Mikko Hyppönen  ,
Ville Hallikainen,
Juhani Niemelä,
Pasi Rautio
,
Ville Hallikainen,
Juhani Niemelä,
Pasi Rautio
The contradictory role of understory vegetation on the success of Scots pine regeneration
Hyppönen M., Hallikainen V., Niemelä J., Rautio P. (2013). The contradictory role of understory vegetation on the success of Scots pine regeneration. Silva Fennica vol. 47 no. 1 article id 903. https://doi.org/10.14214/sf.903
Abstract
In North-East Finland, severe problems have been encountered in the natural regeneration of Scots pine (Pinus sylvestris L.) on sites where regeneration through site preparation usually is quite successful. We hypothesized that in that area understory vegetation, especially heather (Calluna vulgaris), crowberry (Empetrum hermaphroditum), mosses and lichens, could play a key role in this pattern. We found that in general, ground- and field-layer vegetation tends to be in a negative relationship with the establishment, growth and survival of pine seedlings. Some positive relationships were also observed. Lingonberry (Vaccinium vitis-idea) tended to improve seedling height growth. Heather, instead, seemed to have a contradictory role. It was positively related to seedling establishment but negatively to seedling growth. This dual role raises further questions about the primary reasons for the regeneration problems in North-East Finland. All in all, our results suggest that conventional methods of forest regeneration in these kinds of areas are not always effective enough and additional measures are needed. These might include severe prescribed burning along with site preparation in order to decrease the impact of the dominant ground- and field-layer vegetation on the success of Scots pine regeneration.
Keywords
site preparation;
seedling establishment;
seedling mortality;
field- and ground-layer vegetation;
modelling approach;
initial growth;
prescribed burning
-
Hyppönen,
Finnish Forest Research Institute, Rovaniemi Unit, P.O. Box 16, FI-96301 Rovaniemi, Finland
E-mail
mikko.hypponen@metla.fi
- Hallikainen, Finnish Forest Research Institute, Rovaniemi Unit, P.O. Box 16, FI-96301 Rovaniemi, Finland E-mail ville.hallikainen@metla.fi
- Niemelä, Finnish Forest Research Institute, Rovaniemi Unit, P.O. Box 16, FI-96301 Rovaniemi, Finland E-mail jn@nn.fi
- Rautio, Finnish Forest Research Institute, Rovaniemi Unit, P.O. Box 16, FI-96301 Rovaniemi, Finland E-mail pasi.rautio@metla.fi
Received 1 October 2012 Accepted 14 January 2013 Published 11 June 2013
Views 233436
Available at https://doi.org/10.14214/sf.903 | Download PDF

1 Introduction
Despite the cool climate and short growing season in the boreal forests of Northern Europe, the capacity for the natural regeneration of Scots pine is good. Natural regeneration using the seed-tree method has great potential as an inexpensive regeneration method for Scots pine, even in the northernmost parts of the conifer forest zone. Nevertheless, favourable conditions and appropriate methods are required for success under such challenging conditions (Hyppönen 2002; Varmola et al. 2004). In the absence of natural or prescribed fire, site preparation is generally necessary for the establishment of favourable seedbed conditions (Bergan 1981; Oleskog et al. 2000). Site preparation clearly improves seedling establishment (Yli-Vakkuri 1961; Ackzell 1993) and increases height growth (Karlsson and Örlander 2000; Hyppönen et al. 2001; Hagner 1962). Site preparation also decreases the variation in height growth between seedlings (Fries 1979).
In Northern Finland severe problems in the natural regeneration of Scots pine have been encountered in nutrient-poor xeric and sub-xeric sites where there are usually no problems in pine regeneration (Niemelä 2002). In a recent study by Hallikainen et al. (2007) Scots pine regeneration on these kinds of sites was generally slow and poor in spite of site preparation, but there was quite a variation in the results. Results of that study suggest that soil properties, field and ground layer vegetation, fungal infections, and reindeer grazing may be associated with slow and poor regeneration, but closer inspection is needed to reveal the ultimate causes. Consequently, this study focuses on the relationships between field- and ground-layer vegetation and natural regeneration of Scots pine.
In boreal and temperate forests, wildfire is the primary major disturbance influencing the composition and ecological effects of understory vegetation (Mallik 2003; Hille and den Ouden 2004; Nilsson and Wardle 2005). In the absence of high severity natural fire, key understory components – like dwarf shrubs, mosses and lichens – can become a dominative driver in forest regeneration, particularly in belowground processes such as decomposition and nutrient cycling. The role of such keystone species can be substantial. They can eventually start to steer forest succession, even to the point that succession turns into a retrogressive mode. In this kind of process, the keystone species can work as ecosystem-level engineers and convert coniferous forests into heaths, i.e. nutrient-poor sites with the dominance of ericaceous shrubs instead of trees (Mallik 1995; Mallik 2003; Nilsson and Wardle 2005).
Of keystone species, crowberry, heather and mosses have been shown to have particularly disadvantageous effects on the regeneration of Scots pine (Nilsson and Zackrisson 1992; Nilsson 1994; Mallik 1995; Steijlen et al. 1995). Heather is adapted to be a rapid colonizer after disturbance (Bråkenhielm and Persson 1980; Nieppola 1992). The rapid vegetative growth of heather results in increasing organic accumulation, nutrient sequestration, soil acidification, and allelochemicals (Robinson 1972; Jalal and Read 1983; Skre et al. 1998). They are thought to be the main factors in conifer regeneration failure and cause a “growth check” (interruption of growth) in disturbed (logging, fire etc.) habitats dominated by heather (Miles 1981). Yet, the interference mechanisms between heather and pine remain unclear (Norberg et al. 2001). Like heather and crowberry, mosses have been shown to have negative effects on seed germination and the growth of pine seedlings (Steijlen et al. 1995). Further, Nilsson et al. (1999) and Zackrisson et al. (1997) observed that feather moss Pleurozium schreberi in combination with crowberry can effectively block pine seedling establishment and growth.
Unlike crowberry, heather and mosses, lingonberry has not been found to be harmful to pine regeneration. On the contrary, it has been noticed that lingonberry offers good conditions for the establishment and growth of pine seedlings (Aaltonen 1919; Hertz 1934). Then again, in some vegetation removal studies (e.g. Wardle et al. 2008) lingonberry has also been found to have negative effect on pine seedlings.
For decades it has been assumed that thick reindeer lichen vegetation (Cladina spp.) inhibits the establishment of Scots pine seedlings (Aaltonen 1919; Hertz 1934; Lakari 1915). Brown and Mikola (1974) showed that lichens have negative allelopathic effects on mycorrhizas and tree seedlings. In contrast, recent studies have revealed that the effects of lichens are either neutral or even beneficial (Stejlen et al. 1995; Kytöviita and Stark 2009). However, the interplay between reindeer grazing and vegetation might create complex interactions that in turn have various effects on the establishment, initial growth, and survival of pine seedlings (Väre et al. 1996; Jonsson 1999).
In this study, we aimed to find out the relationships between understory vegetation and the regeneration of Scots pine in an area in North-East Finland where especially heather has been implicated as a contributor to the considerable problems of pine regeneration (Hallikainen et al. 2007). The study area is located in a part of Finland that is characterized by the highest abundance of heather in Finland (Salemaa 2001). Based on earlier findings (Hallikainen et al. 2007), we hypothesized that in addition to the harsh climatic conditions, the ground- and field-layer vegetation, especially heather, crowberry, mosses, and lichens, would inhibit the establishment and growth of pine seedlings and contribute to seedling mortality.
