Teija Ruuhola,
Ari Nikula  ,
Nivala Vesa,
Seppo Nevalainen,
Juho Matala
,
Nivala Vesa,
Seppo Nevalainen,
Juho Matala
Effects of bedrock and surficial deposit composition on moose damage in young forest stands in Finnish Lapland
Ruuhola T., Nikula A., Vesa N., Nevalainen S., Matala J. (2016). Effects of bedrock and surficial deposit composition on moose damage in young forest stands in Finnish Lapland. Silva Fennica vol. 50 no. 3 article id 1565. https://doi.org/10.14214/sf.1565
Highlights
- The effect of bedrock and soil on moose damage in forest plantations were examined
- Moose damage were concentrated in nutrient rich bedrock areas
- Bedrock of damaged stands contained a higher proportion of mafic and alkaline rocks
- Pine-dominated stands on fine grained fertile forest sites had the highest damage risk.
Abstract
There is evidence that moose are attracted to fertile growth habitats apparently due to better quality and larger quantities of food. The nutrients in mineral soils originate from the weathering of bedrock and the composition of parental bedrock affects the fertility of produced mineral soil, thus affecting also the import of nutrients into the whole food web. We surveyed the connection between moose damage in forest plantations and the composition of bedrock and surficial deposits in Finnish Lapland. We used a database of compensated moose damage in private forests in years 1997−2010. Undamaged stands in National Forest Inventories (NFI) from years 1986–2008 served as a control data and moose-damaged NFI-stands as a reference data. Bedrock and surficial depositions and the location of studied stands in relation to ancient shorelines were explored by using the digital databases of the Geological Survey of Finland. Moose-damaged stands were concentrated in southwestern and east Lapland in the areas of the Peräpohja Schist Belt and Lapland’s Greenstone Belt that are both composed of nutrient-rich rocks. The bedrock of damaged stands contained a higher proportion of mafic and alkaline rocks than did the control stands. Moose-damaged stands were pine-dominated and grew in more fertile forest sites than did control stands. Part of pine stands probably located in soils formerly occupied by spruce, which may increase the stands’ vulnerability to biotic threats. Especially, there were relatively more moose damage in pine plantations regenerated on fine-grained mineral soils derived from nutrient rich rocks than in less fertile soils.
Keywords
Pinus sylvestris;
plantation;
Alces alces;
soil;
damage risk;
topography;
geochemistry
- Ruuhola, Natural Resources Institute Finland (Luke), Natural Resources and Bioproduction, Yliopistokatu 6, FI-80101 Joensuu, Finland; University of Eastern Finland, Faculty of Science and Forestry, P.O. Box 111, FI-80101 Joensuu, Finland E-mail teija.ruuhola@uef.fi
-
Nikula,
Natural Resources Institute Finland (Luke), Economics and Society, Eteläranta 55, FI-96300 Rovaniemi, Finland
E-mail
ari.nikula@luke.fi
- Vesa, Natural Resources Institute Finland (Luke), Economics and Society, Eteläranta 55, FI-96300 Rovaniemi, Finland E-mail vesa.nivala@luke.fi
- Nevalainen, Natural Resources Institute Finland (Luke), Natural Resources and Bioproduction, Yliopistokatu 6, FI-80101 Joensuu, Finland E-mail seppo.nevalainen@luke.fi
- Matala, Natural Resources Institute Finland (Luke), Natural Resources and Bioproduction, Yliopistokatu 6, FI-80101 Joensuu, Finland E-mail juho.matala@luke.fi
Received 10 February 2016 Accepted 30 May 2016 Published 23 June 2016
Views 175622
Available at https://doi.org/10.14214/sf.1565 | Download PDF

1 Introduction
Moose (Alces alces L.) populations and consequently moose damage have been at a high level in Fennoscandia for the last decades (see e.g. Markgren 1970; Lavsund 1987; Bergqvist et al. 2014). For example, National Forest Inventory (NFI) shows moose damage in almost one million hectares in Finland (Korhonen et al. 2013). In the most problematic areas, repeated browsing is able to totally prevent the regeneration of forests and in less damaged areas, moose browsing may impair the growth of trees and reduce the quality of sawn timber (Lavsund 1987; Heikkilä and Härkönen 1993; Bergqvist et al. 2003).
