Eva Ring  ,
Märtha Wallgren,
Erland Mårald,
Per Westerfelt,
Line Djupström,
Aron Davidsson,
Johan Sonesson
,
Märtha Wallgren,
Erland Mårald,
Per Westerfelt,
Line Djupström,
Aron Davidsson,
Johan Sonesson
Forest roads in Sweden – infrastructure with multiple uses and diverse impacts
Ring E., Wallgren M., Mårald E., Westerfelt P., Djupström L., Davidsson A., Sonesson J. (2024). Forest roads in Sweden – infrastructure with multiple uses and diverse impacts. Silva Fennica vol. 58 no. 4 article id 24021. https://doi.org/10.14214/sf.24021
Highlights
- The extensive network of forest roads in Sweden facilitates both forestry transportation and other activities including recreation, berry-picking, hunting, tourism, and firefighting
- The multiple uses of roads can lead to conflicts when the interests between stakeholders and forest owners collide
- Despite its extent, research into the social, ecological, and environmental consequences of Sweden’s forest road network is limited.
Abstract
Since the 1950s, more than 200 000 km of roads have been built in Sweden’s forests, making them easily accessible and open to multiple uses. The aim of this study was to review the impacts of forest roads in Sweden from a broad perspective encompassing social, ecological, and environmental factors. The Swedish case is interesting because it has an extensive network of permanent forest roads which were built primarily for forestry-related transportation but are currently used by many other stakeholders for many different purposes. Forest roads not only facilitate transportation of wood, machinery, personnel, and equipment into and out of the forest but also enable emergency response to wildfires and support berry and mushroom picking, hunting, recreation, tourism, and access to second homes. The roads increase the opportunities for members of the public to experience forests in various ways. Conflicts arise when different interests collide, for example when the interests of the forest owner clash with those of commercial berry-picking companies, tourism entrepreneurs, or reindeer (Rangifer tarandus L.) herding. Forest roads may have ecological impacts such as barrier and disturbance effects, fragmentation or loss of habitats, altering fauna movement patterns, and changing the composition of plant and insect species. The environmental impacts of forest roads relate to, among other things, hydrology, water quality, and erosion. Predicted changes in the climate are likely to place new demands on Swedish forest roads but, despite their extent, this review shows that there is only a small amount of rather fragmented research on their social, ecological, and environmental consequences. Overall, few studies appear to cover both social and ecological/environmental factors and their interactions, either in Sweden or elsewhere. This review provides examples of such interactions in the case of Sweden, and suggests that more research into these and the specific social, ecological, and environmental factors involved is warranted.
Keywords
insects;
forestry;
recreation;
reindeer;
water;
plants;
wildlife
-
Ring,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden
 https://orcid.org/0000-0002-8962-9811
E-mail
eva.ring@skogforsk.se
https://orcid.org/0000-0002-8962-9811
E-mail
eva.ring@skogforsk.se
-
Wallgren,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden; Swedish University of Agricultural Sciences, Department of Wildlife, Fish, and Environmental Studies, 907 36 Umeå, Sweden
https://orcid.org/0000-0002-3172-4496
E-mail
martha.wallgren@skogforsk.se
-
Mårald,
Umeå University, Department of Historical. Philosophical and Religious Studies, 901 87 Umeå, Sweden
https://orcid.org/0000-0002-2291-9910
E-mail
erland.marald@umu.se
-
Westerfelt,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden
https://orcid.org/0000-0003-2040-8305
E-mail
per.westerfelt@skogforsk.se
-
Djupström,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden; Swedish University of Agricultural Sciences, Department of Wildlife, Fish, and Environmental Studies, 907 36 Umeå, Sweden
https://orcid.org/0000-0003-4536-7765
E-mail
line.djupstrom@skogforsk.se
-
Davidsson,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden
https://orcid.org/0009-0007-2736-8656
E-mail
Aron.davidsson@skogforsk.se
-
Sonesson,
Skogforsk (The Forestry Research Institute of Sweden), Uppsala Science Park, 751 83 Uppsala, Sweden
https://orcid.org/0000-0002-2018-7496
E-mail
Johan.sonesson@skogforsk.se
Received 25 March 2024 Accepted 26 August 2024 Published 9 September 2024
Views 195487
Available at https://doi.org/10.14214/sf.24021 | Download PDF

1 Introduction
One of the largest structural changes to the forest landscape in Sweden since the mid-20th century has been the construction of the forest road network. There are around 210 000 km of permanent forest roads in Sweden (Bernes and Lundgren 2009; Eliasson 2011), corresponding to more than five laps around the equator. Nonetheless, further expansion of the forest-road network is expected in future (Bergqvist et al. 2023). Unlike many other countries in the sparsely populated northern boreal coniferous forest belt, where such roads can be more temporary or of poorer quality (Kuklina et al. 2020; Larson and Rew 2022), the vast majority of forest roads in Sweden are permanent and designed to cope with heavy traffic for most of the year. The expansion of forest roads took off in the 1950s in connection with a shift in the forest management regime towards a large scale and mechanised clearcutting system (Eliasson 2011; Mårald et al. 2017). In this system, forest roads are used not only during final felling, but regularly throughout the whole forest rotation in conjunction with various management measures. An individual forest road may reach several stands at different stages of the rotation, increasing the frequency with which it is used. The road network benefits forest management by making it easier to implement the right measures at the right time (Filipsson 2001). Access to the forest is also central to coping with disturbance events such as insect outbreaks, wildfires, and storm felling (Kolström et al. 2011).
Today, there are many demands on the functions provided by forests in Sweden including timber production, biodiversity, water quality, climate change mitigation, and social activities (Sandström et al. 2011; Lundmark et al. 2014). Forest roads may influence several of these functions. The forest road network is indispensable for forestry transportation. However, as a large proportion of forest roads are open to the public, they also increase general accessibility to forests and open them up for different uses and nature experiences, through activities such as hiking, cycling, berry- and mushroom picking, hunting, and fishing (Fig. 1). In Sweden, the Right of Public Access allows people to access and use privately owned forests as long as nature is not harmed and the landowner´s interests are not compromised (Sténs and Sandström 2013). In the northern half of Sweden, Sámi people herd their reindeer (Rangifer tarandus L.) in forests and alpine heathlands for grazing and breeding. Over recent decades, this transhumance system has been increasingly affected by changes in land cover, most of which relate to forestry, in which forest roads are a factor (Sandström et al. 2003; Horstkotte 2013). Thus, there are many stakeholders with an interest in forest roads in Sweden.

Fig. 1. Illustration of the multiple uses of forest roads in Sweden encompassing forestry transportation, firefighting, and public access to the forest and its benefits. Left panel: timber awaiting transportation (top), forest road providing access to a house (middle), and horse riding on a forest road (bottom). Right panel: burnt forest one year after a severe wildfire (firefighting) (top), hunting tower by a forest road (middle), and a berry-picking site (bottom). Photos by E. Ring and M. Wallgren.
Globally, numerous scientific studies have investigated the ecological and environmental impacts of building, maintaining, and using forest roads (Lugo and Gucinski 2000; Trombulak and Frissell 2000; Gucinski et al. 2001; Robinson et al. 2010). Many of their findings are broadly applicable to any region of the world. It is well-known that forest roads have consequences for flora, fauna, and hydrology (Robinson et al. 2010; Boston 2016; Kastridis 2020). The presence of forest roads means that nearby forest environments and ecosystems will be affected by traffic, road maintenance, and increased human presence and activity (Fig. 2). Forest and public roads can have a wide range of impacts such as barrier and disturbance effects, habitat loss or fragmentation, direct animal mortality caused by vehicle collisions, changing fauna movement patterns, reducing gene flow, enabling the dispersal of new predators and competitors, changing the composition of plant and insect species, altering hydrological patterns, increasing erosion and sediment transport, and pollution (Forman and Alexander 1998; Trombulak and Frissell 2000; Coffin 2007; Benítez-López et al. 2010; Boston 2016; Kastridis 2020; Oliveira et al. 2020). How these play out in a specific geographical location depends on the regional or national context.

Fig. 2. Illustration of some of the ecological and environmental factors that can be affected by forest roads, including wildlife, plants and insects, hydrology, and reindeer. Left panel: capercaillie on a forest road (top), flowering plants in a road verge (middle), a bee pollinating maple flowers (bottom). Right panel: roadside ditch draining water (top), road culvert beneath a forest road (middle), and reindeer moving along a forest road (bottom). Photos by E. Ring, L. Djupström, M. Wallgren.
The aim of this review is to provide a broad overview of the multiple uses and social, ecological, and environmental consequences of forest roads in Sweden, and to identify or highlight possible interactions between different factors, gaps in knowledge, and future challenges. Sweden presents an interesting case for this type of analysis due to its large network of forest roads and the many stakeholders involved. The study reviews the literature around the following focal themes: social values, reindeer herding, wildlife, plants, insects, and water.
2 Material and methods
The character of this review shows similarities to both the integrative review approach described by Snyder (2019) and the scoping study approach discussed by Arksey and O’Malley (2005). The purpose was to provide an accessible and concise narrative description of the available research on several topics relating to Swedish forest roads and to analyse this research. The aim was to highlight the multiple roles of forest roads at a more general level, rather than providing a complete analysis of each selected topic. The expertise of the author team spans a wide range of disciplines which has influenced the selection of topics. Information about forest roads within the selected topics were collected from various sources. The search engine Google Scholar and the Web of Science databases were used to find relevant scientific articles, and information was collected from books, reports, official statistics, reference lists, and authors´ own literature collections.
Despite our focus on forest roads in Sweden, studies about other types of roads and from other parts of the world, particularly Northern Europe and North America, were included if found to be relevant. The relevance of the literature was determined by considering factors such as geographic region, topography, target species or species groups, and similarity to other types of roads and linear clearings. The search process was not systematic. To identify additional literature on forest roads in Sweden, Norway, Finland, and Denmark, a broad literature search was performed on 7 December, 2023, using the databases Web of Science Core Collection, Scopus, CABI: CAB Abstracts, and GreenFILE. The search string was (“forest road”* OR “forestry road”* OR “logging road”* OR “gravel road”*) AND (Swed* OR Norw* OR Finland* OR Finnish OR Scandinavia* OR Nordic* OR Fennoscand* OR Denmark* OR Danish) (* meaning truncation). The search terms were searched for in the title, abstract, and keywords of articles published in English, without any restriction by publication year. These searches identified a few additional articles as worthy of inclusion in this study.
2.1 Forest roads in Sweden – a brief overview
Forestry in Sweden is a nationwide and transport-intensive industry that uses the entire Swedish road network all year round, from small gravel roads to large motorways. More than 90% of the transport work of forest biomass (transported biomass of timber, pulpwood, energy wood, and logging residue multiplied by distance) is performed on public roads and the remaining part on private roads (Davidsson et al. 2023). However, most forest biomass transports (in 2020, 87% of the transports) start on private roads, half of which were forest roads (Davidsson et al. 2023; Skogskunskap 2024). Truck transportation of forest biomass has approximately quadrupled between 1954 and 2020 (Skogsbrukets motortransportkommitté 1957; Davidsson et al. 2023). In 2020, 72.2 million tonnes of forest biomass were transported by truck in Sweden (Davidsson et al. 2023). Information about the Swedish road network is stored in a national database (National Road Database, https://www.nvdb.se/).
Implementing current forest management strategies efficiently requires a road network of a certain density and standard. The forest road network helps to reduce energy consumption and costs by shortening off-road transport distances and total transport time. Forest roads are also used to transport forestry machinery such as harvesters, forwarders and site preparation equipment, staff, seedlings, and various types of equipment.
Forest roads in Sweden are typically built for a maximum vehicle gross weight of 64 tonnes. They are classified by function, accessibility, and geometric standard, with different classes for each characteristic (Skogskunskap 2024). Forest roads are usually built from material excavated from the roadside and/or locally established gravel pits (the road terrace) and specially produced gravel- or bedrock material (the road surface). Forest road maintenance includes a range of practices such as grading of the road surface, adding gravel or bedrock material to strengthen the road surface, cleaning roadside ditches, cutting roadside vegetation, and adding salt to reduce dusting and consolidate the road surface (Skogskunskap 2024). Maintenance varies in intensity depending on how frequently the road is used. Construction and management of forest roads in Sweden is generally the responsibility of a specified organisation such as a joint property association, or co-managed by the landowners concerned (Bergqvist et al. 2023).
This review focuses on permanent forest roads in Sweden. Temporary winter roads that are sometimes used in northern Sweden and strip roads for terrain vehicles are not included. The forest roads considered here are always privately owned, unpaved and run mainly through production forest. Traffic intensity is generally low compared with paved motorways but increases in conjunction with forestry operations. Some of the roads are open for public access while others are closed depending on landowners’ decisions. In the more populated areas of Sweden, a substantial fraction of gravel roads are multipurpose roads, leading to farms, houses, summer houses, or farm fields, and serve other uses besides forestry. The results of this study are probably also applicable to most of these roads, if they run through forest and are to some extent used for forestry transportation. In Fig. 3, some terms used in this article to describe different forest road elements are graphically illustrated. In addition, “roadside” refers to the area (or a part of the area) between the travel way and the forest edge.