More specifically the principal aim of the study was to find out, using a modelling approach, the relationships of 1) understory vegetation and 2) key environmental variables (elevation, temperature sum, thickness of humus layer etc.) with the success of natural regeneration of Scots pine in Northern Finland. As indicators for the success of pine regeneration we used 1) establishment, 2) growth and 3) survival of seedlings.
2 Materials and methods
2.1 Sampling and measurements
The cross-sectional inventory data were collected in the summer 2003. The data consisted of three hierarchical levels: forest stand, plot and seedling level. At each level, different parameters described below, were measured or determined.
Stand level. Fifteen study stands (Fig. 1) were randomly selected from among the stands harvested for natural regeneration via establishment (seed-tree) cutting in the state-owned forests of North-East Finland in 1986–1997 (see Hallikainen et al. 2007). The forest stands represented the following site types: barren Cladina (ClT) type (1 stand), xeric Myrtillus-Calluna-Cladina (MCClT) type (5 stands), sub-xeric Empetrum-Myrtillus (EMT) type (8 stands), and mesic Ledum-Myrtillus (LMT) type (1 stand) (Cajander 1913). Typical field-layer dwarf shrubs in the study area are lingonberry, bilberry (Vaccinium myrtillus), crowberry, bog bilberry (Vaccinium uliginosum) and heather.
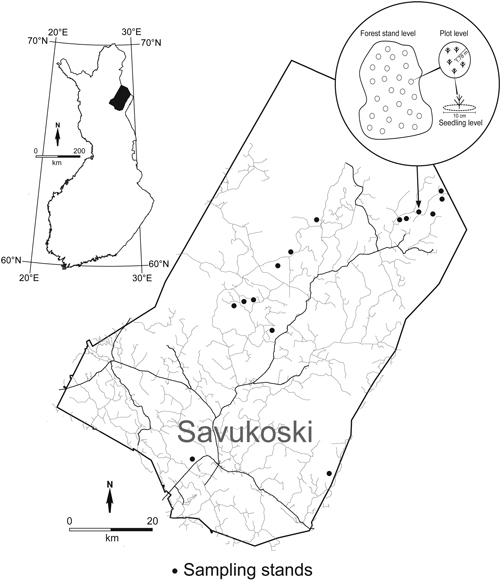
Fig. 1. Location of the regeneration stands and schematic illustration of data hierarchy. Parameters were measured either at forest stand level, plot level (25 plots of 10 m2 on each stand) or seedling level (5 cm radius around a seedling).
The average temperature sum for the regenerating stands (threshold value of five degrees centigrade) for the period of 1961–1990 was estimated using a model by Ojansuu & Henttonen (1983). For each regenerating area, the temperature sum describes long-run, average temperature conditions that reflect the particular altitude and latitude of each forest stand. In this study, temperature sum is the only climatic explanatory variable that can be used reliably at stand level. Other variables measured at forest stand level were: elevation, time since seed tree cutting and number of seed trees ha–1.
Plot level. In each of the 15 stands, 25 randomly located sample plots (i.e. total of 375 plots) of 10 m2 (radius 1.78 m) were measured (Fig. 1). Location of sample plots in the stands were randomized on a geographical co-ordinate grid and localized with GPS. Measurements and assessments for sample plots included: vegetation cover; number of seedlings and saplings by tree species [total number of pine, spruce (Picea abies), and birch (Betula pendula, B. pubescens) seedlings]; the number of pine seedlings that had emerged after the seed-tree cutting; soil stoniness (no visible stones, stones in sight); thickness of humus layer; abundance of slash (no slash = 0% of ground cover, a little = 1–49%, much ≥ 50%); and the proportion of exposed mineral soil (at the moment of measurement). From here on in this article we use the term seedling to denote both seedlings and saplings. The number of seedlings recruited since the time of seed-tree cutting was determined using the age of the seedling and the known time since the seed-tree cutting and, in most cases, soil scarification.
The cover of ground- and field-layer vegetation was analysed either by species (bilberry, lingonberry, bog bilberry, crowberry, and heather) or by groups of species (Polytrichum spp., other mosses, Cladina spp., other lichens, grass and sedge species, and other vascular plants). Below the term mosses denotes the group “other mosses” and lichens denotes the group “other lichens”. The cover of different species, or groups of species, was analysed in sample plots (radius 1.78 m) using visual estimation.
Seedling level. In each of the 25 sample plots, age and height of seedlings and seedling viability (viable, dying, dead) were assessed. In addition, abundance of plant species (or groups of species) was recorded as percentages of an area of a five-centimetre radius around a seedling (Fig. 1). When modelling seedling viability the cover of different understory species at a whole plot could not be used. This small circular area (diameter 10 cm) around a seedling was assumed to represent better the growth conditions of the seedling. These measurements around individual seedlings were used in the binomial logistic regression model for the probability of the existence of a single, observed dead or dying pine seedling, described in more detail later in the paper (Model 3).
2.2 Data analysis
2.2.1 Correspondence analysis
In addition to descriptive statistical analyses like averages and deviations, the patterns in ground vegetation communities were analysed using constrained canonical analysis (CCA). CCA reveals possible interrelations of studied parameters, which helped to select appropriate parameters for the three statistical models used to analyse seedling establishment, height growth and mortality (see below). Correspondence analysis using Chi-square metrics was selected because it can handle unimodal species responses, and does not imply linear relationships between axes and species abundance (Wildi 2010). The forest stand was used as a conditioning variable (“partialled out”) in the analysis (Eskelinen and Oksanen 2006). Constraining gradient parameters closely related to the ground and field layer plant communities, such as thickness of humus, stoniness, and the number of seedlings, were included in the model. Constraining gradients are usually considered as explanatory variables in CCA analysis, but the relationships between so-called gradient variables and the vegetation composition must be considered as multi-way relations in ecology (Wildi 2010). The relationships between vegetation and gradient variables (Table 2) were tested using permutation tests.
2.2.2 Seedling establishment model
An ordered multinomial logistic model (Model 1) was used in analysing and predicting the seedling establishment of Scots pine. There are a couple of reasons to model the number of seedlings using ordered multinomial logistic model with a random factor instead of a Poisson or a negative binomial model. Firstly, there are too many sample plots (58%) with no seedlings (i.e. the data is heavily zero inflated). Thus, the Poisson or negative binomial models would yield biased result. Secondly, the multinomial model enables to model the cumulative probabilities for at least 1, 2, 3 and 4 seedlings on a 10 m2 sample plot corresponding to at least 1000, 2000, 3000 and 4000 seedling ha–1. One can also get the probability for less than 1000 seedlings ha–1. These levels have a meaningful interpretation in terms of regeneration result for practical forestry. A minimum of 2000 seedlings ha–1 denotes a sufficient regeneration success, but 1000 seedlings ha–1 denotes a need for additional planting or direct seeding (Keskimölö et al. 2007). Furthermore less than 10% of plots had more than 4 seedlings (corresponding to 4000 seedlings ha–1). Thus, it was not reasonable to try to model an infrequent phenomenon of more than 4 seedlings per plot.