Moose damage mostly Scots pine (Pinus sylvestris L.) and birches (Betula spp.), although these are only moderately palatable forage species for the moose. More palatable species like aspen (Populus tremula L.), rowan (Sorbus aucuparia L.), willows (Salix spp.) and juniper (Juniperus communis L.) are also browsed regularly but the large-scale utilization of Scots pine in the regeneration of clear cuts has made it abundantly available and thus the most important food plant for the moose in winter time. Norway spruce (Picea abies L.) is the least palatable forage species and it is browsed less frequently than other tree species in Sweden and Finland (Hörnberg 2001; Månsson et al. 2007).
In fall, part of moose population migrate to winter pastures, which are characterized by a mosaic of young and older forests (Nikula et al. 2004). Stands of an age class less than 30 years are usually preferred by moose (Månsson 2009) and especially pine-dominated young stands seem to characterize moose winter pastures (Nikula et al. 2004). Due to aggregation of several individual moose in the same winter pasture areas, the local density of the moose population can be high and since moose often use the same winter pasture for several consecutive years, moose damage in winter pasture forests can be substantial (Heikkilä and Härkönen 1993).
Resource selection in animals operates on various spatial and temporal scales (Johnson 1980; Senft et al. 1987), and thus an animal’s decision to use a resource, e.g. food patch, may depend on both the availability of food within the patch and the spatial arrangement of food patches at broader spatial scales, such as an animal’s home range. Earlier studies have indicated that resource selection by moose also occurs at several levels of hierarchy in the following order: a landscape (Nikula et al. 2004), home range (Cassing et al. 2006), food patch and food plant (Berqvist et al. 2012; Nikula et al. 2008), although it has been suggested that the food selection of moose mainly occurs at the food plant level (Danell et al. 1991; Vehviläinen and Koricheva 2006; Berqvist et al. 2012). From the point of view of moose damage risk, multi-scale resource selection suggests that underlying factors that affect browsing risk at the plantation or plant level may originate from larger scale processes or factors and should thus be taken into account in the moose damage risk assessment. Moose damage risk is mostly affected by the density of the moose population but also many other factors affect the damage risk (Heikkilä and Härkönen 1993). Vulnerability of saplings to moose damage has been studied, for example, in relation to the chemical properties of the plants (Löyttyniemi 1985; Löyttyniemi and Hiltunen 1978; Haukioja et al. 1983; Niemelä and Danell 1988), the genetic origin of the seedlings (Haukioja et al. 1983), structural characters and species composition of stands (Heikkilä 1990; Heikkilä and Härkönen 1993; Vehviläinen and Koricheva 2006), and in relation to wider structural features of the forest landscape (Heikkilä 1990; Heikkilä and Härkönen 1993; Månsson 2009; Nikula et al. 2004, 2008). In addition, habitat fertility has regularly been shown to increase the risk of moose damage (Niemelä and Danell 1988; Heikkilä and Härkönen 1993; Månsson 2009; Nikula et al. 2008).
The fertility of soil and habitat depends on several different factors such as climate, soil type and thickness, composition and grain size of soil particles. In addition, the bedrock beneath the soil affects soil fertility (Kalliola 1973; Sutinen et al. 2011). The bedrock erodes by physical and chemical factors and the weathered material together with organic material constitute the soil. Therefore, essential nutrients for plants, except nitrogen, are originated from the bedrock beneath the soil (Lindroos 2003; Kujansuu and Kejonen 2005).