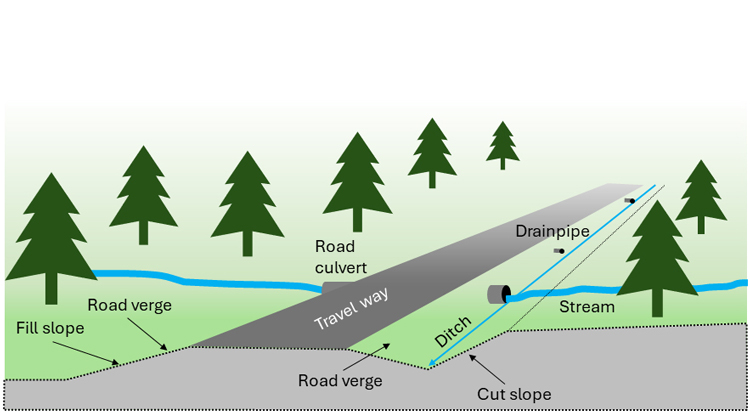
Fig. 3. Schematic design of a forest road running across a gentle hillslope. Forest roads are constructed to prevent water from accumulating in the road. The travel way is compacted to divert precipitation to roadside ditches, which also collect and transport water coming from the cut slope. Drainpipes transport water from upslope areas across the road.
3 Results
3.1 Social values
3.1.1 Access to game, fish, berries, and mushrooms
Forest roads are important to many people in enabling them to access and use the forest in various ways. In Sweden, there is a long tradition of locally embedded usufructuary rights which provide access to privately owned forests for individuals and groups who are not the landowner, to use them for grazing, hunting, reindeer herding, and gathering fuel, berries, or materials for construction and handicrafts. For example, in the past four years nearly 300 000 hunting permits have been registered every year (Swedish Environmental Protection Agency 2024). During the 20th century many of the usufructuary rights were regulated through the Right of Public Access, the Reindeer Management Right, and hunting legislation (Sténs et al. 2016). The Right of Public Access stipulates that the public may roam on almost all land, irrespective of ownership, as long they do not interfere with the landowner´s interests or cause any damage to the land and what grows, lives, or stands on it. It also provides leeway for picking berries, mushrooms, common wildflowers, and herbs (Åslund 2008). Forest roads are vital for accessing game, fish, berries, mushrooms, and other benefits. In Sweden about half of all visits to the forest are carried out by car, where forest roads are the primary source of access to forests and means of reaching locations and resources that lie deeper within them (Hannerz et al. 2016; Gundersen and Vistad 2016).
It is fair to say that access to forests is contested, as debates about forest private property rights and conflicting interests in the use of forest landscapes illustrate (Hellström 2001; Sténs and Mårald 2020). For instance, large scale commercial berry harvesting has long been disputed, the debate recently intensifying due to an influx of foreign professional berry harvesters (Sténs and Sandström 2013). Aside from complaints about camp sites and littering, the debate has focused on the boundaries of the Right of Public Access and to what extent commercial actors can commercialise non-timber forest products without any agreement with the forest owner. Closing forest roads, using gates, is one response to this situation.
3.1.2 Recreation and tourism
In Sweden, there is a strong tradition of using forests for recreation (Hörnsten and Fredman 2000). According to earlier research, on average Swedes visit a forest at least every other week (Lindhagen and Hörnsten 2000). Most regular recreational use of forest takes place in people’s immediate surroundings (Hannerz et al. 2016). Due to urbanisation, the recreational use of forests is highly concentrated in urban forests or natural reserves close to major cities. Immigrants, who are more likely to live in cities, are significantly less frequent visitors to forest areas (Lisberg Jensen and Ouis 2014).
Forest management in urbanised areas has been adapted to accommodate recreation and promote “social values” (Hannerz et al. 2016; Sténs et al. 2016). This entails the construction of marked pathways, walking and bicycle paths, and resting places, and managing stands to provide open views, increase the sense of security and connection to nature, and promote public health and meaningful public involvement (Holgén et al. 2000; Meier 2013; Hannerz et al. 2016). In these areas, forest roads have often been improved specifically to suit such recreational purposes (Gundersen and Vistad 2016).
From a tourism perspective, forests are seen as valuable amenities (Hall and Müller 2004). In rural areas, tourism has been promoted as a vehicle for local development and generating new forms of income through providing accommodation and camping facilities or opportunities for health and wellbeing (Almstedt et al. 2014). An increasing number of forest owners have diversified their businesses, combining forest production with tourism and other remunerative activities (Zhang et al. 2022). In Sweden, second homes, often located in remote areas on the coasts or in rural and mountainous areas, are common. More than half the population owns or has access to a second home (Hall and Müller 2004). Mobility and access to specific locations and attractions are central to tourism (Hall and Müller 2004). Hence, the forest road network plays an important role in rural tourism.
3.1.3 Aesthetics and cultural heritage
Only a few studies mention people’s experiences of forest roads. While forest visitors ideally wish to use moderately prepared forest paths for walking, in practice they walk on forest roads (Gundersen and Frivold 2008). Forest roads that are adapted to the terrain are well accepted by users, but what was considered to be more brutal road constructions (i.e., newly built roads that cut through the terrain) are poorly accepted (Lind et al. 1974). The visual character of a forest road is thus influenced by factors like road curvature, ditches, road surface, and surrounding vegetation, and an overgrown old forest road is particularly appreciated (Gundersen and Vistad 2016). Substantial traces of logging and off-road transportation, like deep wheel tracks, are aesthetically displeasing (Lindhagen and Hörnsten 2000; Gundersen et al. 2016). In what is experienced as wilderness, the presence of paths, fire sites and constructions relating to outdoor recreation were acceptable, while roads and winter tracks for timber harvesting and transportation are disturbing and detract from the experience (Hallikainen 1998).
Increasing attention is being paid to the cultural heritage that is present in forests (Sténs et al. 2016). In Sweden, the protection of cultural heritage is regulated by several laws and permission from the state is required before a new forest road can be built in an area that contains known ancient remnants (Swedish National Heritage Board 2010). However, only a small proportion of cultural remnants has been identified and mapped. Thus, many remnants do get damaged by forestry, for example when building forest roads, before they have even been discovered and documented (Berg and Gustafson 2013).
3.1.4 Wildfire risk and control
Forest fire is one of the most important natural disturbances that has shaped the ecology of boreal forests. In Sweden, efforts to control forest fires have been so successful that certain fire-dependent species are now threatened (SLU Swedish Species Information Centre 2024). Forest roads provide important infrastructure for firefighting but may at the same time contribute to more human-induced ignitions since they provide people with easy access to forest landscapes. Narayanaraj and Wimberly (2012) studied spatial patterns of wildfire ignitions in Washington State and found that human-induced fires were concentrated close to roads while the greatest area of burning was caused by large lightning-induced fires in roadless areas. The human-induced fires close to roads were mostly small because they were reported sooner and easier for the fire fighters to control. Narayanaraj and Wimberly (2012) concluded that, from the standpoint of the total area burned, the effect of forest roads on restricting fire size is likely greater than the impact of roads on increasing fire ignitions.
In Sweden, the number of forest fire ignitions varied between approximately 1000 and 4000 per year during 1998–2014 and the total area burnt varied between a few hundred to around 6000 ha per year, with the exception of one large fire of 13 000 ha in 2014 (Ou 2017). More than 70% of the fires were less than 0.1 ha in size. Of the total number of forest fires, only 7% were known to be ignited by lightning and 52% by human activity. Campfires and grilling (11%) and children playing with fire (9%) were the most common reasons for human-caused ignitions. The cause of ignition was unknown for 41% of the fires. Analysing a dataset of 124 000 wildfire ignitions in Sweden over 25 years, Sjöström and Granström (2023) found a strong positive correlation between population density and number of ignitions. However, fires larger than 10 ha mostly occurred in sparsely populated areas and were more often ignited by lightning or heavy machinery.
The forest road network is important for wildfire control as it gives fire brigades rapid access to the forest landscape, making it possible to control fires when they are still small and easy to put out (Björheden 2019). Furthermore, forest roads may function as firebreaks for low-intensity ground and surface fires, although rapidly spreading high-intensity fires are normally not stopped by a road (Gucinski et al. 2001). Recent developments in road planning and construction have sought to improve the usefulness of forest roads for firefighting (Laschi et al. 2019; Thompson et al. 2021).
Pinto et al. (2020) analysed the occurrence and size of forest fires in relation to population and road densities in Sweden and found more frequent fires in landscapes with high population and high road density. On the other hand, fires were larger in landscapes with low road density. The results of Pinto et al. (2020) suggest that earlier findings by Narayanaraj and Wimberly (2012) and Gucinski et al. (2001) are equally valid for Swedish conditions.
3.1.5 Sanitation and salvage logging
Forest roads are crucial both for sanitation logging, to mitigate insect outbreaks, and for salvage logging to salvage wood value after disturbances. In a simulation study on the effects of sanitation logging of wind-felled trees on subsequent disturbance by spruce bark beetle (Ips typographus L.), it was concluded that focusing on sanitation logging along forest roads created a “firebreak effect” on the spread of bark beetle and was moderately effective in reducing landscape-scale bark beetle disturbance (Dobor et al. 2020).
3.1.6 Concluding remarks and research gaps
Forest roads are important for people’s access to and use of forests. This applies to berry picking, hunting, and reindeer herding as well as recreation and access to second homes. While there is much research examining these uses individually, there is little that examines the multiple functions of forest roads themselves. The same applies to research on forestry and aesthetic values. In studies of how people experience the forest and forest management measures, forest roads are often used for transport and walking. However, the focus is on the surrounding forest, while the road itself is taken for granted and becomes part of the background. There is an urgent need to map and document cultural heritage prior to forestry operations, to be able to minimise forestry´s negative impact on it.
In the future, forest roads may become even more important for dealing with forest disturbances because of the increased risk of wildfire, insect damage, and wind felling associated with climate change (Ikonen et al. 2017; Pureswaran et al. 2018; Venäläinen et al. 2020).
3.2 Reindeer herding
Sámi reindeer owners herd their animals in the reindeer management area of Sweden, Norway, and Finland. Along with direct benefits such as meat and other material products, reindeer herding has a cultural value as it is a traditional livelihood amongst indigenous Sámi people and also attracts tourism.
Reindeer require access to a wide range of forests and alpine heathlands, depending on the season, to graze and breed. Summer is spent grazing in the mountains and after mid-summer herders gather their animal stock to earmark the calves. In the autumn culling (round-ups) herds are gathered again and split up into smaller groups after which the migration to winter grazing grounds begins. This transhumance system has been increasingly affected by changes in land cover over recent decades, most of which are directly related to forestry: for example changes in the age distribution of forests and reduced areas of winter pasture (Sandström et al. 2003; Horstkotte 2013). Since the 1960s, the forest road network in northern areas has expanded considerably as a result of increased logging and tourism (Kivinen 2015).
Research on how reindeer herding is affected by forest roads and other infrastructure has focused on factors such as disturbance and avoidance behaviour. Previous research has shown that all reindeer (wild or domesticated) avoid forest roads, although the avoidance pattern may shift between seasons and years (Vistnes and Nellemann 2008; Skarin and Åhman 2014; Turunen et al. 2020). The construction of roads and the increased traffic and tourism they entail is highlighted as a detrimental disturbance, particularly during calving and round-ups.
In Norway, the abundance of wild reindeer was found to be substantially higher in undisturbed areas which are not affected by roads (>5 km from infrastructure), in both winter and summer including the insect harassment period (Nellemann et al. 2003). Wild reindeer (caribou) in Canada have also been found to show avoidance behaviour in response to forest roads (Courtois et al. 2008; Pinard et al. 2012; Boan et al. 2014; Vanlandeghem et al. 2021). One model-based study found that forest cover was the best measure of a habitat’s suitability for caribou, and that including logging roads and the presence of wolf (Canis lupus L.) improved the model’s accuracy (Boan et al. 2014). Skarin and Åhman (2014) reviewed how domesticated reindeer respond to various disturbance sources, including logging roads. The authors compared studies on domesticated reindeer and those focusing on wild reindeer and North American caribou. Despite extensive domestication, the Sàmi reindeer exhibited comparable large-scale avoidance behaviour to that of wild reindeer and North American caribou, with occasional variations in strength and response. Forest roads may disturb reindeer migration, leading the animals to wrong pastures and making it more difficult for herders to find them. Roads that provide access to the herds may also attract poachers and reindeer thieves. At the same time, forest roads can facilitate the practical work of reindeer herders, for example when gathering reindeer and organising supplementary winter feeding (Turunen et al. 2020 and references cited therein).