Model 1 consisted of the calculation of the cumulative probabilities for the categories depicting the number of pine seedlings established since seed-tree cutting on a sample plot: ≥1, ≥2, ≥3, and ≥4. The model for expected value (E) included a random factor and two hierarchical levels:
![]()
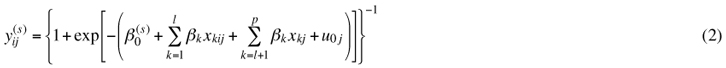
where ![]() = observed cumulative proportion for a sample plot ij;
= observed cumulative proportion for a sample plot ij; ![]() = cumulative response probability for a sample plot;
= cumulative response probability for a sample plot; ![]() = probability of a sample plot representing the value of a certain categorical response variable; h = index for a certain category (ordered categories of ≥1, ≥2, ≥3, and ≥4 seedlings per plot); t = total for categories,
= probability of a sample plot representing the value of a certain categorical response variable; h = index for a certain category (ordered categories of ≥1, ≥2, ≥3, and ≥4 seedlings per plot); t = total for categories, ![]() ;
; ![]() fixed part constant, specific for the cumulative models; βk = fixed part coefficient, general for all levels of response variable; k = explanatory variable index, k = 1,…,l, l + 1,…,p; i = sample-plot index; j = stand index; xkij = explanatory variables at sample-plot level; and xkj = explanatory variables at forest stand level. The term u0j denotes the random part of the model, consisting of the random intercept for a forest stand.
fixed part constant, specific for the cumulative models; βk = fixed part coefficient, general for all levels of response variable; k = explanatory variable index, k = 1,…,l, l + 1,…,p; i = sample-plot index; j = stand index; xkij = explanatory variables at sample-plot level; and xkj = explanatory variables at forest stand level. The term u0j denotes the random part of the model, consisting of the random intercept for a forest stand.
Assuming that the random intercept (general for all levels of the response variable) is normally distributed, [u0jtu] ~ N(0,σu2), where t describes the number of categories, and assuming there are two categories, r and s, the covariance of the observed cumulative proportion between the categories is
![]()
in which nij is the total number of responses over all categories, and α specifies extra-multinomial variation (Yang et al. 2001).
In seedling establishment model (Model 1), as well as in median height model (Model 2) and mortality model (Model 3) ecologically interpretable interactions were tested to enhance the performance of the models. None of these interactions turned out to be statistically significant and are hence not reported in the results (Tables 3–5).
2.2.3 Median height model
The linear mixed model with the random factor for median seedling height (Model 2) on a sample plot was:

where k = explanatory variable index; i = sample-plot index; j = stand index; yij = of the median height of the seedlings on a plot; xkij, xkj = explanatory variables; β0, βk = fixed parameters; u0j random stand effect; and eij = residual error term or random sample plot effect. The response variable was log-transformed for the model.
The predicted values using the fixed part of the model at the original scale were calculated using bias correction with estimated variances for different levels, u and e (Flewelling and Pienaar 1981) as follows:
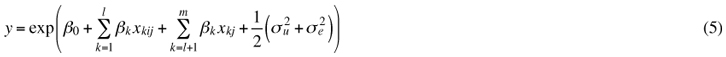
2.2.4 Mortality model
The logistic regression model with the random factors for the probability of the dead or dying pine seedling (Model 3) was:
![]()
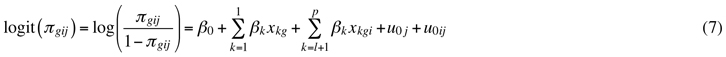
where g = seedling level index; i = sample plot level index; j = forest stand level index; πgij = probability that a seedling is dead or dying; xkgi, xkg = explanatory variables in a sample plot and seedling level, β0, βk = fixed parameters; u0j = random stand effect, and u0ij = random sample plot effect. The symbols k, l and p were similar to the parameters presented in the above models. A logit link function was used in this generalized linear model (McCullogh and Searle 2001). Similarly to the multinomial model, the dispersion parameter was allowed to be estimated freely (the scale was not assumed to be 1). All the explanatory variables in the binomial logistic model were measured at the sample plot or seedling level. The model estimates the probability for a single seedling on a plot to be classified as dead or dying. It is probable that there have been seedlings that have regenerated, died and disappeared and escaped our notice. These seedlings have not been taken into account in the modelling.
The fact that some of the dead seedlings in the plots could have remained undetected could have led the model to underestimate the proportion of dead or dying seedlings. On the other hand, in the site types used in this study (xeric and sub-xeric sites) even dead seedlings remain visible for many years if not for decades, hence it is presumable that most of the established dead seedlings were detected. Further, instead of modelling the proportions of healthy and dying or dead seedlings at sample plot level, the response in the model was the health condition of a single perceived seedling that has obviously been established since seed-tree cutting, the predictors being the environmental factors measured in the immediate vicinity of the seedlings. Although the coefficients in this model may be slightly biased, the model could distinguish how the environmental conditions differ in the surroundings of healthy and dead or dying seedlings, thus providing indicative implications about “good” and “bad” micro-sites for seedlings.
2.2.5 Fit of the models
The performance of the logistic regression models were estimated using a Receiver Operating Characteristic (ROC) curve associated with a fitted binary-response model. This is a plot of the sensitivity against 1-specificity values associated with the observations’ predicted probabilities of the events. The area under the ROC-curve suggests model’s ability to predict the events correctly. The ROC-curve was calculated for four cumulative responses of the multinomial logistic model. The coefficient of determination (R2) for the linear height model was defined by 1 minus the proportion of residual variance calculated using the fixed part of the model (e.g. Lappi 1993).
The multinomial logistic model was developed using MLwiN (version 2.15), while the vegetation analysis (CCA ordination), the binomial logistic model (library glmmPQL), and the height model were developed using R statistical environment (version 2.10.1, R Core Team 2009).
3 Results
3.1 Forest stand description
On average, slightly over ten years had elapsed since the seed-tree cutting (Table 1). The number of seed trees per hectare varied substantially between stands. The stands were located in fairly harsh climatic conditions characterized by high altitude a.s.l. and a low temperature sum. The vegetation in the sample plots mostly indicated xeric or sub-xeric site types, but mesic plots were also encountered. The cover of the main dwarf shrub species varied a lot between sample plots within stands, and maximum values for individual plots were quite high: heather 80%, lingonberry 50%, bilberry 60%, and crowberry 60%, respectively.