The mineral composition of bedrock affects the fertility of formed mineral soil (Mälkönen 2003). For example, soil phosphorus (P) is derived from apatite, which is present in most of Finnish rock types. Potassium (K), in turn, is derived from minerals contained by our most common type of rock, granite. Calcium (Ca), which has a major impact on soil fertility, is an abundant mineral in limestone and dolomite, which are found only in local small areas in Finland. The weathering of granite and other acidic rocks produces nutrient-poor, coarse-grained till, whereas the weathering of mafic and ultramafic rocks produces nutrient-rich, fine-grained till (Kalliola 1973; Koljonen and Tanskanen 1992).
Moraine or till is the most common mineral soil in Fennoscandia and it is composed of different sizes of grain particles: from gravelly to silty till (Koljonen and Tanskanen 1992). Fine-grained till is more fertile than coarse-grained till and it has also a better water-retention capacity (Kalliola 1973; Sutinen et al. 2011). The till composition is usually closely related to the bedrock composition of the area, since the rock materials were drifted rather short distances during last glacial period when till was formed (Koljonen and Tanskanen 1992; Kujansuu and Johansson 2005).
Previous studies have indicated that the fertility of the growth site clearly affects the moose damage risk of young forest stands (Niemelä and Danell 1988; Heikkilä and Härkönen 1993; Månsson 2009). However, a more detailed analysis concerning the relation of soil features and moose damage risk is still lacking. In addition, most of the studies have been conducted in southern Fennoscandia (below 64°N), whereas the moose damage problems in northern Fennoscandia have received much less attention (Nikula et al. 2008). Especially serious moose damage has been reported in the southwestern Lapland (Jalkanen et al. 2005; Nikula et al. 2008). This area is known for alkaline, Ca-rich, bedrock and atypically rich vegetation in otherwise barren Lapland (Kalliola 1973).
The aim of the study was to investigate whether the soil and bedrock type in Finnish Lapland is linked to the occurrence of moose damage on a wider scale. Our working hypothesis was that rock type beneath the soil affects the fertility of habitats and thus, also the selection of moose winter pastures: the moose damage could be more intensive in the areas where the bedrock is composed of nutrient-rich rock types. In addition, the elevation of stands and location in relation to subaquatic areas were examined, since also these factors affect the fertility and other characteristics of soil.
2 Materials and methods
2.1 Description of study area
The study area consists of the province of Lapland, North Finland, without the three northernmost municipalities (65°38´N–68°28´N, 23°15´E–30°00´E) (Fig. 1). The climate of Lapland varies from subarctic in the north to continental in the east. Variation in altitude also causes large variation in climatic factors (Tikkanen 1987). The average annual temperature in the study area varied from +1 °C to −2 °C and the annual precipitation from 500 to 700 mm in years 1981−2010 (Finnish Meteorological Institute 2014). Southwest (SW) Lapland belongs to the mid-boreal zone and northern parts of Lapland to the northern boreal vegetation zone (Kalliola 1973). In our study area about one third of the forestry land is privately owned (Finnish Statistical Yearbook of Forestry 2009).
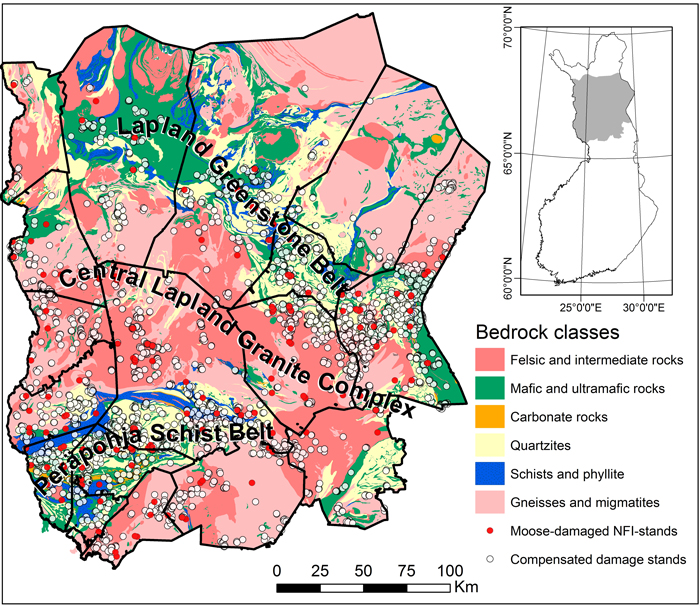
Fig. 1. Bedrock map (a) of the study area in Finnish Lapland, generalized from the Digital bedrock map (1:200 000©, Geological Survey of Finland). Individual rocks were robustly classified to six groups according to their origin and features (see Koljonen 1992). Gneisses and migmatites are grouped into one class independent of their origin. Three major lithological provinces exist in the study area: Lapland Greenstone Belt (LGB), Peräpohja Schist Belt (PSB) and Central Lapland Granitoid Complex (CLGC). Compensated moose damage stands are marked on the map with white symbols and moose-damaged stands investigated in the National Forest Inventory with red labels. Basemap©, the National Land Survey of Finland.