3.2.1 Concluding remarks and research gaps
Research on the effect of forest roads on reindeer herding is fragmented. The response of domesticated reindeer to various disturbance sources, including forest roads, needs to be addressed at different spatial and temporal scales. For example, migration and movement between seasonal ranges and feeding areas must be assessed at both regional and local scale and across the herding year to cover all the activities involved in reindeer herding. Using different scales is important to fully detect the influence of forest roads and identify the circumstances under which roads may hinder or force animals to choose a certain habitat. These studies should address the animals’ migration patterns and behavioural response and draw on the traditional knowledge of Sàmi people.
3.3 Wildlife
There is a deficiency of studies on how forest roads affect wildlife in Sweden. While more than 500 scientific articles were found on the topic for boreal and temperate forests in the northern hemisphere, fewer than 20 originated in Sweden. Given the lack of studies from Sweden, the following overview regarding forest roads and wildlife in Sweden is largely based on studies from other countries in ecosystems that are considered relevant to Sweden.
If not decommissioned (Larson and Rew 2022), forest roads constitute a permanent change to continuous forest habitat structures that may affect mobile organisms´ access to nearby ranges (Yemshanov et al. 2022). This applies to the forest road network in Sweden. Animals´ responses to linear features, such as forest roads, depend on the width of the clearing and intensity of motor-vehicle traffic among other factors. Small, low-traffic roads are generally expected to have small impacts, while high-traffic roads have higher impacts (Wallgren et al. 2009).
Roads and other infrastructure may affect wild animals in various ways, for instance by establishing barriers, inducing disturbances, or modifying habitats (Boston 2016). Possible effects include habitat loss or fragmentation, direct mortality through collisions, altered movement patterns, reduced gene flow and genetic diversity of populations, dispersal of new predators and competitors, modification of the physical and chemical environment (e.g. by altering hydrology and pollution), and increased access for humans (Forman and Alexander 1998; Trombulak and Frissell 2000; Coffin 2007; Benítez-López et al. 2010; Steyaert et al. 2011; Boston 2016; Oliveira et al. 2020). Road infrastructure may also increase habitat heterogeneity by creating gaps, linear openings and tree layering, increasing field layer vegetation along roadsides, and establishing corridors for faster or more efficient movement (Mackenzie 1946; Haddad et al. 2003; Beyer et al. 2013). The effects of roads on wildlife generally impact a considerably wider zone than the road prism and adjacent cleared areas (Forman and Alexander 1998; Soultan et al. 2021). One difficulty in interpreting the effects of road density on animal abundance is that road densities are often correlated with other effects, such as habitat loss (Fahrig and Rytwinski 2009; Yemshanov et al. 2022). According to Findlay and Houlahan (1997), wetland species richness of herptiles and birds may be significantly reduced at a distance of 2 km from the nearest road or logging site, probably in conjunction with secondary impacts such as changes to hydrology or forest continuity.
3.3.1 Forest grouse
Birds respond to disruption of forest structure differently depending on their specific habitat demands, with those species preferring old, closed-canopy forests being most negatively affected by the presence of forest roads (Bayne et al. 2016). Capercaillie (Tetrao urogallus L.) prefer tree stands >150 m from forest roads for leks (Bolibok et al. 2014). Chick-rearing females of black grouse (Lyrurus tetrix L.) avoid forest roads, tracks and walking paths (Patthey et al. 2012). One study in Sweden showed that capercaillie moved faster and more directionally when they were close to access roads to wind turbines than when they were far from them suggesting that, despite sparse traffic, the roads stressed the birds (Kämmerle et al. 2021).
Forest roads increase the species richness of local bird communities (Mackenzie 1946), and this is usually attributed to the increase in habitat heterogeneity and thus suitability for a higher number of species (Šálek et al. 2010). Residual habitats such as forest edges and roadsides have been shown to be important for some bird species in Sweden (Berg and Pärt 1994). The openness associated with forest roads and edges in general may increase the distance and speed of bird song transmission (Yip et al. 2017), which may affect social interactions as well as population assessments of birds.
The deterrent effect of forest roads on predators may be beneficial for forest grouse. An experiment using artificial ground nests revealed that predation was lower at <1 m distance from forest logging roads than at distances of >50 m from them (Yahner and Mahan 1997). Similarly, Matysek et al. (2021) found that tourist trails and unpaved roads reduced the predation rate of artificial hazel grouse (Tetrastes bonasia L.) nests. Comparable results have been found for other cleared areas and explained as relating to the ability of vegetation to provide hiding places for the predator (Hummel et al. 2017).
The presence of humans can have a deterrent effect on birds. In bird communities in Finland the presence of ground nesting species was negatively affected by the frequency of human hikers on forest paths (Kangas et al. 2010). Forest roads that increase the presence of humans in previously less disturbed forest habitats can be expected to have the same effect.
3.3.2 Herptiles
A vast amount of scientific literature reports significant effects of road mortality on local populations of amphibians (deMaynadier and Hunter Jr. 1995; Elzanowski et al. 2009 and references therein), particularly in close proximity to wetlands (Boston 2016). Aside from the risk of being run over by motor vehicles, there is an increased risk of predation by exposure on the open surfaces created by roads (deMaynadier and Hunter Jr. 1995). Although at considerable risk of road mortality (Chmielewski 2016), snake responses to roads seem equivocal (Baxley and Qualls 2009; Baxley et al. 2011). It is possible that warm temperatures and access to food attract snakes to road surfaces and roadsides.
In a study from Maine, USA, deMaynadier and Hunter Jr (2000) found that forest roads restricted the movements of salamanders, but not of frogs and toads. Natal dispersal of salamanders was particularly negatively affected and more so by larger roads than small, less trafficked roads (deMaynadier and Hunter Jr 2000). On the other hand, toads may exhibit higher travelling speeds on forest roads than in the forest, possibly due to the effect of warmer temperatures on roads (Deguise and Richardson 2009). The road-effect zone for woodland salamanders was shown to extend 35 m to either side of relatively narrow, low-use forest roads (Semlitsch et al. 2007).
The barrier effect of roads may cause local extinctions of amphibian populations through isolation (deMaynadier and Hunter Jr. 1995). Even moderate genetic, demographic, and environmental stochasticity may have detrimental consequences for fragmented populations.
Other features associated with forestry and forest roads may be important for herptiles, for example ditches (Dibner et al. 2014; Homyack et al. 2016). However, in the eastern USA, Johnson et al. (2016) identified slightly lower species richness of amphibians in roadside ditches than in small natural streams.
3.3.3 Mammals
Roads may induce avoidance or barrier effects for mammals (Coffin 2007; McGregor et al. 2008; Boston 2016; Soultan et al. 2021). Pattison and Catterall (2019) conclude that narrow (i.e., 8 m) non-road seismic lines in boreal forest environments only marginally reduced the propensity to cross in some mammal groups: hares, medium sized carnivores and large and medium sized herbivores. On the other hand, movements along (rather than across) linear clearings were preferred by most mammal groups covered by the study, except small mammals (Pattison and Catterall 2019). Small mammals’ tendency to avoid linear forest clearings has been explained by the higher exposure and different ground surfaces (including vegetation and soil properties) the clearings present than is found in intact forest vegetation (Haddad et al. 2003; Pattison and Catterall 2019). Research by McGregor et al. (2008) showed that rodents strongly avoided the road surface itself, rather than emissions such as noise and lights from the vehicles on the roads. Linear movements were most pronounced in the mammal groups with the highest energetic requirements: large carnivores and herbivores, followed by medium sized carnivores and herbivores (Pattison and Catterall 2019).
Two studies from Canada indicate that clearings along roads and other linear features may increase the densities of shrubs and saplings, which may attract large herbivores (Pattison and Catterall 2019; Darlington et al. 2022). Similar results have been shown for moose (Alces alces L.) in Sweden and Norway, who in both cases avoided forest roads but not power lines (Eriksen et al. 2009; Neumann et al. 2013; Bartzke 2014). However, in Norway moose showed no selectivity related to roads in wintertime, which Bartzke (2014) suggests was an effect of higher food availability close to roads counteracting disturbance from traffic. Loosen et al. (2021) have also observed higher browsing occurrence near secondary roads (including forest roads) but lower browsing along primary roads (paved roads and highways). They argue that the greater amounts of highly selected deciduous forage attracted moose to roads in general but high traffic speeds and volumes and barrier effects from fences counteracted the effect of increased forage availability on primary roads. Further, the presence of young commercial forest intensified the pattern of increased browsing on pine close to secondary roads when compared with, for example, the presence of clearcuttings which generally provide less browse (Loosen et al. 2021), indicating that moose react to overall food access and/or disturbance levels in proximity to forest roads.
In Finland, browsing damage by moose was lower close to roads than further away (Nikula et al. 2019). Similar results were found by Gicquel et al. (2020) in Sweden, but they also showed that size of road was important. Unlike large roads, on forest roads the presence and level of moose browsing damage to pine decreased at increasing forest road length (assessed for 1×1 km squares) (Gicquel et al. 2020). However, these lower levels of damage close to forest roads may very well be due to a dilution effect rather than road avoidance, because in Sweden most young forests are reachable via forest roads and there should thus be a positive correlation between forest road length and amount of young forest available for browsing.
Although there is a connection between large herbivore browsing and distance to roads, there may be more complexity behind this pattern. Loosen et al. (2021) found that moose browsing on rowan (Sorbus aucuparia L.), a highly selected forage species, occurred further away from secondary roads if wolf territories were present than if they were not. Wolves use roads to travel (Zimmermann et al. 2014) and thus the predation risk from wolves can be higher close to roads, which may explain why prey species such as large herbivores avoid roads (Eriksen et al. 2009; DeMars and Boutin 2018; Francis et al. 2021; Vanlandeghem et al. 2021). However, resting wolves prefer sites with thickets located away from forest roads, suggesting that wolves also seek concealment during periods of low activity (Bojarska et al. 2021). Zimmermann et al. (2014) and Thorsen et al. (2022) found that forestry roads were frequently used by wolves in Scandinavia, but avoidance of human settlements had a stronger impact on their movement patterns at some scales. By contrast, brown bear (Ursus arctos L.) in Norway have been found to avoid forest roads but may tolerate being in their vicinity if vegetation nearby these roads is very dense (Steyaert et al. 2011).
Ambiguous results have been reported regarding the effects of forest roads on a small carnivore: marten (genus Martes). Pine martens (Martes martes L.) can be tolerant to human activities and various landscape features. Small local roads may not impede the genetic connectivity of pine marten in France (Larroque et al. 2016). However, because trapping of pine martens is highly skewed towards roads, with traps generally placed in their close vicinity (Wiebe et al. 2013), mortality risk is higher closer to small roads. Pacific marten (Martes caurina, Merriam) has been shown to select home ranges and display higher level of activity farther away from salvage logging roads than can be explained purely by chance (Volkmann and Hodges 2021), but in the absence of natural predators it has also been shown to select habitats close to forest roads, both active and abandoned, possibly due to higher prey abundance and occasional deer carrion (Breault et al. 2021). In other words, their response towards forest roads seems to be interacting with other variables, such as mortality risk or food availability.
3.3.4 Concluding remarks and research gaps
The effects of forest roads on wildlife have primarily been studied at a species level and less so at a community level. Variables such as species richness have been investigated, whereas the effects on ecological interactions, such as predator-prey and competition dynamics, are much more difficult to assess and have received little attention, not least in Sweden where large carnivores are scarce. Nevertheless, impacts on key species in ecological communities may have cascading effects on other species as well as the functionality of the whole community. Therefore, future studies into the effects of forest roads on wildlife in Sweden should aim to include more than a single species, and preferably more than one higher taxon such as order (for example ungulates or carnivores) or class (for example mammals or birds).
Further, although numerous studies of wildlife species movement patterns in relation to forest roads have observed some level of avoidance or attraction, causal relationships have rarely been shown. In most studies the reported effects seem to be attributed to direct disturbance by roads through habitat fragmentation or disturbance factors related to human activity, but others argue that indirect factors such as forestry operations, especially clearcutting, or altered predation or competition levels between wildlife species, may be the key drivers of the effects observed. In Sweden, wildlife disturbance by human activity is probably low along most forest roads, whereas the impact of forest roads on habitat loss and fragmentation may be much more significant. At the same time, vegetation along road corridors typically includes deciduous species with high forage value for large herbivores. The browsing pressure from large herbivores is generally high in Swedish forest landscapes, particularly with respect to highly selected food items such as some deciduous species. The roadside vegetation may benefit these animals by providing high quality forage. Furthermore, Swedish production forests are typically monocultures of Norway spruce (Picea abies (L.) H. Karst.) or Scots pine (Pinus sylvestris L.), and corridors of deciduous vegetation may provide beneficial habitat heterogeneity for birds and small mammals.