| Table 1. The characteristics of the forest stands (the averages at stand level). Expression “est. since cut” denotes the seedlings that have been emerged since seed-tree cutting. | |||||||
| Mean | 95% cl. lower | 95% cl. upper | Sd | Min | Max | Median | |
| Temperature sum, d.d. | 690 | 678 | 701 | 23 | 659 | 757 | 694 |
| Elevation, m asl | 257 | 242 | 272 | 30 | 207 | 310 | 260 |
| Time since seed-tree cut, years | 12 | 10 | 13 | 4 | 6 | 17 | 11 |
| Number of seed trees ha–1 | 33 | 19 | 48 | 28 | 13 | 120 | 24 |
| Thickness of humus layer, cm | 2.0 | 1.4 | 2.6 | 1.2 | 0.5 | 5.8 | 1.7 |
| Cover of crowberry, % | 15.4 | 11.3 | 19.6 | 8.2 | 0.7 | 33.0 | 13.5 |
| Cover of heather, % | 11.1 | 5.9 | 16.2 | 10.2 | 0.0 | 34.2 | 11.8 |
| Cover of lingonberry, % | 2.0 | 1.1 | 2.8 | 1.7 | 0.4 | 6.5 | 1.7 |
| Cover of bilberry, % | 6.3 | 3.9 | 8.8 | 4.8 | 0.4 | 16.2 | 5.9 |
| Cover of Polytrichum spp., % | 8.8 | 5.1 | 12.5 | 7.3 | 1.1 | 23.3 | 7.9 |
| Cover of other mosses, % | 21.6 | 15.5 | 27.7 | 12.1 | 2.7 | 44.7 | 24.0 |
| Cover of reindeer lichens, % | 16.1 | 9.8 | 22.4 | 12.4 | 0.4 | 44.8 | 15.0 |
| Cover of other lichens, % | 3.2 | 1.5 | 4.8 | 3.2 | 0.0 | 10.5 | 1.6 |
| Cover of exposed mineral soil, % | 5.2 | 2.0 | 8.5 | 6.1 | 0.0 | 16.4 | 1.5 |
| Number of pine seedlings (est. since cut) ha–1 | 1387 | 658 | 2116 | 1441 | 160 | 5200 | 1120 |
| Total number of pine seedlings / saplings ha–1 | 1874 | 943 | 2806 | 1840 | 240 | 7033 | 1280 |
| Total number of spruce seedlings /saplings ha–1 | 273 | 92 | 455 | 359 | 0 | 1120 | 120 |
| Total number of birch seedlings / saplings ha–1 | 1167 | 662 | 1672 | 998 | 40 | 3680 | 920 |
| Age of pine seedlings (est. since cut), years | 6 | 5 | 7 | 2 | 3 | 10 | 6 |
| Height of seedlings (est. since cut), cm | 17.8 | 12.5 | 23.1 | 10.5 | 5.0 | 43.2 | 16.3 |
Soil scarification had been carried out in 87% of the sample stands. The proportion of exposed mineral soil varied from 0% to 16%, the average being about 5% at stand level. The number of seedlings by species varied considerably between stands (Table 1). Their spatial distribution was aggregated. The proportion of empty sample plots (no tree seedlings) by tree species was: Scots pine 45%, Norway spruce 84%, and birches 59%. In 59% of the sample plots no pine seedlings had emerged since seed-tree cutting.
3.2 Vegetation communities
The vegetation mostly reflected xeric or sub-xeric types both at stand and at plot level. The x-axis (CCA1) in Fig. 3 represents the mesic–xeric dimension. Moss species (group other mosses) dominated the ground layer vegetation of the most mesic patches, contrary to Polytrichum species that can be seen in the xeric end of the x-axis (Fig. 2). In addition to Polytrichum mosses, lichens (Cladina species and group other lichens) characterized the ground layer of the xeric parts of the stands. Bilberry was found on more xeric patches than the other Vaccinium species (in accord with Cajander’s (1913) site type classification for northern areas), and its abundance was closely related to stoniness. Heather characterized the xeric site type rather than the mesic one, but its abundance varied a lot, and its cover did not strongly correlate with other plant species (Table 2, Fig. 2)
| Table 2. The results of permutation tests for the CCA analysis. The test is for the marginal effects of the constraining terms. N.Perm denotes the number of permutations. | |||||
| Df | Chi-square | F | N.Perm | p | |
| Thickness of humus layer, cm (HUM) | 1 | 0.009 | 2.690 | 299 | 0.017 |
| Stoniness, (S1, categories: 1 = stony, 0 = not stony, measured on a sample plot) | 1 | 0.014 | 4.352 | 199 | 0.005 |
| Total number of pine seedlings on a sample plot (PINE) | 1 | 0.014 | 4.389 | 199 | 0.005 |
| Total number of spruce seedlings on a sample plot (SPRUCE) | 1 | 0.008 | 2.347 | 399 | 0.023 |
| Residual | 356 | 1.166 | |||
The y-axis (CCA2) in Fig. 2 represents the aggregation, abundance and variation in the coverage of the species. Heather has the greatest values in CCA2 denoting the strong aggregation, heather was found only on the half of the sample plots. In addition, the coverage of heather on the plots is among the highest if it is present, and the variation among the plots is high. On the other hand, crowberry, bilberry and lingonberry were found on 90–95% of the sample plots, and the variation in the cover was remarkably small compared to heather.
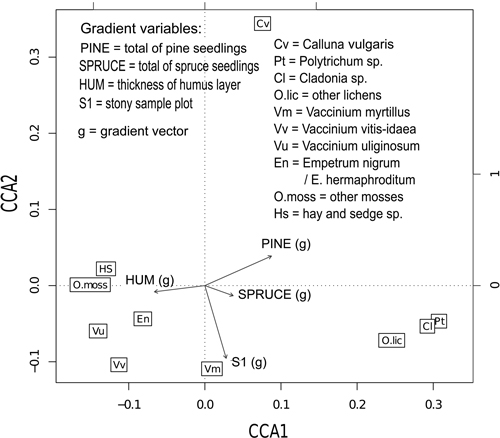
Fig. 2. Canonical correspondence analysis (CCA) plot describing the relations involved in the ground- and bottom-layer vegetation and the statistically significant (at 5% risk level) gradient variables indicated by gradient vectors.
A thick humus layer was closely related to mesic vegetation, whereas the abundance of pine and spruce seedlings was closely related to lichen-dominated sites (Table 2, Fig. 2). Increasing gradient of pine seedlings tended slightly towards heather, suggesting that heather dominance slightly facilitates the establishment of pine seedlings.
About 70% of the total inertia (1.682) in the CCA-analysis fell to the unconstrained component (1.167), 28% (0.465) to the conditional forest stand and the rest, 3% (0.051) to the constraining gradients.
3.3 Seedling establishment since seed-tree cutting
The seedling establishment model (Table 3, Model 1) indicated a significant but not very strong positive relationship between the cover of heather and the establishment of pine seedlings (Fig. 3a). According to the model, if the cover of heather increases from 0% to 80%, the establishment probability increases from 5% to 20%, depending on the required number of seedlings in a sample plot. The cover of moss species, such as Pleurozium schreberi and certain Dicranum species (other than Polytrichum spp.), as well as a thick humus layer, considerably inhibits the establishment of seedlings (Fig. 3b and 3c). The probability to achieve even one seedling on a plot is very low if the site is 80% covered with moss (Fig. 3b). Similarly, the probability gets lower as the humus layer gets thicker (Fig. 3c).
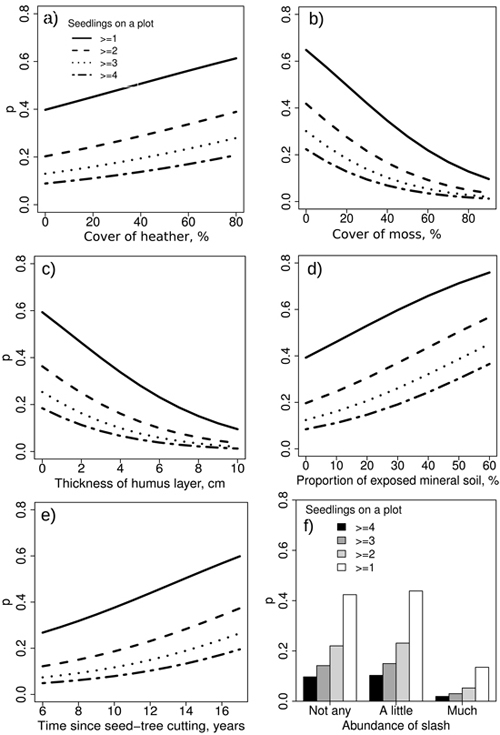
Fig. 3. The predicted seedling establishment probabilities by independent variables in Model 1. The covers were measured at plot level.