The bedrock of northern Finland is mainly composed of 3100−2500 million-year-old Archean rocks and 1930−1800 million-year-old Paleoproterozoic rocks (Lehtinen et al. 1998). A major part of Finnish bedrock is composed of felsic granitoides (53%) and migmatites (22%) and only a minor part is composed of nutrient-rich, basic rocks (Lehtinen et al. 1998). The bedrock composition of Finnish Lapland is, however, very variable consisting of nutrient-rich rocks like mafic and ultramafic vulcanites, schists and Ca-rich dolomite and limestones in addition to more common felsic granitoides, migmatites and granulites (Pulkkinen and Sarala 2009; Fig. 1). Three major lithological provinces exist in the study area: Lapland Greenstone Belt (LGB), Peräpohja Schist Belt (PSB) and Central Lapland Granitoid Complex (CLGC) (Fig. 1). Non-stratified glacial till or moraine is the most common mineral soil in Lapland; drumlins, hummocky moraine, eskers and other glacial formations and postglacial deposits are also typical features of the landscape (Tikkanen 1987).
2.2 Data of compensated moose damage stands
In Finland, private forest-owners have a possibility to apply compensation fees for financial losses caused by moose browsing in their forest stands (Finlex 2014). Compensation can be applied for the value loss due to a decrease in wood quality and production and also for the costs for reforestation of moose-damaged stands. The damaged area needs to be at least 0.1 hectare and the estimated total loss to the forest owner has to be at least 170 euros. Moose damage evaluation is conducted by an official of the Finnish Forest Centre (Finlex 2014), and during the evaluation, the exact location of the damaged stand, main tree species, the number of other tree species and numerous other variables are recorded. The data of compensated moose damage stands (later referred to as compensated damage stands) from Finnish Lapland between years 1997−2010 were used in this study.
2.3 National Forest Inventory
The data produced by National Forest Inventory (NFI) is based on extensive field work and the NFI-plots are in clusters that form a regular network throughout Finland (Metsäntutkimuslaitos 2002; Nevalainen 1999; Nuutinen et al. 2000). In this study, the forest plots of three national forest inventories from years 1989−1994, 1996−2003 and 2004−2008 in the area of the Finnish Lapland were used as control data. These plots form an extensive network through the study area providing statistically reliable information about the occurrence of moose damage in privately owned land in different parts of the study area. Tree development classes of 2−4, i.e. young seedling stands, advanced seedling stands and young thinning stands (Metsäntutkimuslaitos 2002) were selected to represent stands potentially vulnerable to moose damage. In young seedling stands, the height of the dominant tree storey is less than 1.3 meters. In advanced seedling stands dominant tree storey is more than 1.3 m and for a major part of the dominant trees the diameter at breast height must be less than 8 cm, and for the largest trees, less than 10 cm. In young thinning stands the major part of cutting removal should be pulp wood-sized (Metsäntutkimuslaitos 2002). Except for using only development classes 2–4, we did not apply any other selection criteria to NFI data.