Clearly, forest roads have consequences for wild mammals beyond the construction of the road itself because they enable clearcutting and other human impacts, for instance hunting. This is especially the case in Sweden, where rotational forestry with large clearcut areas is the dominant forest management regime and hunting is the most important cause of mortality for large herbivores as large carnivores are scarce. In addition, both roads and hiking trails may negatively affect the occupancy of wild mammals, even in protected areas such as nature reserves, and, thus, planning of all such infrastructure should take into account how its impacts on wildlife habitat can be minimised (Soultan et al. 2021). One way of doing that is to place roads and trails in the periphery of the protected area rather than allowing them to pass through. In parallel with that, forest roads in Sweden could be planned to circumnavigate, rather than dissect, larger continuous forests to minimise disturbance. Finally, improving existing understanding about the ecological effects of forest roads on wildlife requires an approach that is both holistic (encompassing movement, predation, and food availability) and domestic. Both favourable and adverse effects are likely to be found. Special attention should be paid to changes in species compositions such as atypical species migration into new forest areas and the disappearance of characteristic species.
3.4 Plants
Forest roads can both favour and disfavour plants depending on the species and type of forest road. Apart from the negative effect of road construction on habitat destruction, the impact of forest roads on plants mainly relates to the frequency of road maintenance as well as the width of the roadway and the cut area adjacent to the road. This affects disturbance intensity and how much sunlight that reaches the ground, i.e., sun exposure, which in turn affects plants (Persson 1995; Runesson 2012).
Plant species associated with forest interior are disfavoured by forest roads because they open up the forest (Lázaro-Lobo and Ervin 2019). However, many plants are adapted to intermediate levels of disturbance, i.e., high enough to inhibit dominance by shading trees and bushes, but low enough to allow plant growth and seed spreading. Most studies of how plants are affected by different management types and intensities have focused on larger public roads and concern plant species historically associated with grasslands and pastures (Runesson 2012). This is probably because road verges have taken on a more significant role as potential habitat for plants that are adapted to traditional grassland management, many of which are currently threatened due to the sizeable loss of grasslands and pastures in Sweden since the 1940s (Ihse 1995).
Road verges along smaller roads are favourable biotopes for plants but are not necessarily a substitute for grasslands and pastures. In Sweden and Finland, plant species richness was found to be similar or higher along smaller roads (including forest roads) than in adjacent grasslands and pastures, but had different species composition and plant traits (Tikka et al. 2000; Jantunen et al. 2006; Runesson 2009).
Compared to road verges on larger public roads, forest road verges in Sweden generally provide shadier conditions and less impact from road maintenance such as salt addition and ditch cleaning (Tikka et al. 2000; Runesson 2009; Auestad and Hanssen 2010). However, forest roads also vary in quality as plant habitat in terms of sun exposure and maintenance. Lower maintenance intensity allows for lusher vegetation, which can benefit other threatened plant-dependent organisms such as bees, butterflies, and beetles (Lennartsson and Gylje 2009). The fact that forest roads create a network of open or semi-open areas in the forest landscape can also increase the abundance of deciduous tree species. Tikka et al. (2000) showed that the deciduous tree species goat willow (Salix caprea L.) and grey alder (Alnus incana (L.) Moench) were more common along forest roads in Finland than in interior parts of the forest.
One concern relating to road construction and maintenance is the spread of invasive plant species (Rauschert et al. 2017). Road construction favours the establishment of exotic plants in all kinds of landscapes, including forests (Lázaro-Lobo and Ervin 2019). Exotic plants are unlikely to become established in mature forests since many non-native plants are favoured by open and disturbed conditions (Dodson and Fiedler 2006; Shields and Webster 2007). However, forest roads can act as suitable biotopes for noxious weeds and ease the spread of non-native plant species into newly disturbed forest areas and other more open biotopes (Tikka et al. 2001; Birdsall et al. 2012). The invasive herb lupin (Lupinus polyphyllus Lindl.) is common along roads in Sweden. In a study from Finland, plant diversity and butterfly abundance in road verges along larger public roads were negatively affected by the presence of lupin (Valtonen et al. 2006a).
3.4.1 Concluding remarks and research gaps
Most studies of plants growing on road verges have been performed along larger public roads with intensive maintenance regimes and have focused on plant species historically associated with grasslands and pastures. Hence, it is unclear how most Swedish forest roads, which normally have shadier conditions and less frequent maintenance, affect plants, including their potential to act as valuable biotopes for plant species that favour open or semi-open conditions and a cutting interval of two to five years.
The management of forest road verges could be used as a tool for increasing local and regional plant biodiversity. For example, sections of forest road with the potential to offer habitat to threatened grassland and meadow species could be identified and specifically managed to support these types of plants. There is ample evidence from Nordic countries on how to design road maintenance to benefit such plants: for example, cutting late in the season, removing plant material after cutting, sowing desirable plants and adapting ditch cleaning (Persson 1995; Schaffers 2002; Jantunen et al. 2007; Runesson 2009; Auestad et al. 2010).
3.5 Insects
Insects are a species-rich group of organisms which can be affected by forest roads in several ways, for example increased mortality risk due to vehicle collisions and changes in habitat characteristics and dispersal possibilities. One review of the effect of roads on insects found a strong positive correlation between traffic volume and insect mortality due to vehicle collisions (Muñoz et al. 2015). This suggests that forest roads, which have low traffic intensity, are unlikely to cause significantly increased insect mortality.
Ground-living beetles in Finland and Sweden are affected by forest roads in terms of both species richness and species assemblages (Koivula 2005; Melis et al. 2010). Koivula (2005) showed that along Finnish forest roads, ground beetle (Carabidae) assemblages were mostly affected by factors related to light conditions (road width, proximity of open habitat, and compass direction). Species richness of ground beetles was higher, and the species assemblages were different, in road verges than on sites 25 meters into the boreal forest (Koivula 2005). Similar results were reported by Melis et al. (2010) who showed that the species richness of ground beetles was higher at road sides than in the adjacent young forests (<7 years since logging) and decreased with increasing distance from the road. Melis et al. (2010) also found that road verges both along roads with low- and high traffic intensity possessed a different ground beetle fauna than was found in young forests.
It has frequently been suggested that flower-visiting insects benefit from forest roads. Hanula et al. (2016) showed that in American forests, bees and butterflies generally preferred open forest habitats such as roadside corridors, regardless of forest type, geographic region or methods used to create those habitats. In an international review, Phillips et al. (2020) demonstrated that road verges are often hotspots of flowers and pollinators. In Sweden, Westerfelt et al. (2018) showed that population sizes of a bee species associated with disturbed forests can be better predicted by the roadside flora along forest roads than by the flora found in young forests. Berg et al. (2011) concluded that Swedish road verges have the potential to be favourable biotopes for pollinating insects, at least considering small roads, such as forest roads, where traffic is limited. There are indications that butterflies prefer forest roads to roads in open landscapes since more Lepidoptera species were found in road verges surrounded by forest while verges surrounded by cultivated fields generally contained low numbers of Lepidoptera (Saarinen et al. 2005).
Forest roads can affect the movement patterns of insects since they create an open corridor with a hard road surface. It seems well established that larger paved roads inhibit the movement of ground-living beetles (Muñoz et al. 2015; Koivula and Vermeulen 2005). It is unclear to what extent Swedish forest roads affect insects´ movement patterns. Smaller gravel roads have been shown to act as barriers to ground-living beetles and spiders in Germany and Japan (Mader et al. 1990; Yamada et al. 2010). Koivula (2005) frequently captured ground beetle species associated with mature closed forests in the centers of forest roads, concluding that forest roads are likely not dispersal barriers for forest carabids, at least for the species studied. Further, larger public roads in Sweden have been shown to restrict the movement of flying flower-visiting insects to some degree (Dániel-Ferreira et al. 2022). Forest roads can also act as dispersal corridors and/or sink habitat (i.e., a low-quality habitat which needs the input of individuals to maintain its population) for carabids that do not typically inhabit forest biotopes. Koivula (2005) showed that, at roadsides surrounded by forest, generalist carabids decreased in abundance with increasing distance to the nearest large open-habitat area along the road.
Helldin et al. (2015) used the concept of ‘responsibility species’ for road and railroad verges in Sweden, meaning red-listed species (following IUCN, The International Union for Conservation of Nature, criteria) at least 20% of the occurrence of which is in road- or railroad verges. This level of occurrence (≥20%) was based on information retrieved from “Artportalen” (https://artportalen.se/, which is managed by SLU Swedish Species Information Centre), an internet-based platform for voluntary reporting of species observations. They found 32 such responsibility species for road- and railroad verges, of which 18 were insects. By analysing the habitat and management requirements of these species, the following qualities and structures of road sides were found to benefit the responsibility species: open or semi-open habitats, drier land on sandy or limey soil, solitary trees, patches of bare soil, and sun exposed wood. Conservation actions prescribed for these species include late or irregular mowing, planting of host species, protecting and providing particular substrates, and special protection of certain sites.
3.5.1 Concluding remarks and research gaps
As for plants, most studies of insects have been performed along larger public roads with intense maintenance regimes (Gardiner et al. 2018) which makes assessing the impact of forest roads on insects more difficult. The environment created along many Swedish forest roads offers open or semi-open conditions with less intense maintenance than larger public roads. This allows for more lush vegetation, which benefits many insects, not least flower-visiting insects (Valtonen et al. 2006b; Lennartsson and Gylje 2009). Such conditions are reminiscent of power-line corridors, which have been shown to be ideal biotopes for many butterfly species (Berg et al. 2011).
As often applies to insect research, there is a lack of studies regarding many insect groups. The most studied insect groups in terms of the effect of forest roads, are ground-living beetles, butterflies, and bees. Forest roads affect ground-living beetles by altering species composition, and the same probably applies to other insect groups that have not yet been studied. Flower-visiting insects generally benefit from forest roads, especially if the road verges are rich in flowers, and bees also benefit from roadsides with bare ground (Munguira and Thomas 1992; Saarinen et al. 2005; Hopwood 2008; Helldin et al. 2015).
How the density of forest roads affects the insect fauna at a landscape level is relatively unknown, but there are indications that the local conditions along Swedish forest roads have a greater influence on abundance and species composition than landscape factors (Berg et al. 2011; Dániel-Ferreira et al. 2020).
3.6 Water
3.6.1 Hydrology
Forest roads in Sweden are constructed with the specific aim of influencing local hydrology to safeguard road functionality (Fig. 3) (Kuttah and Arvidsson 2017). Obviously, this affects not just road functionality but local hydrology more broadly. Road ditches (along any type of road) account for 25% of total water channel length in 11 regions across Sweden where the land cover type is predominantly forest (Paul et al. 2022). The remainder are forest ditches (56%), natural channels (13%) and agricultural ditches (6%). According to a review by Kastridis (2020), forest roads have multiple impacts on hydrology: they increase surface runoff when rainfall intensity exceeds road surface infiltration capacity; intercept surface and subsurface water flow from road cut slopes; enhance hydrological connectivity with receiving surface water bodies; and increase the density of the hydrographic network and change the shape of the hydrograph for the catchment. Hydrological impact and/or erosion caused by forest roads is generally more intense during storm events, in steep terrain, and in areas with highly erosive soils, peatlands, and other wet areas (Kastridis 2020). Inadequate road construction can cause increased erosion, landslips, sediment export to surface water bodies, and adverse impacts on local hydrology such as drainage of nearby wetlands (Boston 2016; Lomander et al. 2016). Road construction in wet areas requires special consideration and technical solutions (Skogskunskap 2024 ) and involves higher risk of altering water flows and harming water quality.
Simulations of the hydrological response under different storm intensities for 20 forested catchments in Sweden indicated that road topography had an effect but this was much smaller than anticipated (Nickman et al. 2016). In this study, it was assumed that the road network, including its embankments, was the macro-scale element in the catchment that affected water routing. All types of roads were included, and simulations were performed with and without roads, i.e., the roads were virtually removed from the digital elevation model. Any changes in micro-topography and/or surface infiltration due to road construction were considered to have minimal impact. In another study, hydrological modelling was used to model peak discharge and water level for three drainage structures at road/stream intersections on major general roads in a catchment in southwest Sweden, using various climate and land-use scenarios (Kalantari et al. 2014). The results indicated that the dimensions of these structures were insufficient to manage the highest water depths simulated. For main roads in Sweden, Kalantari and Folkeson (2013) suggest various measures for adapting drainage system construction, operations, and maintenance to climate change based on a survey of professionals working with road drainage. They propose that the capacity of the drainage facilities should be increased, ditch slopes stabilised, and measures should be taken to prevent clogging of culverts. These measures could also be relevant for forest roads.
3.6.2 Water quality and aquatic organisms
Construction and maintenance of forest roads may increase sediment exports to nearby surface water bodies, and this could be detrimental to aquatic life (Croke and Hairsine 2006; Boston 2016). Factors that influence the amount of sediment generated from forest roads include the condition of the road, maintenance practices for roadside ditches, extent of exposed surfaces on the cut and fill slopes, soil texture and climate since this affects the amount, type, and intensity of precipitation events (Boston 2016).
Salt (CaCl2 or MgCl2) or lignosulfonate is applied once or twice per year to Swedish forest roads to reduce dusting and consolidate the road surface (Skogskunskap 2024). De-icing of public roads using salt (NaCl) damages soil, groundwater, and stream water chemistry (Löfgren 2001; Lazur et al. 2020). Likewise, salt addition to forest roads could potentially affect the chemistry of nearby water, but no studies on this topic were found which related to forest roads. Investigations have been undertaken to determine the consequences of using wood ash in the construction of gravel roads. From a study in central Sweden, Nordmark et al. (2022) concluded that mixing 30% wood fly ash into the road base material had low and acceptable impacts on nearby soil and vegetation.