The cover of exposed mineral soil has a positive effect on the probability of seedling establishment. At a maximum soil exposure of 60% the probability of seedling establishment is 35–75% (Fig. 3d). Time since seed-tree cutting affects the seedling establishment probability as well, but the statistical significance of the variable is marginal (Table 3, Fig. 3e). As an example, according to the model predictions at 15 years post-seed-tree cutting and with 50% of mineral soil exposed, the predictions for at least 2, 3 and 4 seedlings on a plot (corresponding 2000, 3000 and 4000 seedlings ha–1) are 60%, 50% and 40%, respectively.
| Table 3. The ordered multinomial regression model with a random factor for predicting the numbers of seedlings on a sample plot emerged after the seed-tree cutting (Model 1). The joined Wald tests were calculated as a general test for the categorical variables. Approximate confidence intervals for random and scale parameters are presented. Pl denotes measurement at plot and st at stand level (in the parenthesis). | |||||
| Variable | Estimate | Std.error | Wald | df | p |
| Fixed effects | |||||
| Intercept (≥ 4 seedling on a plot, pl) | –2.918 | 1.119 | 6.797 | 1 | 0.009 |
| Intercept (≥ 3 seedling on a plot, pl) | –2.423 | 1.116 | 4.714 | 1 | 0.030 |
| Intercept (≥ 2 seedling on a plot, pl) | –1.787 | 1.113 | 2.575 | 1 | 0.109 |
| Intercept (≥ 1 seedling on a plot, pl) | –0.603 | 1.110 | 0.295 | 1 | 0.587 |
| Time since seed-tree cutting, years (st) | 0.164 | 0.088 | 3.476 | 1 | 0.062 |
| Thickness of humus layer, cm (pl) | –0.332 | 0.076 | 19.089 | 1 | 0.000 |
| Cover of moss species, % (pl) | –0.039 | 0.007 | 30.527 | 1 | 0.000 |
| Cover of heather, %, (pl) | 0.014 | 0.006 | 5.037 | 1 | 0.025 |
| Cover of exposed mineral soil, % (pl) | 0.034 | 0.015 | 4.919 | 1 | 0.027 |
| Abundance of slash and other cutting waste (joined test, pl) | 12.619 | 2 | 0.002 | ||
| Much | –1.948 | 0.578 | 11.341 | 1 | 0.001 |
| A little | 0.080 | 0.223 | 0.129 | 1 | 0.719 |
| Not any (ref. category) | 0.000 | 0.000 | - | - | - |
| Random effects | Estimate | 95 % confidence intervals | |||
| Variance of random intercept of forest stand σj2 | 1.162 | 0.244–2.080 | |||
| Scale parameter (compare to 1) | 0.814 | 0.754–0.874 | |||
A small amount of slash does not affect the probability of seedling establishment, but abundant slash strongly decreases the probability (Fig. 3f). According to the model, the probability of having at least one pine seedling in a sample plot (1000 seedlings ha–1) within ten years is about 43% if there is no slash on a site. The respective probabilities for at least 2, 3 and 4 seedlings are 14%, 8% and 5%.
3.4 Seedling height
According to the seedling height model (Model 2), the predictions for the seedling of a median height on a plot show that the cover of lingonberry is related to faster-than-average growth (Table 4). In contrast to lingonberry, heather is related to slower height growth. A fifteen-year-old seedling growing on a site where the cover of heather is 50% is around ten centimetres lower in height than a seedling of the same age growing on a site with no heather cover (Fig. 4). Reindeer lichens (Cladina spp.) and Polytrichum species did not have a statistically significant relation on seedling height in the analyses.
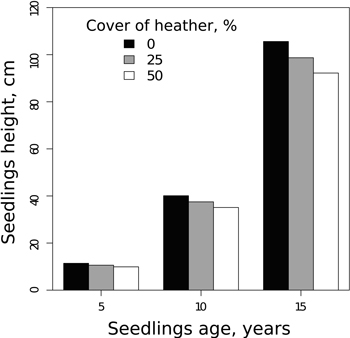
Fig. 4. The predictions for the effect of heather cover on the average height of the pine seedlings calculated at 5, 10 and 15 year seedling age (Model 2).
| Table 4. The estimates and significance tests for the model of pine seedling height (median height on a plot) (Model 2). The response variable is the logarithm of the height of the seedlings on a sample plot that were emerged since seed-tree cutting. The model was done for the forest stands where the average of temperature sum during 1960–1991 was < 750 (one stand omitted), and the maximum age was 15 years (one sample plot omitted). The R2 basing on the fixed predictions was 71.5%. Pl denotes measurement at plot and st at stand level (in the parenthesis). | |||||
| Variable | Estimate | Std. error | df | t | p |
| Fixed part | |||||
| Intercept (pl) | –5.668 | 1.524 | 126 | –3.719 | 0.000 |
| Age of seedling of median height, years (pl) | –0.195 | 0.071 | 126 | –2.755 | 0.007 |
| Square root of age of median height seedling, years (pl) | 2.366 | 0.351 | 126 | 6.733 | 0.000 |
| Cover of heather, % (pl) | –0.002 | 0.001 | 126 | –1.733 | 0.086 |
| Cover of lingonberry, % (pl) | 0.019 | 0.006 | 126 | 2.878 | 0.005 |
| Temperature sum, d.d. (st) | 0.005 | 0.002 | 12 | 2.602 | 0.021 |
| Random part | Estimate | 95% confidence intervals | |||
| Variance of random intercept of forest stand σj2 | 0.002 | 0.000–0.742 | |||
| Residual σij2 | 0.088 | 0.069–0.112 | |||
Temperature sum had a positive coefficient in the model. However, since the data included no more than 15 stands, this variable needs to be considered as a tentative one; nevertheless it was included as an ecologically highly relevant covariate.
3.5 Dead and dying seedlings (mortality)
The logistic binomial regression model, predicting the probability of a single pine seedling either being dead or dying, shows that the probability increases with increasing age of the seedling (Table 5). Roughly half of the counted seedlings are predicted to be dead or dying at the age of fifteen years (Fig. 5a and 5b).
