Ips typographus) in mountainous conditions. For Ecol Manage 331: 196–207. https://doi.org/10.1016/J.FORECO.2014.07.031.
Müller M, Olsson P, Eklundh L, Jamali S, Ardö J (2022) Features predisposing forest to bark beetle outbreaks and their dynamics during drought. For Ecol Manage 523, article id 120480. https://doi.org/10.1016/j.foreco.2022.120480.
Netherer S, Matthews B, Katzensteiner K, Blackwell E, Henschke P, Hietz P, Pennerstorfer J, Rosner S, Kikuta S, Schume H, Schopf A (2015) Do water‐limiting conditions predispose Norway spruce to bark beetle attack? New Phytol 3: 1128–1141. https://doi.org/10.1111/nph.13166.
Netherer S, Panassiti B, Pennerstorfer J, Matthews B (2019) Acute drought is an important driver of bark beetle infestation in Austrian Norway spruce stands. Front For Glob Chang 2, article id 39. https://doi.org/10.3389/ffgc.2019.00039.
Nygren M (2011) The basic setting for growing trees. In: Rantala S (ed) Finnish forestry. Practice and management. Metsäkustannus Oy, Keuruu, pp 1–34. ISBN 978-952-569-462-8.
Økland B, Nikolov C, Krokene P, Vakula J (2016) Transition from windfall-to patch-driven outbreak dynamics of the spruce bark beetle Ips typographus. For Ecol Manage 363: 63–73. https://doi.org/10.1016/j.foreco.2015.12.007.
Pebesma E (2018) Simple features for R: standardized support for spatial vector data. R J 10: 439–446. https://doi.org/10.32614/RJ-2018-009.
Peltonen M (1999) Windthrows and dead-standing trees as bark beetle breeding material at forest-clearcut edge. Scand J For Res 14: 505–511. https://doi.org/10.1080/02827589908540815.
R Core Team (2023) R: a lenguage and environment for statitical computing. R Foundation for Statistical Computing, Vienna Austria.
Rundel C, Cetinkaya-Rundel M, Clyde M, Banks D (2021) statsr: Companion software for the coursera statistics with R specialization.
Stereńczak K, Mielcarek M, Modzelewska A, Kraszewski B, Fassnacht FE, Hilszczański J (2019) Intra-annual Ips typographus outbreak monitoring using a multi-temporal GIS analysis based on hyperspectral and ALS data in the Białowieża Forests. For Ecol Manage 442: 105–116. https://doi.org/10.1016/j.foreco.2019.03.064.
Tikkanen O, Lehtonen I (2023) Changing climatic drivers of European spruce bark beetle outbreaks: a comparison of locations around the Northern Baltic Sea. Silva Fenn 57, article id 23003. https://doi.org/10.14214/sf.23003.
Triviño M, Potterf M, Tijerín J, Ruiz-Benito P, Burgas D, Eyvindson K, Blattert C, Mönkkönen M, Duflot R (2023) Enhancing resilience of boreal forests through management under global change: a review. Curr Landsc Ecol Reports 8: 103–118. https://doi.org/10.1007/s40823-023-00088-9.
Venäläinen A, Lehtonen I, Laapas M, Ruosteenoja K, Tikkanen OP, Viiri H, Ikonen V, Peltola H (2020) Climate change induces multiple risks to boreal forests and forestry in Finland: a literature review. Glob Chang Biol 26: 4178–4196. https://doi.org/10.1111/gcb.15183.
Wermelinger B (2004) Ecology and management of the spruce bark beetle Ips typographus – a review of recent research. For Ecol Manag 202: 67–82. https://doi.org/10.1016/j.foreco.2004.07.018.
Wermelinger B, Seifert M (1998) Analysis of the temperature dependent development of the spruce bark beetle Ips typographus (L.) (Col., Scolytidae). J Appl Entomol 122: 185–191. https://doi.org/10.1111/J.1439-0418.1998.TB01482.X.
Total of 51 references.