Culverts, which are used to enable forest roads to cross streams, must be correctly installed to avoid adverse effects on aquatic organisms. Specific recommendations are available in Sweden on installing road culverts to reduce soil stability problems on gullies and slopes (Lomander et al. 2016). Diebel et al. (2015) concluded that studies of the effects of road crossings on fish passage (mainly individual crossings, typically culverts, located in small streams) have generally reported lower rates of fish passage through road crossings with outlet drops, high water velocities and shallow water depths than through crossings that better resemble a natural stream reach. In seven small streams in northern Finland/Norway, the effects of restoring impassable road culverts on the distribution of juvenile Atlantic salmon (Salmo salar L.) were evaluated (Erkinaro et al. 2017). Restoration allowed culvert passage of juvenile salmon and increased the distribution area of salmon parr in the streams.
3.6.3 Concluding remarks and research gaps
Although regulations and guidelines relating to forest roads in Sweden do consider hydrology (Lomander et al. 2016; Swedish Forest Agency 2023; Skogskunskap 2024), there are few peer-reviewed articles on the water-related effects of forest roads. The number of physical barriers removed, of which road culverts are one, is used as an indicator for the implementation of the Swedish Environmental Objective “Flourishing Lakes and Streams” (Sveriges miljömål 2024).
The projected changes in climate must be taken into consideration when planning future expansion, maintenance, and use of the forest road network. Road alignment, construction, and maintenance with respect to hydrological conditions may become even more important (Kalantari and Folkeson 2013). Further, the conditions for off-road transportation may worsen (Swedish Meteorological and Hydrological Institute 2024) and forest roads may then become more important for avoiding the soil disturbance and associated sediment export to surface water bodies caused by off-road transportation. In addition, well-built forest roads with properly installed road culverts may provide better stream crossings than temporary bridges, possibly causing lower sediment exports to streams.
Given the limited research into and lack of systematic monitoring of forest road construction, the hydrological and water-quality impacts of the forest road network in Sweden remain rather unclear. More information on the impact of Swedish forest roads on local hydrology and surface water quality would be valuable, both for low-and high-relief terrain (cf. Webster et al. 2015).
4 Discussion
Forest roads in Sweden are built for long-term use and are therefore likely to have long-lasting impacts. This review shows that there is limited and rather fragmented research on the social, ecological, and environmental effects of Swedish forest roads. This supports the findings of Johansson et al. (2013) who concluded that the scientific literature regarding forest roads in Sweden was too limited to enable an evaluation of whether existing environmental considerations, according to legislation and forest certification, were sufficient. Previous research has often focused on how individual factors are affected by roads. Studies covering a broader range of topics related to forest roads are scarce, both in Sweden and internationally. This review shows that the impacts of forest roads are diverse and complex, and all the more so if multiple topics are addressed simultaneously.
When assessing the impacts of forest roads more generally, both consistent and contradictory patterns can be discerned, for example regarding different forms of transport. Forest roads not only enable efficient forestry transportation and increased public access, but they can also act as transport routes for wildlife, reindeer, and water (Pattison and Catterall 2019; Kastridis 2020; Turunen et al. 2020). Conversely, they can be a barrier to the movement of wildlife, ground-dwelling insects, and groundwater flow (Mader et al. 1990; deMaynadier and Hunter Jr 2000; Kastridis 2020). Further, forest roads may both enhance and destroy ecological and social values. For example, forest roads can increase plant and insect diversity in road verges. However, this could entail altering the species composition in an undesirable direction and spreading invasive species (Tikka et al. 2000; Melis et al. 2010; Rauschert et al. 2017; Lázaro-Lobo and Ervin 2019). In addition, forest roads enable recreation and various forest experiences, but improper road delineation can damage or destroy existing habitats, scenic views, and cultural remnants.
Fig. 4 provides a schematic illustration of the potential pathways to ecological, environmental, social, and economic impacts relating to forest roads in Sweden, taking Robinson et al.´s (2010) conceptual framework for addressing the ecological impact of forest roads as a starting point. More research is needed to better understand the connections and interactions between different factors. A comprehensive description of the current status of private roads in Sweden has recently been published by the Swedish Forest Agency, and was produced by a working group consisting of representatives from commercial forestry, forest trade associations, authorities, and research (Bergqvist et al. 2023). The aim was to provide a common basis for identifying measures and initiatives for developing the private road network, particularly the forest road network. The report points out many shortcomings and future challenges.
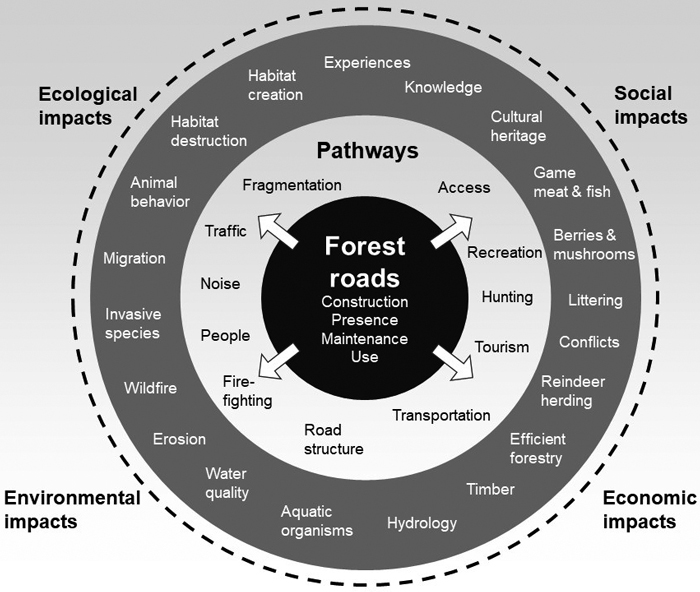
Fig. 4. Schematic illustration of pathways to potential ecological, environmental, social, and economic impacts of forest roads in Sweden. The outer circle (dashed line) represents the interactions between different factors. The illustration focuses on the topics of this study.
In a changing climate with altered precipitation patterns, forest road construction and maintenance procedures may need to be adapted to maintain the functionality of forest roads (Kalantari and Folkeson 2013; Bergqvist et al. 2023). In addition, forest roads may become even more important for firefighting, as the size and number of forest fires in Sweden are expected to increase as a result of climate change (Swedish Portal for Climate Change Adaptation 2024). However, the right of forest owners to block public access to forest roads by installing gates, to avoid problems such as additional maintenance, littering and garbage dumping, could complicate firefighting and rescue operations (Bergqvist et al. 2023). A higher future demand for forest products could lead to more intense forestry and greater need for forestry transportation, increasing the importance of forest roads (Lindén et al. 2023). Recently, uneven-aged and continuous-cover forestry have been proposed as alternatives to even-aged silvicultural management and clearcutting (European Commission 2021). These approaches would probably require more frequent transportation on forest roads and more road maintenance. Finally, technical developments such as electrification of forestry transports and the use of drones could change the future role of forest roads (Pernestål et al. 2023; Buchelt et al. 2024).
In Sweden, the expansion of forest roads only began about 70 years ago, but their spread has been rapid and today they are found in almost all forests. They are taken for granted, and there is no discussion about their existence or non-existence, as has arisen in the USA for example. Indeed, forest roads tend to fall under the radar in the otherwise lively debate about forestry. Many people see it as their right to use the roads to access berry picking sites, recreational areas or as a basis for hiking and cycling. Conflicts arise when different interests coincide, for example when the interests of commercial berry-picking companies, tourism entrepreneurs, reindeer herding, and snowmobiling collide with the interests of the forest owners, when co-ownership and management of forest roads leads to disputes or when increased access to the forest affects threatened species and biodiversity. With the individualisation of society, an increasingly urbanised population, discussions about strengthening private ownership of forests, multiple rights being pitted against each other, and more signs of climate change, the invisible existence of forest roads may come more into the spotlight (Sténs and Mårald 2020).
The forest roads in Sweden have been built primarily to serve the interests of the forest industry in combination with the state’s ambition to strengthen different parts of the country (Mårald et al. 2017). Today, these roads are used for many more purposes than was initially anticipated. As shown in this study, a whole range of social, ecological, and environmental factors are linked to forest roads in Sweden. Finding trade-offs here is not easy. However, by taking a broad perspective and identifying pros and cons, it is possible to understand the contradictions related to forest roads in a holistic way (Fig. 4). Finding solutions to these contradictions and tackling future challenges will require a similarly broad perspective. However, since few studies do this, particularly which focus on forest roads in Fennoscandia, further research is needed.
Acknowledgements
We thank Anna Sténs for contributing to the formulation of the project and participating in the early discussions. Sees-editing Ltd. (https://www.sees-editing.co.uk/) contributed with linguistic refinement of the manuscript.
Funding
The study was funded by Skogssällskapet (grant no. 2019-643) and the Skogforsk framework program 2021-24.
Authors’ contributions
JS conceived the original idea. AD, JS, and ER had the main responsibility for the parts related to forestry. Social values were the main responsibility of EM (sections 3.1.1–3.1.3) and JS (section 3.1.4), reindeer herding was covered by LD, wildlife by MW, plants and insects by PW and water by ER. MW and ER performed the literature search in December 2023. ER led the overall compilation and organisation of the article. All authors contributed to discussions and read and commented several versions of the manuscript.
References
Almstedt Å, Brouder P, Karlsson S, Lundmark L (2014) Beyond post-productivism: from rural policy discourse to rural diversity. Eur Countrys 6: 297–306. https://doi.org/10.2478/euco-2014-0016.
Arksey H, O’Malley L (2005) Scoping studies: towards a methodological framework. Int J Soc Res Method 8: 19–32. https://doi.org/10.1080/1364557032000119616.
Åslund Å (2008) Allemansrätten och marknyttjande: Studier av ett rättsinstitut. [The right of public access and land use: studies of a legal institution]. Doctoral thesis, Department of Management and Engineering, Law and Legal Philosophy, Linköping University. https://urn.kb.se/resolve?urn=urn%3Anbn%3Ase%3Aliu%3Adiva-12082.
Auestad I, Hanssen U (2010) Salt SMART: Endringer i vegkantvegetasjon. [Salt SMART: Changes in roadside vegetation]. Statens vegvesen, Teknologiavdelingen Rapport No. 2583.
Auestad I, Rydgren K, Jongejans E, de Kroon H (2010) Pimpinella saxifraga is maintained in road verges by mosaic management. Biol Conserv 143: 899–907. https://doi.org/10.1016/j.biocon.2009.12.037.
Bartzke GS (2014) Effects of power lines on moose (Alces alces) habitat selection, movements and feeding activity. Doctoral thesis, Faculty of Natural Sciences and Technology, Department of Biology. Norwegian University of Science and Technology. http://hdl.handle.net/11250/245426.
Baxley DL, Qualls CP (2009) Black pine snake (Pituophis melanoleucus lodingi): spatial ecology and associations between habitat use and prey dynamics. J Herpetol 43: 284–293. https://doi.org/10.1670/08-074R1.1.
Baxley D, Lipps GJ, Qualls CP (2011) Multiscale habitat selection by black pine snakes (Pituophis melanoleucus lodingi) in Southern Mississippi. Herpetologica 67: 154–166. https://doi.org/10.1655/HERPETOLOGICA-D-10-00029.1.
Bayne E, Leston L, Mahon CL, Sólymos P, Machtans C, Lankau H, Ball JR, Van Wilgenburg SL, Cumming SG, Fontaine T, Schmiegelow FKA, Song SJ (2016) Boreal bird abundance estimates within different energy sector disturbances vary with point count radius. Condor 118: 376–390. https://doi.org/10.1650/CONDOR-15-126.1.
Benítez-López A, Alkemade R, Verweij PA (2010) The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol Conserv 143: 1307–1316. https://doi.org/10.1016/j.biocon.2010.02.009.
Berg A, Gustafson IE (eds) (2013) Kulturarv i gammelskog. [Cultural heritage in old forest]. Swedish Forest Agency. ISBN 978-91-87535- 03-1.
Berg Å, Pärt T (1994) Abundance of breeding farmland birds on arable and set-aside fields at forest edges. Ecography 17: 147–152. https://doi.org/10.1111/j.1600-0587.1994.tb00087.x.
Berg Å, Ahrné K, Öckinger E, Svensson R, Söderström B (2011) Butterfly distribution and abundance is affected by variation in the Swedish forest-farmland landscape. Biol Conserv 144: 2819–2831. https://doi.org/10.1016/j.biocon.2011.07.035.