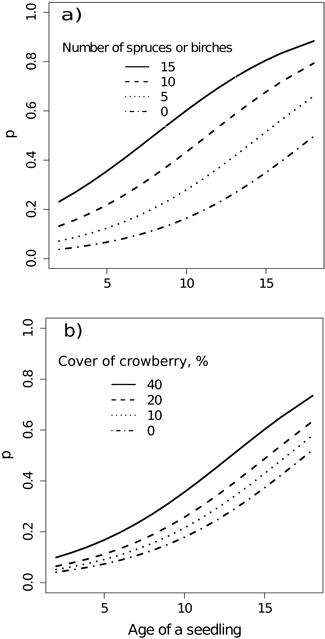
Fig. 5. Predicted probability (p) of the occurrence of dead or dying seedlings by the independent variables (Model 3). The number of spruce and birch seedlings was measured at plot level and the cover of crowberry at the base of a seedling (at seedling level).
| Table 5. The estimates of the model for predicting the probability of a seedling’s poor health (seedling classified as dead or dying) (Model 3). The maximum of the total number of seedlings was restricted as <20 seedlings on a plot for computational reasons. Thus the total number of seedlings in the model was 491 out of 510. Pl denotes measurement at plot and se at seedling level (in the parenthesis). | |||||
| Variable | Estimate | Std.error | t-value | df | p |
| Fixed effects | |||||
| Intercept (se) | –4.208 | 0.484 | 333 | –8.690 | 0.000 |
| Age of a seedling, years (se) | 0.202 | 0.040 | 333 | 5.007 | 0.000 |
| Total number of seedlings (pl) | 0.081 | 0.040 | 139 | 2.084 | 0.039 |
| Cover of crowberry at seedling’s base (se) | 0.026 | 0.008 | 333 | 3.198 | 0.002 |
| Cover of lichens at seedling’s base (se) | 0.015 | 0.008 | 333 | 1.994 | 0.047 |
| Random effects | Estimate | 95% confidence intervals | |||
| Variance of random intercept of forest stand σj2 | 0.024 | 0.000–1.230e+03 | |||
| Variance of random intercept of sample plot σij2 | 1.429 | 0.800–2.553 | |||
| Scale parameter (compare to 1) | 0.624 | 0.542–0.719 | |||
The probability of a seedling being in poor vigour increases by about 20–40% when the number of other seedlings (spruce and birch) besides pine increases in a sample plot. The probability also increases with increasing age of the pine seedlings emerged since seed-tree cutting (Fig. 5). When the number of other seedlings reaches its maximum of 15 per plot in the data, corresponding to the density of 15 000 seedlings ha–1 the predicted probability for a dead or dying pine seedling is some 80%.
We also observed a clear relationship between the probability of a dead or dying pine seedling being present on a plot and the abundance of crowberry in the immediate vicinity (within 5 cm radius) of seedlings (Table 5, Fig. 5b). For example, a 40% increase in crowberry cover causes about a 20% increase in the probability of a dead or dying pine seedling at the age of around fifteen years (Fig. 5b). The same kind of relationship is found between dead or dying pine seedlings and lichens, respectively. For instance, a 60% increase in lichen cover at the seedling base causes about a 20% increase in the probability of a dead or dying pine seedling at the age of fifteen years (Table 5). The correlation between the cover of lichens and crowberry was weak, the value of Spearman correlation coefficient being about –0.1. Thus, they were taken in the same model, because both could be considered as ecologically meaningful when talking about the probability of death of the seedlings.
3.6 The performance of the models
The classification efficiency for the ordered multinomial model suggested by the ROC-curves for the four ordered categories varied from 77.3–83.0%, the greatest value being for the model of ≥ 4 seedlings on a plot. The predicted probabilities for the multinomial model remained few percents lower compared to the observed probabilities, in general. Furthermore, the classification efficiency for the binary logistic model for predicting seedling’s bad health condition was 90%. The specificity of the model was 76%. Thus, the model could better predict poor than good seedling conditions. The coefficient of determination for the height model was 76.6%.
4 Discussion
The average regeneration result was quite poor and the restocking process had been slow. Poor natural regeneration of Scots pine in harsh climate conditions is known to be associated with northern latitude, high elevation, and low temperature sum causing small seed crops, a low seedling establishment rate, slow height growth and high seedling mortality (Varmola et al. 2004; Hyppönen et al. 2005; Juntunen and Neuvonen 2006). In North-East Finland, the regeneration of Scots pine has been even more difficult than elsewhere in the timberline regions (Niemelä 2002; Hallikainen et al. 2007). One reason has been assumed to be ground vegetation (Hallikainen et al. 2007). This assumption is supported by the fact that the study area is one of the most typical parts of the range of heather (Salemaa 2001).
The abundance of certain ground- and field layer species were either negatively or positively related to the establishment, growth and survival of pine seedlings. Interestingly, one of these, heather, indicated both negative and positive relationships in different phases of seedlings’ life span. A cover of heather had a positive correlation on seedling establishment (Model 1) but a negative correlation on seedling growth (Model 2). This dual role seems to be inconsistent but similar observations have been made in some early studies (Kangas 1931; Sarvas 1949). According to these, heather seems to promote the establishment of pine seedlings and to shelter pine seedlings from drought. This so-called nurse-plant effect is a classic example of facilitation, i.e. positive species interaction (Bruno et al. 2003). Another protective function of heather might be that it shelters pine seedlings from the mechanical injuries of reindeer grazing (i.e. functioning as biotic refuge for pine).
It is also possible that immediately after felling, pine has time to emerge while heather has not yet fully colonized the area and does not inhibit the establishment of pine through competition and/or allelopathy. Later, when heather increases in abundance, the competition with pine seedlings intensifies and seedling mortality increases (Aaltonen 1919; Hertz 1934). Heather is known to be a strong competitor with pine seedlings (Lehto 1956), a feature that might arise from its allelopathic properties (Miles 1981; Mallik 1995). Establishment cutting, site preparation and prescribed burning are not always effective enough to control heather (Norberg et al. 2001) because heather recolonizes rapidly after a disturbance (Skre et al. 1998).
In contrast to heather, lingonberry had a positive correlation on the height growth of pine seedlings (Model 2). This may be attributable to its strong positive effect on litter decomposition and soil microbial activity (Wardle and Zackrisson 2005).
Moss cover had a negative influence on pine seedling establishment (Model 1). This result is in line with many earlier findings. For example, Hertz (1934) noticed that regeneration success was poorer with a higher abundance of feather moss. Similarly, Steijlen et al. (1995) found feather moss to have negative chemical effects on the emergence of pine seeds but not on the growth of radicles. In field experiments conducted by Zackrisson et al. (1997) the negative influence of feather moss on Scots pine seed germination and seedling survival was suggested to be attributable to moisture, chemical interference, and limitation of nutrient availability. In their experiment (Zackrisson et al. 1997) mortality of pine seedlings was higher on plots where the moss layer was dominant, compared to plots where lichens dominated. Germination, seedling establishment and pine growth were all better when mosses and crowberry were removed.
In the present study, crowberry seemed to negatively affect the survival of pine seedlings. In earlier studies, crowberry has had a strong negative effect on the regeneration, growth, and survival of Scots pine through allelopathy and resource competition (Nilsson 1994; Zackrisson et al. 1995, Zackrisson et al. 1997; Jonsson 1999). Lichens also seemed to increase the mortality of seedlings (Model 3). This result is in accordance with some earlier studies (Lakari 1915; Brown and Mikola 1974). Conversely, some recent studies indicated that lichens would be a better establishment substrate than many others, such as mosses and crowberry (Steijlen et al. 1995; Zackrisson et al. 1995; den Herder et al. 2003). One must take into account, however, that reindeer grazing and trampling complicates the interpretation of the impact of lichen cover. In places where there is an abundant lichen cover, there is generally strong reindeer grazing pressure in the reindeer herding area. Grazing tends to reduce the lichen layer and this seems to change the vegetation towards a type that is dominated by mosses, small dwarf shrubs, bare soil and minute-cup lichens (Cladonia spp.) (Suominen and Olofsson 2000; den Herder et al. 2003). The grazing effect was highlighted in a recent enclosure experiment by Lappalainen (2010) in northern Finland where the number of pine seedlings was double in the ungrazed (fenced) area compared to the grazed one.