2.4 Lithological and superficial deposit maps and classification
The data for the bedrock and soil composition survey were obtained from two databases: the Digital bedrock map 1:200 000© (Bedrock of Finland 2014 – DigiKP, Geological Survey of Finland (GSF), version 1.0.) and Superficial deposit map of Finland 1:200 000© (Superficial deposits of Finland 2013 – DigiMP, GSF, version 1.0). For the survey of the location of studied stands in relation to subaquatic and supra-aquatic areas, the Ancient Shoreline Database© (ASD; GSF, version 6.1) was used (see details Ojala et al. 2013). For the survey of the elevation of control and compensated damage stands, the Topographic database© of National Land Survey of Finland (NLS) was used. The positional accuracy of the database corresponds to that of scales 1:5000 − 1:10 000 (National Land Survey of Finland 2014).
The individual rock types were robustly classified into six larger groups according to their origin and features (Fig. 1; see Koljonen 1992a,b): 1) Carbonate rocks (limestone, dolomitic carbonate rock, carbonatite, calcsilicate-rock and skarn) 2) Mafic and ultramafic rocks (e.g. gabbro class rocks, mafic tuff, tholeiitic basalt, peridotic komatiite, peridotite) 3) Schist and phyllite-class (graphite, graphite sulphide and biotite paraschist, black schist, phyllite, semipelite) 4) Felsic and intermediate rocks (e.g. granite, dacite, diorite, granophyre, tonalite, felsic tuff) 5) Gneiss class rocks (e.g. migmatitic paragneiss, tonalitic migmatite, garnet-cordierite gneiss, quartz feldspar paragneiss, arkose gneiss) and 6) Quartzite class rocks (arkose quartzite, orthoquartzite, sericite quartzite). Gneisses and migmatites were grouped to one class independent of their origin (Koljonen 1992b). First three classes can be considered as “fertile” rock types and three last ones as “non-fertile” rock types according to their effects on the fertility of the growth sites (Kalliola 1973).
Surface and subsoil types were reclassified into five larger classes 1) Fine-grained stratified soil (mainly silt) 2) Coarse-grained stratified soil (from fine sand to gravel) 3) Rocky soil (bedrock openings, block fields, cobbles and boulders 4) Non-stratified soil (till) 5) Peat (thin and thick peat formations).
2.5 Study setup
NFI-stands without moose damage (later NFI-control stands) served as a control for the compensated damage stands on private land. The number of NFI-control stands was 4551 and the number of compensated stands was 5362. In addition, there were 279 plots in NFI-data with visible moose damage (later NFI-damage stands) and they were used as reference data for the compensated damage stands to ensure that the differences in the treatments were not due to different sampling methods (NFI vs compensated moose damage data). Since compensated damage stands were located on privately owned forest land, only NFI-stands, which were located on private land, were included in the study. We made this restriction since it was expected that the private land would be more fertile than that of state’s land due to the settling history of the Lapland (Järvitalo and Erävuori 2011).
There was a single co-ordinate position for each stand both in NFI and compensated moose damage databases and we assessed attributes of soil and bedrock for each stand by simple overlay analysis of NFI- and compensated damage data with bedrock and soil map data. Although, our data includes only one data point per stand, the extensive number of stands provides a representative overview of the composition of bedrock and superficial deposits in the study area. The altitude of the stands and the location of stands in relation to subaquatic and supra-aquatic areas were assessed similarly for each stand. All the overlay analysis and GIS-data handling were conducted with the ArcMap© (10.1, ESRI) program.
Bedrock composition of the whole private land area was used as reference data for the control stands for checking the validity of NFI-data to represent the general feature of the bedrock and soil composition of the study area. However, this difference was not statistically tested due to different type of the data: whole private land was an aerial data (m2) and NFI-data was a point data.