Bergqvist M, Danielsson G, Fjeld D, Gunnarsson S, Larsson M, Larsson R, Lindström D, Noreland D, Löfstedt J, Persson J, Pettersson D, Sunesdotter E, Svedin E, Thureson K, Wiborgh T (2023) Nulägesbeskrivning av enskilda vägar med inriktning på skogsbrukets transporter. [Description of the current situation of private roads with a focus on forestry transports]. Fries C, Thureson K (eds) Swedish Forest Agency, Rapport 2023/18.
Bernes C, Lundgren LJ (2009) Use and misuse of nature’s resources: an environmental history of Sweden. Swedish Environmental Protection Agency. ISBN 9789162012755.
Beyer HL, Ung R, Murray DL, Fortin M-J (2013) Functional responses, seasonal variation and thresholds in behavioural responses of moose to road density. J Appl Ecol 50: 286–294. https://doi.org/10.1111/1365-2664.12042.
Birdsall JL, McCaughey W, Runyon JB (2012) Roads impact the distribution of noxious weeds more than restoration treatments in a lodgepole pine forest in Montana, U.S.A. Restor Ecol 20: 517–523. https://doi.org/10.1111/j.1526-100X.2011.00781.x.
Björheden R (2019) Effekter på svenskt skogsbruk av sommaren 2018. [The effects on Swedish forestry of the summer 2018]. Skogforsk, Arbetsrapport 1012-2019.
Boan JJ, Malcolm JR, McLaren BE (2014) Forest overstorey and age as habitat? Detecting the indirect and direct effects of predators in defining habitat in a harvested boreal landscape. Forest Ecol Manag 326: 101–108. https://doi.org/10.1016/j.foreco.2014.03.052.
Bojarska K, Maugeri L, Kuehn R, Król W, Theuerkauf J, Okarma H, Gula R (2021) Wolves under cover: the importance of human-related factors in resting site selection in a commercial forest. Forest Ecol Manag 497, article id 119511. https://doi.org/10.1016/j.foreco.2021.119511.
Bolibok L, Brzeziecki B, Drozdowski S, Zawadzka D, Zawadzki J (2014) Application of classification trees for assessment species habitat preferences on the example of capercaillie (Tetrao urogallus). Sylwan 158: 267–276.
Boston K (2016) The potential effects of forest roads on the environment and mitigating their impacts. Current Forestry Reports 2: 215–222. https://doi.org/10.1007/s40725-016-0044-x.
Breault DN, Johnson CJ, Todd M, Gillingham MP (2021) Resource use by an apex mesocarnivore, Pacific marten, in a highly modified forested island ecosystem. Forest Ecol Manag 492, article id 119167. https://doi.org/10.1016/j.foreco.2021.119167.
Buchelt A, Adrowitzer A, Kieseberg P, Gollob C, Nothdurft A, Eresheim S, Tschiatschek S, Stampfer K, Holzinger A (2024) Exploring artificial intelligence for applications of drones in forest ecology and management. Forest Ecol Manag 551, article id 121530. https://doi.org/10.1016/j.foreco.2023.121530.
Chmielewski S (2016) A high number of dead grass snakes Natrix natrix on a local road. KULON 21: 89–92.
Coffin AW (2007) From roadkill to road ecology: a review of the ecological effects of roads. J Transp Geogr 15: 396–406. https://doi.org/10.1016/j.jtrangeo.2006.11.006.
Courtois R, Gingras A, Fortin D, Sebbane A, Rochette B, Breton L (2008) Demographic and behavioural response of woodland caribou to forest harvesting. Can J Forest Res 38: 2837–2849. https://doi.org/10.1139/x08-119.
Croke JC, Hairsine PB (2006) Sediment delivery in managed forests: a review. Environ Rev 14: 59–87. https://doi.org/10.1139/a05-016.
Dániel-Ferreira J, Bommarco R, Wissman J, Öckinger E (2020) Linear infrastructure habitats increase landscape-scale diversity of plants but not of flower-visiting insects. Sci Rep 10, article id 21374. https://doi.org/10.1038/s41598-020-78090-y.
Dániel-Ferreira J, Berggren Å, Wissman J, Öckinger E (2022) Road verges are corridors and roads barriers for the movement of flower-visiting insects. Ecography 2022, article id e05847. https://doi.org/10.1111/ecog.05847.
Darlington S, Ladle A, Burton AC, Volpe JP, Fisher JT (2022) Cumulative effects of human footprint, natural features and predation risk best predict seasonal resource selection by white-tailed deer. Sci Rep 12, article id 1072. https://doi.org/10.1038/s41598-022-05018-z.
Davidsson A, Gustavsson O, Parklund T (2023) Skogsbrukets vägtransporter 2020 – En nulägesbeskrivning av flöden av biomassa från skog till industri [Road transports in forestry 2020 – a description of the current situation of biomass flows from forest to industry]. Skogforsk, Arbetsrapport 1142-2023.
Deguise I, Richardson JS (2009) Movement behaviour of adult western toads in a fragmented, forest landscape. Can J Zool 87: 1184–1194. https://doi.org/10.1139/Z09-109.
DeMars CA, Boutin S (2018) Nowhere to hide: effects of linear features on predator–prey dynamics in a large mammal system. J Anim Ecol 87: 274–284. https://doi.org/10.1111/1365-2656.12760.
deMaynadier PG, Hunter Jr. ML (1995) The relationship between forest management and amphibian ecology: a review of the North American literature. Environ Rev 3: 230–261. https://doi.org/10.1139/a95-012.
deMaynadier PG, Hunter Jr ML (2000) Road effects on amphibian movements in a forested landscape. Nat Area J 20: 56–65.
Dibner R, Lawton C, Marnell F (2014) Reproduction of common frogs, Rana temporaria, in a managed conifer forest and bog landscape in Western Ireland. Herpetol Conserv Bio 9: 38–47.
Diebel MW, Fedora M, Cogswell S, O’Hanley JR (2015) Effects of road crossings on habitat connectivity for stream-resident fish. River Res Appl 31: 1251–1261. https://doi.org/10.1002/rra.2822.
Dobor L, Hlásny T, Rammer W, Zimová S, Barka I, Seidl R (2020) Spatial configuration matters when removing windfelled trees to manage bark beetle disturbances in Central European forest landscapes. J Environ Manage 254, article id 109792. https://doi.org/10.1016/j.jenvman.2019.109792.
Dodson EK, Fiedler CE (2006) Impacts of restoration treatments on alien plant invasion in Pinus ponderosa forests, Montana, USA. J Appl Ecol 43: 887–897. https://doi.org/10.1111/j.1365-2664.2006.01206.x.
Eliasson P (2011) The state-owned forests: silviculture, mechanisation and institutional change. In: Jansson U, Antonson H (eds) Agriculture and forestry in Sweden since 1900: geographical and historical studies, pp 390–405. The Royal Swedish Academy of Agriculture and Forestry. ISBN 978-91-86573-11-9.
Elzanowski A, Ciesiołkiewicz J, Kaczor M, Radwańska J, Urban R (2009) Amphibian road mortality in Europe: a meta-analysis with new data from Poland. Eur J Wildlife Res 55: 33–43. https://doi.org/10.1007/s10344-008-0211-x.
Eriksen A, Wabakken P, Zimmermann B, Andreassen HP, Arnemo JM, Gundersen H, Milner JM, Liberg O, Linnell J, Pedersen HC, Sand H, Solberg EJ, Storaas T (2009) Encounter frequencies between GPS-collared wolves (Canis lupus) and moose (Alces alces) in a Scandinavian wolf territory. Ecol Res 24: 547–557. https://doi.org/10.1007/s11284-008-0525-x.
Erkinaro J, Erkinaro H, Niemelä E (2017) Road culvert restoration expands the habitat connectivity and production area of juvenile Atlantic salmon in a large subarctic river system. Fisheries Manag Ecol 24: 73–81. https://doi.org/10.1111/fme.12203.
European Commission (2021) New EU forest strategy for 2030. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions, COM(2021) 572 Final. European Commission: Brussels, Belgium.
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an empirical review and synthesis. Ecol Soc 14, article id 21. https://doi.org/10.5751/ES-02815-140121.
Filipsson S (2001) Skogsbilvägar. [Forest roads]. Swedish Forest Agency, Rapport 81.
Findlay CS, Houlahan J (1997) Anthropogenic correlates of species richness in southeastern Ontario wetlands. Conserv Biol 11: 1000–1009. https://doi.org/10.1046/j.1523-1739.1997.96144.x.
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29: 207–231. https://doi.org/10.1146/annurev.ecolsys.29.1.207.
Francis AL, Procter C, Kuzyk G, Fisher JT (2021) Female moose prioritize forage over mortality risk in harvested landscapes. J Wildlife Manage 85: 156–168. https://doi.org/10.1002/jwmg.21963.
Gardiner MM, Riley CB, Bommarco R, Öckinger E (2018) Rights-of-way: a potential conservation resource. Front Ecol Environ 16: 149–158. https://doi.org/10.1002/fee.1778.
Gicquel M, Sand H, Månsson J, Wallgren M, Wikenros C (2020) Does recolonization of wolves affect moose browsing damage on young Scots pine? Forest Ecol Manag 473, article id 118298. https://doi.org/10.1016/j.foreco.2020.118298.
Gucinski H, Furniss MJ, Ziemer RR, Brookes MH (eds) (2001) Forest roads: a synthesis of scientific information. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, General Technical Report PNW-GTR-509.
Gundersen V, Vistad OI (2016) Public opinions and use of various types of recreational infrastructure in boreal forest settings. Forests 7, article id 113. https://doi.org/10.3390/f7060113.
Gundersen V, Clarke N, Dramstad W, Fjellstad W (2016) Effects of bioenergy extraction on visual preferences in boreal forests: a review of surveys from Finland, Sweden and Norway. Scand J Forest Res 31: 323–334. https://doi.org/10.1080/02827581.2015.1099725.
Gundersen VS, Frivold LH (2008) Public preferences for forest structures: a review of quantitative surveys from Finland, Norway and Sweden. Urban For Urban Gree 7: 241–258. https://doi.org/10.1016/j.ufug.2008.05.001.
Haddad NM, Bowne DR, Cunningham A, Danielson BJ, Levey DJ, Sargent S, Spira T (2003) Corridor use by diverse taxa. Ecology 84: 609–615. https://doi.org/10.1890/0012-9658(2003)084[0609:CUBDT]2.0.CO;2.
Hall CM, Müller DKE (eds) (2004) Tourism, mobility and second homes: between elite landscape and common ground. Channel View Publications, Blue Ridge Summit, Bristol. https://doi.org/10.21832/9781873150825.
Hallikainen V (1998) The Finnish wilderness experience. Finnish Forest Research Institute Research Papers 711. http://urn.fi/URN:ISBN:951-40-1656-4.
Hannerz M, Lindhagen A, Forsberg O, Fries C, Rydberg D (2016) Skogsskötsel för friluftsliv och rekreation. [Silviculture for outdoor activities and recreation]. Swedish Forest Agency, Skogsskötselserien no. 15.
Hanula JL, Ulyshen MD, Horn S (2016) Conserving pollinators in North American forests: a review. Nat Area J 36: 427–439. https://doi.org/10.3375/043.036.0409.
Helldin J-O, Wissman J, Lennartsson T (2015) Abundance of red-listed species in infrastructure habitats – ”responsibility species” as a priority-setting tool for transportation agencies´ conservation action. Nat Conserv 11: 143–158. https://doi.org/10.3897/natureconservation.11.4433.
Hellström E (2001) Conflict cultures – qualitative comparative analysis of environmental conflicts in forestry. Silva Fenn Monogr 2. https://doi.org/10.14214/sf.sfm2.
Holgén P, Mattsson L, Li CZ (2000) Recreation values of boreal forest stand types and landscapes resulting from different silvicultural systems: an economic analysis. J Environ Manage 60: 173–180. https://doi.org/10.1006/jema.2000.0377.
Homyack JA, O’Bryan CJ, Thornton JE, Baldwin RF (2016) Community occupancy of herpetofauna in roadside ditches in a managed pine landscape. Forest Ecol Manag 361: 346–357. https://doi.org/10.1016/j.foreco.2015.11.037.
Hopwood JL (2008) The contribution of roadside grassland restorations to native bee conservation. Biol Conserv 141: 2632–2640. https://doi.org/10.1016/j.biocon.2008.07.026.
Hörnsten L, Fredman P (2000) On the distance to recreational forests in Sweden. Landscape Urban Plan 51: 1–10. https://doi.org/10.1016/S0169-2046(00)00097-9.
Horstkotte T (2013) Contested landscapes: social-ecological interactions between forestry and reindeer husbandry. Doctoral thesis, Department of Ecologyand Environmental Science, Umeå University. https://urn.kb.se/resolve?urn=urn%3Anbn%3Ase%3Aumu%3Adiva-66386.
Hummel S, Meyer L, Hackländer K, Weber D (2017) Activity of potential predators of European hare (Lepus europaeus) leverets and ground-nesting birds in wildflower strips. Eur J Wildlife Res 63, article id 102. https://doi.org/10.1007/s10344-017-1158-6.
Ihse M (1995) Swedish agricultural landscapes – patterns and changes during the last 50 years, studied by aerial photos. Landscape Urban Plan 31: 21–37. https://doi.org/10.1016/0169-2046(94)01033-5.