In addition to heather and moss cover, other explanatory variables affecting the establishment of pine seedlings were (i) time since seed-tree cutting, (ii) cover of exposed mineral soil, (iii) thickness of humus layer and (iv) abundance of slash. The first two variables had a positive relation with the number of seedlings, as expected (Varmola et al. 2004; Hyppönen et al. 2005), while the other two were shown to decrease the number of seedlings. The negative effects of a thick humus layer on seedling establishment have been reported frequently (Niemistö et al. 1993; Hyppönen 2002). It is also known that a moderate amount of slash has a positive effect on seedling establishment (Lehto 1956, Lehto 1969) while abundant slash has a negative influence (Jonsson 1999). Accordingly, Karlsson et al. (2002) have noticed that slash removal has a positive effect on pine regeneration in clear-cuts.
Due to the large number of studied explanatory variables, typical to inventory data (Table 1), we didn’t inspect all the possible interactions between them. Instead, we focused on the interactions between the significant main effects in the models. These interactions could mean that, for example, the effect of heather on the pine regeneration is dependent on the temperature sum or the presence of lingonberry. However, the interaction terms between the significant variables selected to the models were not statistically significant. Further, because our main interest was to find out if the explanatory variables have any effects on regeneration success on average, we ended up with the main effects models presented above. Although we did not find any significant interactions it is still possible that, for instance, the effect of heather on pine regeneration success is dependent on temperature, which could be found with e.g. larger temperature gradient.
5 Conclusions
Our hypothesis that the ground and field layer vegetation, especially heather, crowberry, mosses, and lichens would have negative effects on pine regeneration, growth and survival of pine seedlings was generally supported. Despite this, we detected some positive relationships: pine seedlings growing in the vicinity of lingonberries were taller than those growing without lingonberries in their neighbourhood. Furthermore, heather turned out to have a dual role in the development of pine seedlings; it was positively related to seedling establishment but negatively to seedling growth.
Considering the fact that, despite soil preparation, many of the measured understory vegetation parameters were negatively related to pine regeneration success, our results suggest that conventional methods of forest regeneration in these kinds of areas are not always effective enough; additional measures are needed. These might include a severe prescribed burning in addition to site preparation to decrease the impact of dominant ground and field layer vegetation.
Acknowledgements
The study was conducted in co-operation between Metsähallitus and the Finnish Forest Research Institute. We are grateful to the organizations and their personnel for technical and financial support. Sauli Valkonen deserves our special thanks for reading the manuscript and providing valuable comments and advice. We also thank Pia Kangas, who collected the data, Raimo Pikkupeura and Jouni Hyvärinen, who assisted with technical details and Zoe Koivu who revised the English language. Comments by two anonymous reviewers helped to improve the paper.
References
Aaltonen V.T. (1919). [Natural regeneration of upland forests in Finnish Lapland, I]. Communicationes Instituti Forestalis Fenniae 1. 375 p. [In Finnish with German summary].
Ackzell L. (1993). A comparison of planting, sowing and natural regeneration for Pinus sylvestris (L.) in boreal Sweden. Forest Ecology and Management 61: 229–245.
Bergan J. (1981). Regeneration of Scots pine forests in Troms and Finnmark. Norsk Institutt for Skogforskning, Rapport 10. 69 p. [In Norwegian with English summary].
Bråkenhielm S., Persson H. (1980). Vegetation dynamics in developing Scots pine stands in Central Sweden. In: Persson T. (ed.). Structure and function of northern coniferous forests – an ecosystem study. Ecology Bulletin 32: 139–152.
Brown R.T., Mikola P. (1974). The influence of fruticose soil lichens upon the mycorrhizae and seedling growth of forest trees. Acta Forestalia Fennica 141. 23 p.
Bruno J.F., Stachowicz J.J., Bertness M.D. (2003). Inclusion of facilitation into ecological theory. Trends in Ecology and Evolution 18(3): 119–125.
Cajander A.K. (1913). Über Waldtypen. [About forest site types]. Acta Forestalia Fennica 1. 175 p. [In German].
Eskelinen A., Oksanen J. (2006). Changes in the abundance, composition and species richness of mountain vegetation in relation to summer grazing by reindeer. Journal of Vegetation Science 17: 245–254.
Flewelling J.W., Pienaar L.V. (1981). Multiplicative regression with lognormal errors. Forest Science 27: 281–289.
Fries J. (1979). Natural regeneration within the Siljanfors experimental park. Sveriges Skogsvårdsförbunds Tidskrift 2: 96–111. [In Swedish with English summary].
Hagner S. (1962). Natural regeneration under shelterwood stands. An analysis of the method of regeneration, its potentialities and limitations in forest management in middle North Sweden. Meddelanden från Statens Skogsforskningsinstitut, Band 52(4). 263 p. [In Swedish with English summary].
Hallikainen V., Hyppönen M., Hyvönen J., Niemelä J. (2007). Establishment and height development of harvested and naturally regenerated Scots pine near the timberline in North-East Finnish Lapland. Silva Fennica 41(1): 71–88.
den Herder M., Kytöviita M.-M., Niemelä P. (2003). Growth of reindeer lichens and effects of reindeer grazing on ground cover vegetation in a Scots pine forest and a subarctic heathland in Finnish Lapland. Ecography 26: 3–12.
Hertz M. (1934). [Studies of the importance of a substrate on the regeneration of Scots pine in the upland forests in South Finland]. Communicationes Instituti Forestalis Fenniae 20(2). 98 p. [In Finnish with German summary].
Hille M., den Ouden J. (2004). Improved recruitment and early growth of Scots pine (Pinus sylvestris L.) seedlings after fire and soil scarification. European Journal of Forest Research 123: 213–218.
Hyppönen M. (2002). Natural regeneration of Scots pine using the seed tree method in Finnish Lapland. Finnish Forest Research Institute, Research Papers 844. 69 p. [In Finnish with English summary].
Hyppönen M., Hyvönen J., Mäkitalo K., Riissanen N., Sepponen P. (2001). [The effect of site preparation on the development of the natural pine sapling stand in Finnish Lapland]. Metsätieteen aikakauskirja 1/2001: 5–18. [In Finnish].
Hyppönen M., Alenius V., Valkonen S. (2005). Models for the establishment and height development of naturally regenerated Pinus sylvestris in Finnish Lapland. Scandinavian Journal of Forest Research 20: 374–357.
Jalal M.A.F., Read D.J. (1983). The organic acid composition of Calluna heathland soil with special reference to phyto- and fungitoxicity. Plant and Soil 70: 257–272.
Jonsson B. (1999). Stand establishment and early growth of planted Pinus sylvestris and Picea abies related to microsite conditions. Scandinavian Journal of Forest Research 14: 425–440.
Juntunen V., Neuvonen S. (2006). Natural regeneration of Scots pine and Norway spruce close to the timberline in Northern Finland. Silva Fennica 40(3): 443–458.
Kangas E. (1931). [About the damage of young reproductions in Siikakangas]. Silva Fennica 17. 107 p. [In Finnish].