Ikonen VP, Kilpeläinen A, Zubizarreta-Gerendiain A, Strandman H, Asikainen A, Venäläinen A, Kaurola J, Kangas J, Peltola H (2017) Regional risks of wind damage in boreal forests under changing management and climate projections. Can J Forest Res 47: 1632–1645. https://doi.org/10.1139/cjfr-2017-0183.
Jantunen J, Saarinen K, Valtonen A, Saarnio S (2006) Grassland vegetation along roads differing in size and traffic density. Ann Bot Fenn 43: 107–117.
Jantunen J, Saarinen K, Valtonen A, Saarnio S (2007) Flowering and seed production success along roads with different mowing regimes. Appl Veg Sci 10: 285–292. https://doi.org/10.1111/j.1654-109X.2007.tb00528.x.
Johansson T, Hjältén J, de Jong J, von Stedingk H (2013) Environmental considerations from legislation and certification in managed forest stands: a review of their importance for biodiversity. Forest Ecol Manag 303: 98–112. https://doi.org/10.1016/j.foreco.2013.04.012.
Johnson BA, Homyack JA, Barrett K, Baldwin RF (2016) Factors influencing herpetofaunal assemblages of aquatic systems in a managed pine forest. Forest Ecol Manag 379: 124–132. https://doi.org/10.1016/j.foreco.2016.08.012.
Kalantari Z, Folkeson L (2013) Road drainage in Sweden: current practice and suggestions for adaptation to climate change. J Infrastruct Syst 19: 147–156. https://doi.org/10.1061/(ASCE)IS.1943-555X.0000119.
Kalantari Z, Briel A, Lyon SW, Olofsson B, Folkeson L (2014) On the utilization of hydrological modelling for road drainage design under climate and land use change. Sci Total Environ 475: 97–103. https://doi.org/10.1016/j.scitotenv.2013.12.114.
Kämmerle J-L, Taubmann J, Andrén H, Fiedler W, Coppes J (2021) Environmental and seasonal correlates of capercaillie movement traits in a Swedish wind farm. Ecol Evol 11: 11762–11773. https://doi.org/10.1002/ece3.7922.
Kangas K, Luoto M, Ihantola A, Tomppo E, Siikamäki P (2010) Recreation-induced changes in boreal bird communities in protected areas. Ecol Appl 20: 1775–1786. https://doi.org/10.1890/09-0399.1.
Kastridis A (2020) Impact of forest roads on hydrological processes. Forests 11, article id 1201. https://doi.org/10.3390/f11111201.
Kivinen S (2015) Many a little makes a mickle: cumulative land cover changes and traditional land use in the Kyrö reindeer herding district, northern Finland. Appl Geogr 63: 204–211. https://doi.org/10.1016/j.apgeog.2015.06.013.
Koivula MJ (2005) Effects of forest roads on spatial distribution of boreal carabid beetles (Coleoptera: Carabidae). Coleopts Bull 59: 465–487. https://doi.org/10.1649/815.1.
Koivula MJ, Vermeulen HJW (2005) Highways and forest fragmentation – effects on carabid beetles (Coleoptera, Carabidae). Landscape Ecol 20: 911–926. https://doi.org/10.1007/s10980-005-7301-x.
Kolström M, Lindner M, Vilén T, Maroschek M, Seidl R, Lexer MJ, Netherer S, Kremer A, Delzon S, Barbati A, Marchetti M, Corona P (2011) Reviewing the science and implementation of climate change adaptation measures in European forestry. Forests 2: 961–982. https://doi.org/10.3390/f2040961.
Kuklina V, Petrov AN, Krasnoshtanova N, Bogdanov V (2020) Mobilizing benefit-sharing through transportation infrastructure: informal roads, extractive industries and benefit-sharing in the Irkutsk oil and gas region, Russia. Resources 9, article id 21. https://doi.org/10.3390/resources9030021.
Kuttah D, Arvidsson H (2017) Effect of groundwater table rising on the performance of a Swedish-designed gravel road. Transp Geotech 11: 82–96. https://doi.org/10.1016/j.trgeo.2017.05.001.
Larroque J, Ruette S, Vandel J-M, Devillard S (2016) Divergent landscape effects on genetic differentiation in two populations of the European pine marten (Martes martes). Landscape Ecol 31: 517–531. https://doi.org/10.1007/s10980-015-0281-6.
Larson CD, Rew LJ (2022) Restoration intensity shapes floristic recovery after forest road decommissioning. J Environ Manage 319, article id 115729. https://doi.org/10.1016/j.jenvman.2022.115729.
Laschi A, Foderi C, Fabiano F, Neri F, Cambi M, Mariotti B, Marchi E (2019) Forest road planning, construction and maintenance to improve forest firefighting: a review. Croat J For Eng 40: 207–219.
Lázaro-Lobo A, Ervin GN (2019) A global examination on the differential impacts of roadsides on native vs. exotic and weedy plant species. Global Ecol Conserv 17, article id e00555. https://doi.org/10.1016/j.gecco.2019.e00555.
Lazur A, VanDerwerker T, Koepenick K (2020) Review of implications of road salt use on groundwater quality – corrosivity and mobilization of heavy metals and radionuclides. Water Air Soil Poll 231, article id 474. https://doi.org/10.1007/s11270-020-04843-0.
Lennartsson T, Gylje S (2009) Infrastrukturens biotoper – en refug för biologisk mångfald. [Biotopes of infrastructure – a refuge for biodiversity]. SLU Swedish Biodiversity Centre, CBM:s skriftserie 31.
Lind T, Oraug J, Rosenfeld IS, Østensen E (1974) Friluftsliv i Oslomarka. Analyse av en intervjuundersøkelse om publikums bruk og krav til Oslomarka. [Recreation in Oslomarka. Analysis of an interview survey about people’s use of and demands on Oslomarka]. Norwegian Institute for Urban and Regional Research, Oslo, Arbeidsrapport 8/74.
Lindén H, Mörling T, Nilsson C, Paulsson J, Wester J (2023) Förslag till ett nationellt mål för ökad hållbar tillväxt i skogen. [Proposal for a national objective for increased and sustainable growth in forests]. Swedish Forest Agency, Rapport 2023/21.
Lindhagen A, Hörnsten L (2000) Forest recreation in 1977 and 1997 in Sweden: changes in public preferences and behaviour. Forestry 73: 143–153. https://doi.org/10.1093/forestry/73.2.143.
Lisberg Jensen E, Ouis P (2014) Det gröna finrummet: etnicitet, friluftsliv och naturumgängets urbanisering. [The green living room: ethnicity, outdoor life and the urbanisation of nature]. Carlssons, Stockholm. ISBN 9789173316385.
Löfgren S (2001) The chemical effects of deicing salt on soil and stream water of five catchments in Southeast Sweden. Water Air Soil Poll 130: 863–868. https://doi.org/10.1023/A:1013895215558.
Lomander A, Lundström K, Hazell P (2016) Möjligheter att minska stabilitetsrisker i raviner och slänter vid skogsbruk och exploatering – exempelsamling över anpassningar av åtgärder för att förhindra erosion och stabilitetsproblem i slänter i samband med skogsbruk och exploatering. [Possibilities to reduce stability risks in gullies and slopes in forestry and exploitation – examples of adaptations of measures to prevent erosion and slope stability problems in forestry and exploitation]. Swedish Forest Agency, Rapport 9.
Loosen AE, Devineau O, Zimmermann B, Cromsigt JPGM, Pfeffer SE, Skarpe C, Marie Mathisen K (2021) Roads, forestry, and wolves interact to drive moose browsing behavior in Scandinavia. Ecosphere 12, article id e03358. https://doi.org/10.1002/ecs2.3358.
Lugo AE, Gucinski H (2000) Function, effects, and management of forest roads. Forest Ecol Manag 133: 249–262. https://doi.org/10.1016/S0378-1127(99)00237-6.
Lundmark T, Bergh J, Hofer P, Lundström A, Nordin A, Poudel BC, Sathre R, Taverna R, Werner F (2014) Potential roles of Swedish forestry in the context of climate change mitigation. Forests 5: 557–578. https://doi.org/10.3390/f5040557.
Mackenzie JMD (1946) Some factors influencing woodland birds. Q J Forest 40: 82–88.
Mader HJ, Schell C, Kornacker P (1990) Linear barriers to arthropod movements in the landscape. Biol Conserv 54: 209–222. https://doi.org/10.1016/0006-3207(90)90052-Q.
Mårald E, Sandström C, Nordin A, Beland Lindahl K, Roberge J-M, Johansson J, Rist L (2017) Forest governance and management across time: developing a new forest social contract. Routledge. https://doi.org/10.4324/9781315696430.
Matysek M, Gwiazda R, Figarski T, Zięba F, Klimecki M, Mateja R, Krzan P, Piątek G (2021) What habitat parameters are important for the survival of ground nests in mountain forests? Recommendations for protection of Hazel Grouse Tetrastes bonasia based on an experiment with artificial nests. Bird Study 68: 258–266. https://doi.org/10.1080/00063657.2022.2026875.
McGregor RL, Bender DJ, Fahrig L (2008) Do small mammals avoid roads because of the traffic? J Appl Ecol 45: 117–123. https://doi.org/10.1111/j.1365-2664.2007.01403.x.
Meier NL (2013) Sustainable outdoor recreation and the Forest Service: agency culture, response to change and implications for the future. Thesis, Utah State University, All Graduate Plan B and other Reports 338. https://doi.org/10.26076/be7e-5588.
Melis C, Olsen CB, Hyllvang M, Gobbi M, Stokke BG, Røskaft E (2010) The effect of traffic intensity on ground beetle (Coleoptera: Carabidae) assemblages in central Sweden. J Insect Conserv 14: 159–168. https://doi.org/10.1007/s10841-009-9240-3.
Munguira ML, Thomas JA (1992) Use of road verges by butterfly and burnet populations, and the effect of roads on adult dispersal and mortality. J Appl Ecol 29: 316–329. https://doi.org/10.2307/2404501.
Muñoz PT, Torres FP, Megías AG (2015) Effects of roads on insects: a review. Biodivers Conserv 24: 659–682. https://doi.org/10.1007/s10531-014-0831-2.
Narayanaraj G, Wimberly MC (2012) Influences of forest roads on the spatial patterns of human- and lightning-caused wildfire ignitions. Appl Geogr 32: 878–888. https://doi.org/10.1016/j.apgeog.2011.09.004.
Nellemann C, Vistnes I, Jordhøy P, Strand O, Newton A (2003) Progressive impact of piecemeal infrastructure development on wild reindeer. Biol Conserv 113: 307–317. https://doi.org/10.1016/S0006-3207(03)00048-X.
Neumann W, Ericsson G, Dettki H, Radeloff VC (2013) Behavioural response to infrastructure of wildlife adapted to natural disturbances. Landscape Urban Plan 114: 9–27. https://doi.org/10.1016/j.landurbplan.2013.02.002.
Nickman A, Lyon SW, Jansson P-E, Olofsson B (2016) Simulating the impact of roads on hydrological responses: examples from Swedish terrain. Hydrol Res 47: 767–781. https://doi.org/10.2166/nh.2016.030.
Nikula A, Nivala V, Matala J, Heliövaara K (2019) Modelling the effect of habitat composition and roads on the occurrence and number of moose damage at multiple scales. Silva Fenn 53, article id 9918. https://doi.org/10.14214/sf.9918.
Nordmark D, Vestin J, Hansson L, Kumpiene J (2022) Long-term evaluation of geotechnical and environmental properties of ash-stabilised road. J Environ Manage 318, article id 115504. https://doi.org/10.1016/j.jenvman.2022.115504.
Oliveira T, Benson JF, Thompson C, Patterson BR (2020) Resource selection at homesites by wolves and eastern coyotes in a Canis hybrid zone. Ecosphere 11, article id e03320. https://doi.org/10.1002/ecs2.3320.
Ou T (2017) Droughts and wildfires in Sweden, past variation and future projection. Swedish Civil Contingencies Agency, Report MSB1112.
Patthey P, Signorell N, Rotelli L, Arlettaz R (2012) Vegetation structural and compositional heterogeneity as a key feature in Alpine black grouse microhabitat selection: conservation management implications. Eur J Wildlife Res 58: 59–70. https://doi.org/10.1007/s10344-011-0540-z.
Pattison CA, Catterall CP (2019) Effects of narrow linear clearings on movement and habitat use in a boreal forest mammal community during winter. PeerJ 7, article id e6504. https://doi.org/10.7717/peerj.6504.
Paul SS, Hasselquist EM, Jarefjäll A, Ågren AM (2022) Virtual landscape-scale restoration of altered channels helps us understand the extent of impacts to guide future ecosystem management. Ambio 52: 182–194. https://doi.org/10.1007/s13280-022-01770-8.
Pernestål A, Svenson G, Gustavsson O, Eriksson A (2023) Elektrifiering av skogsbrukets vägtransporter – Analys av barriärer och möjligheter. [Electrification of forestry road transports – analysis of barriers and opportunities]. Skogforsk, Arbetsrapport 1176–2023.
Persson TS (1995) Management of roadside verges: vegetation changes and species diversity. Thesis, Department of Ecology and Environmental Research, Swedish University of Agricultural Sciences.
Phillips BB, Wallace C, Roberts BR, Whitehouse AT, Gaston KJ, Bullock JM, Dicks LV, Osborne JL (2020) Enhancing road verges to aid pollinator conservation: a review. Biol Conserv 250, article id 108687. https://doi.org/10.1016/j.biocon.2020.108687.
Pinard V, Dussault C, Ouellet J-P, Fortin D, Courtois R (2012) Calving rate, calf survival rate, and habitat selection of forest-dwelling caribou in a highly managed landscape. J Wildlife Manage 76: 189–199. https://doi.org/10.1002/jwmg.217.
Pinto GASJ, Rousseu F, Niklasson M, Drobyshev I (2020) Effects of human-related and biotic landscape features on the occurrence and size of modern forest fires in Sweden. Agr Forest Meteorol 291, article id 108084. https://doi.org/10.1016/j.agrformet.2020.108084.
Pureswaran DS, Roques A, Battisti A (2018) Forest insects and climate change. Curr For Rep 4: 35–50. https://doi.org/10.1007/s40725-018-0075-6.
Rauschert ESJ, Mortensen DA, Bloser SM (2017) Human-mediated dispersal via rural road maintenance can move invasive propagules. Biol Invasions 19: 2047–2058. https://doi.org/10.1007/s10530-017-1416-2.
Robinson C, Duinker PN, Beazley KF (2010) A conceptual framework for understanding, assessing, and mitigating ecological effects of forest roads. Environ Rev 18: 61–86. https://doi.org/10.1139/A10-002.
Runesson K (2009) Grassland plant species on road verges in Mid Sweden – influence of semi-natural grasslands and impact of road maintenance. Examensarbete 2009:4. Department of Ecology, Swedish University of Agricultural Sciences.
Runesson K (2012) Vegetation och flora i vägkanter – effekter av olika metoder för skötsel och underhåll [Roadside vegetation and flora – effects of different management and maintenance practices]. SLU Swedish Biodiversity Centre, CBM:s skriftserie 63.
Saarinen K, Valtonen A, Jantunen J, Saarnio S (2005) Butterflies and diurnal moths along road verges: does road type affect diversity and abundance? Biol Conserv 123: 403–412. https://doi.org/10.1016/j.biocon.2004.12.012.
Šálek M, Svobodová J, Zasadil P (2010) Edge effect of low-traffic forest roads on bird communities in secondary production forests in central Europe. Landscape Ecol 25: 1113–1124. https://doi.org/10.1007/s10980-010-9487-9.
Sandström C, Lindkvist A, Öhman K, Nordström E-M (2011) Governing competing demands for forest resources in Sweden. Forests 2: 218–242. https://doi.org/10.3390/f2010218.
Sandström P, Granqvist Pahlén T, Edenius L, Tømmervik H, Hagner O, Hemberg L, Olsson H, Baer K, Stenlund T, Brandt LG, Egberth M (2003) Conflict resolution by participatory management: remote sensing and GIS as tools for communicating land-use needs for reindeer herding in Northern Sweden. Ambio 32: 557–567. https://doi.org/10.1579/0044-7447-32.8.557.
Schaffers AP (2002) Soil, biomass, and management of semi-natural vegetation – part II. Factors controlling species diversity. Plant Ecol 158: 247–268. https://doi.org/10.1023/A:1015545821845.
Semlitsch RD, Ryan TJ, Hamed K, Chatfield M, Drehman B, Pekarek N, Spath M, Watland A (2007) Salamander abundance along road edges and within abandoned logging roads in Appalachian forests. Conserv Biol 21: 159–167. https://doi.org/10.1111/j.1523-1739.2006.00571.x.
Shields JM, Webster CR (2007) Ground-layer response to group selection with legacy-tree retention in a managed northern hardwood forest. Can J Forest Res 37: 1797–1807. https://doi.org/10.1139/X07-052.
Sjöström J, Granström A (2023) Human activity and demographics drive the fire regime in a highly developed European boreal region. Fire Safety J 136, article id 103743. https://doi.org/10.1016/j.firesaf.2023.103743.
Skarin A, Åhman B (2014) Do human activity and infrastructure disturb domesticated reindeer? The need for the reindeer’s perspective. Polar Biol 37: 1041–1054. https://doi.org/10.1007/s00300-014-1499-5.
Skogsbrukets motortransportkommitté (1957) Skogsbrukets biltransporter. [Car transport in the forestry sector]. Stockholm.
Skogskunskap (2024) Vägar i skogen. [Roads in the forest]. https://www.skogskunskap.se/vagar-i-skogen/. Accessed 15 August 2024.
SLU Swedish Species Information Centre (2024) Artfakta. [Species information]. https://artfakta.se/. Accessed 2 January 2024.
Snyder H (2019) Literature review as a research methodology: an overview and guidelines. J Bus Res 104: 333–339. https://doi.org/10.1016/j.jbusres.2019.07.039.
Soultan A, Attum O, Lahue W (2021) The relationship between landscape features and domestic species on the occupancy of native mammals in urban forests. Urban Ecosyst 24: 1117–1128. https://doi.org/10.1007/s11252-021-01100-y.
Sténs A, Mårald E (2020) “Forest property rights under attack”: actors, networks and claims about forest ownership in the Swedish press 2014–2017. Forest Policy Econ 111, article id 102038. https://doi.org/10.1016/j.forpol.2019.102038.
Sténs A, Sandström C (2013) Divergent interests and ideas around property rights: the case of berry harvesting in Sweden. Forest Policy Econ 33: 56–62. https://doi.org/10.1016/j.forpol.2012.05.004.
Sténs A, Bjärstig T, Nordström E-M, Sandström C, Fries C, Johansson J (2016) In the eye of the stakeholder: the challenges of governing social forest values. Ambio 45: 87–99. https://doi.org/10.1007/s13280-015-0745-6.
Steyaert SMJG, Støen O-G, Elfström M, Karlsson J, Lammeren RV, Bokdam J, Zedrosser A, Brunberg S, Swenson JE (2011) Resource selection by sympatric free-ranging dairy cattle and brown bears Ursus arctos. Wildlife Biol 17: 389–403. https://doi.org/10.2981/11-004.
Sveriges miljömål (2024) Levande sjöar och vattendrag. [Flourishing lakes and streams]. https://www.sverigesmiljomal.se/miljomalen/levande-sjoar-och-vattendrag/. Accessed 27 May 2024.
Swedish Environmental Protection Agency (2024) Vanliga frågor om jakt. [Frequently asked questions on hunting]. https://www.naturvardsverket.se/amnesomraden/jakt-och-vilt/vanliga-fragor-om-jakt/. Accessed 27 February 2024.
Swedish Forest Agency (2023) Skogsvårdslagstiftningen, Gällande regler 1 september 2022. [Forestry legislation, Rules in force 1 September 2022].
Swedish Meteorological and Hydrological Institute (2024) Basic climate change scenario service. https://www.smhi.se/en/climate/future-climate/basic-climate-change-scenario-service/sverige/medeltemperatur/rcp45/2071-2100. Accessed 27 May 2024.
Swedish National Heritage Board (2010) Vägledning för tillämpningen av Kulturminneslagen: samråd och tillståndsprövning i samband med skogs- och jordbruk m.m. [Guidance on the application of the Cultural Heritage Act: consultation and authorisation in connection with forestry, agriculture, etc.].
Swedish Portal for Climate Change Adaptation (2024) Forest fire. https://www.klimatanpassning.se/en/climate-change-in-sweden/climate-effects/forest-fire-1.96642. Accessed 2 January 2024).
Thompson MP, Gannon BM, Caggiano MD (2021) Forest roads and operational wildfire response planning. Forests 12, article id 110. https://doi.org/10.3390/f12020110.
Thorsen NH, Hansen JE, Støen OG, Kindberg J, Zedrosser A, Frank SC (2022) Movement and habitat selection of a large carnivore in response to human infrastructure differs by life stage. Mov Ecol 10, article id 52. https://doi.org/10.1186/s40462-022-00349-y.
Tikka PM, Koski PS, Kivelä RA, Kuitunen MT (2000) Can grassland plant communities be preserved on road and railway verges? Appl Veg Sci 3: 25–32. https://doi.org/10.2307/1478915.
Tikka PM, Högmander H, Koski PS (2001) Road and railway verges serve as dispersal corridors for grassland plants. Landscape Ecol 16: 659–666. https://doi.org/10.1023/A:1013120529382.
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14: 18–30. https://doi.org/10.1046/j.1523-1739.2000.99084.x.
Turunen MT, Rasmus S, Järvenpää J, Kivinen S (2020) Relations between forestry and reindeer husbandry in northern Finland – perspectives of science and practice. Forest Ecol Manag 457, article id 117677. https://doi.org/10.1016/j.foreco.2019.117677.
Valtonen A, Jantunen J, Saarinen K (2006a) Flora and lepidoptera fauna adversely affected by invasive Lupinus polyphyllus along road verges. Biol Conserv 133: 389–396. https://doi.org/10.1016/j.biocon.2006.06.015.
Valtonen A, Saarinen K, Jantunen J (2006b) Effect of different mowing regimes on butterflies and diurnal moths on road verges. Anim Biodiv Conserv 29: 133–148. https://doi.org/10.32800/abc.2006.29.0133.
Vanlandeghem V, Drapeau P, Prima M-C, St-Laurent M-H, Fortin D (2021) Management-mediated predation rate in the caribou–moose–wolf system: spatial configuration of logging activities matters. Ecosphere 12, article id e03550. https://doi.org/10.1002/ecs2.3550.
Venäläinen A, Lehtonen I, Laapas M, Ruosteenoja K, Tikkanen O-P, Viiri H, Ikonen V-P, Peltola H (2020) Climate change induces multiple risks to boreal forests and forestry in Finland: a literature review. Global Change Biol 26: 4178–4196. https://doi.org/10.1111/gcb.15183.
Vistnes I, Nellemann C (2008) The matter of spatial and temporal scales: a review of reindeer and caribou response to human activity. Polar Biol 31: 399–407. https://doi.org/10.1007/s00300-007-0377-9.
Volkmann LA, Hodges KE (2021) Post-fire movements of Pacific marten (Martes caurina) depend on the severity of landscape change. Movement Ecology 9, article id 49. https://doi.org/10.1186/s40462-021-00286-2.
Wallgren M, Bergström R, Danell K, Skarpe C (2009) Wildlife community patterns in relation to landscape structure and environmental gradients in a Swedish boreal ecosystem. Wildlife Biol 15: 310–318. https://doi.org/10.2981/08-045.
Webster KL, Beall FD, Creed IF, Kreutzweiser DP (2015) Impacts and prognosis of natural resource development on water and wetlands in Canada’s boreal zone. Environ Rev 23: 78–131. https://doi.org/10.1139/er-2014-0063.
Westerfelt P, Weslien J, Widenfalk O (2018) Population patterns in relation to food and nesting resource for two cavity-nesting bee species in young boreal forest stands. Forest Ecol Manag 430: 629–638. https://doi.org/10.1016/j.foreco.2018.08.053.
Wiebe PA, Fryxell JM, Thompson ID, Börger L, Baker JA (2013) Do trappers understand marten habitat? J Wildlife Manage 77: 379–391. https://doi.org/10.1002/jwmg.471.
Yahner RH, Mahan CG (1997) Effects of logging roads on depredation of artificial ground nests in a forested landscape. Wildlife Soc B 25: 158–162.
Yamada Y, Sasaki H, Harauchi Y (2010) Effects of narrow roads on the movement of carabid beetles (Coleoptera, Carabidae) in Nopporo Forest Park, Hokkaido. J Insect Conserv 14: 151–157. https://doi.org/10.1007/s10841-009-9236-z.
Yemshanov D, Haight RG, Liu N, Rempel R, Koch FH, Rodgers A (2022) Exploring the tradeoffs among forest planning, roads and wildlife corridors: a new approach. Optim Lett 16: 747–788. https://doi.org/10.1007/s11590-021-01745-w.
Yip DA, Bayne EM, Sólymos P, Campbell J, Proppe D (2017) Sound attenuation in forest and roadside environments: implications for avian point-count surveys. Condor 119: 73–84. https://doi.org/10.1650/CONDOR-16-93.1.
Zhang JJ, Mårald E, Bjärstig T (2022) The recent resurgence of multiple-use in the Swedish forestry discourse. Soc Natur Resour 35: 430–446. https://doi.org/10.1080/08941920.2022.2025550.
Zimmermann B, Nelson L, Wabakken P, Sand H, Liberg O (2014) Behavioral responses of wolves to roads: scale-dependent ambivalence. Behav Ecol 25: 1353–1364. https://doi.org/10.1093/beheco/aru134.
Total of 177 references.
Send to email