Karlsson C., Örlander G. (2000). Soil scarification shortly before a rich seed fall improves seedling establishment in seed-tree stands of Pinus sylvestris. Scandinavian Journal of Forest Research 15: 256–266.
Karlsson M., Nilsson U., Örlander G. (2002). Natural regeneration in clear-cuts: effects of scarification, slash removal and clear-cut age. Scandinavian Journal of Forest Research 17: 131–138.
Keskimölö A., Heikkinen E., Keränen K. (2007). [Recommendations for forest management in northern Finland]. 58 p. [In Finnish].
Kytöviita M.-M., Stark S. (2009). No allelopathic effect of the dominant forest-floor lichen Cladonia stellaris on pine seedlings. Functional Ecology 23: 435–441.
Lakari O.J. (1915). [Studies of the seed years of Scots pine and the age class distributions of Scots pine upland forests in North Finland]. Acta Forestalia Fennica 5(1). 211 p. [In German].
Lappalainen M. (2010). Effect of reindeer grazing on natural regeneration of Pinus sylvestris. Thesis in the University of Applied Sciences in Rovaniemi. 29 p. [In Finnish with English abstract].
Lappi J. (1993). [Methods of forest biology]. Silva Carelica 24. 182 p. [In Finnish].
Lehto J. (1969). Studies conducted in northern Finland on the regeneration of Scots pine by means of the seed tree and shelterwood methods. Communicationes Instuti Forestalis Fenniae 67(4). 140 p.
Lehto J. (1956). Studies on the natural reproduction of Scots pine on the upland soils of Southern Finland. Acta Forestalia Fennica 66. 106 p. [In Finnish with English summary].
Mallik A.U. (1995). Conversion of temperate forests into heaths: role of ecosystem disturbance and ericaceous plants. Environmental Management 19:19(5): 675–684.
Mallik A.U. (2003). Conifer regeneration problems in boreal and temperate forests with ericaceous understory: role of disturbance, seedbed limitation, and keystone species change. Critical Review in Plant Sciences 22(3&4): 341–366.
McCullogh C.E., Searle S.R. (2001). Generalized, linear, and mixed models. Wiley Series in Probability and Statistics. Wiley, New York. 358 p. ISBN 0–471–19364–X.
Miles J. (1981). Problems in heathland and grassland dynamics. Vegetatio 46: 61–74.
Niemelä J. (2002). [Natural regeneration of Scots pine in Savukoski]. Metsähallituksen metsätalouden julkaisuja 42. 41 p. [In Finnish].
Niemistö P., Lappalainen E., Isomäki A. (1993). Growth of Scots pine seed bearers and the development of seedlings during a protracted regeneration period. Folia Forestalia 826. 26 p. [In Finnish with English summary].
Nieppola J. (1992). Long term vegetation changes in stands of Pinus sylvestris in southern Finland. Journal of Vegetation Science 3: 475–484.
Nilsson M.-C. (1994). Separation of allelopathy and resource competition by the boreal dwarf shrub Empetrum hermaphroditum Hagerup. Oecologia 98(1): 1–7.
Nilsson M.-C., Wardle D.A. (2005). Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment 3(8): 421–428.
Nilsson M.-C., Zackrisson O. (1992). Inhibition of Scots pine seedling establishment by Empetrum hermaphroditum. Journal of Chemical Ecology 18(10): 1857–1870.
Nilsson M.-C., Wardle D.A., Dahlberg A. (1999). Effects of plant litter species composition and diversity on the boreal forest plant-soil system. Oikos 86(1): 16–26.
Norberg G., Dolling A., Jäderlund A., Nilsson M.-C., Zackrisson O. (2001). Control of heather (Calluna vulgaris (L.) Hull) by steam treatment: Effects on establishment and early growth of Scots pine. New Forests 21: 187–198.
Ojansuu R., Henttonen H. (1983). [Estimation of the local values of monthly mean temperature, effective temperature sum and precipitation sum from the measurements made by the Finnish meteorological Office]. Silva Fennica 17(2): 143–158. [In Finnish].
Oleskog G., Grip H., Bergsten U., Sahlén K. (2000). Seedling emergence of Pinus sylvestris in characterized seedbed substrates under different moisture conditions. Canadian Journal of Forest Research 30: 1766–1777.
R Core Team. (2009). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. www.R-project.org/.
Robinson R.K. (1972). The production by roots of Calluna vulgaris of a factor inhibitory to growth of some mycorrhizal fungi. Journal of Ecology 60: 219–224.
Salemaa M. (2001). Calluna vulgaris. [Heather]. In: Reinikainen A., Mäkipää R., Vanha-Majamaa I., Hotanen J.-P. (eds.). Changes in the frequency and abundance of forest and mire plants in Finland since 1950. p. 109–111. [In Finnish with English summary].
Sarvas R. (1949). Seed-tree cutting as a regeneration method in Scots pine forests of Southern Finland. Communicationes Instituti Forestalis Fenniae 37(6). 43 p. [In Finnish with English summary].
Skre O., Wielgolaski F.E., Moe B. (1998). Biomass and chemical composition of common forest plants in response to fire in western Norway. Journal of Vegetation Science 9: 501–510.
Steijlen I., Nilsson M.-C., Zackrisson O. (1995). Seed regeneration of Scots pine in boreal forest stands dominated by lichen and feather moss. Canadian Journal of Forest Research 25(5): 713–723.
Suominen O., Olofsson J. (2000). Impacts of semi-domesticated reindeer on structure of tundra and forest communities in Fennoscandia: a review. Annales Zoologi Fennici 37: 233–249.
Väre H., Ohtonen R., Mikkola K. (1996). The effect and extent of heavy grazing by reindeer in oligotrophic pine heaths in northeastern Fennoscandia. Ecography 19: 245–253.
Varmola M., Hyppönen M., Mäkitalo K., Mikkola K., Timonen M. (2004). Forest management and regeneration success in protection forests near the timberline in Finnish Lapland. Scandinavian Journal of Forest Research 19: 424–441.
Wardle D.A., Zackrisson O. (2005). Effects of species and functional group loss on island ecosystem properties. Nature 435: 806–810.
Wardle D.A., Lagerström A., Nilsson M.-C. (2008). Context dependent effects of plant species and functional group loss on vegetation invasibility across an island area gradient. Journal of Ecology 96: 1174–1186.
Wildi O. (2010). Data analysis in vegetation ecology. Wiley-Blackwell, Cornwall. 211 p.
Yang M., Rasbash J., Goldstein H., Barbosa M. (2001). MLwiN Macros for advanced Multilevel modelling. University of Bristol, Centre for Multilevel Modelling. 48 p.
Yli-Vakkuri P. (1961). Experimental studies on the emergence and initial development of tree seedlings in spruce and pine stands. Acta Forestalia Fennica 75(1). 122 p. [In Finnish with English summary].
Zackrisson O., Nilsson M.-C., Steijlen I., Hörnberg G. (1995). Regeneration pulses and climate-vegetation interactions in nonpyrogenic boreal Scots pine stands. Journal of Ecology 83: 469–483.
Zackrisson O., Nilsson M.-C., Dahlberg A., Jäderlund A. (1997). Interference mechanisms in conifer-Ericaceae-feather moss communities. Oikos 78: 209–220.
Total of 61 references