2.6 Statistics and calculations
To account for the possible regional variation in the amount of private land and forest age we calculated the number of damage per 1000 ha of development classes 2–4 and the proportion of damage area per these, per each municipality (Fig. 2). An area of development classes was calculated from satellite image-based, multi-source national forest inventory data (Tomppo et al. 2008), which corresponds to year 2005. We also wanted to find out whether the number and area of damage were dependent on the available amount of development classes 2–4 and, therefore, we also calculated correlations between these. Correlation tests were conducted with the Spearman’s non-parametric correlation test (SPSS 20.0).
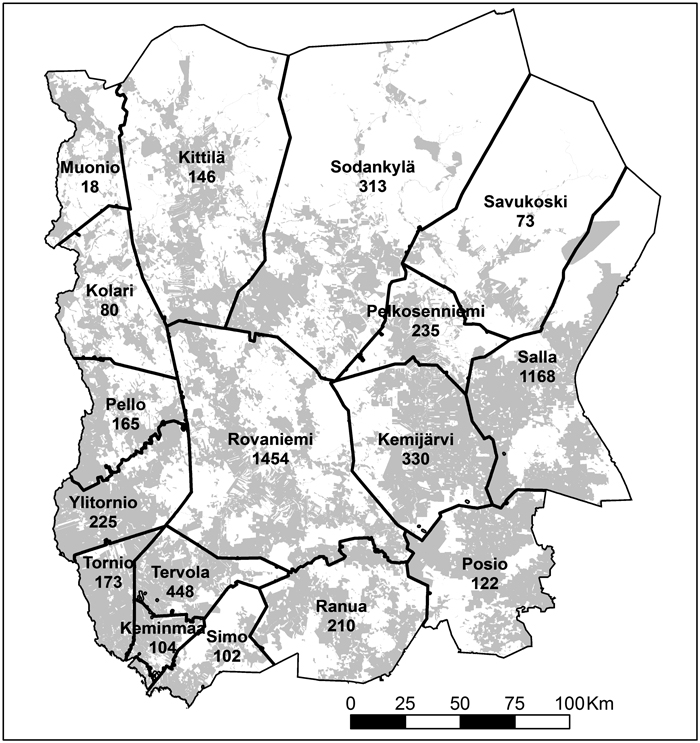
Fig. 2. Municipalities, the number of compensated damage stands in each municipality and private land (grey) in the study area.
Statistical differences in the distributions of different rock and soil classes, as well as forest types between NFI-control stands, NFI-damage stands and compensated damage stands were tested with the crosstabs function of SPSS (20.0) by using the Pearson’s Chi-square test for overall differences and z-test for column (dependent) proportions. The difference in altitudes between control and compensated damage stands were tested with the t-test for the equality of means (SPSS 20.0).
3 Results
3.1 The number and area of compensated moose damage stands in the municipalities of Lapland
The private forest-owners of Finnish Lapland submitted ca. 5400 applications for moose damage compensations between years 1997−2010, which represent a damage area of about 18 500 hectares. The largest absolute number of compensations were paid for and damage area located in two municipalities: Rovaniemi and Salla; about half of the compensations in Lapland were paid in these municipalities. When the number of compensated damage stands was calculated by 1000 ha of young stands in each municipality, the largest number of compensated damage stands were found in the municipalities of Tervola (9.93), Rovaniemi (8.8), Salla (7.71), Pelkosenniemi (7.15), Keminmaa (6.69) and Tornio (4.48), i.e. in eastern and south-western Lapland. The lowest number of the compensations, in turn, were paid in the municipalities of Muonio (0.89), Posio (1.43), Kittilä (1.73) and Kolari (1.83), i.e. mainly in western and south-eastern Lapland (Fig. 2). The largest proportion of moose-damaged stands of the area of young stands were found in the municipalities of Salla (3.9%), Pelkosenniemi (2.9%), Rovaniemi (2.45%) and Tervola (2.41%), and the smallest relative damage areas were found in Muonio (0.29%), Posio (0.42%) and Savukoski (0.67%) (Data not shown).
There were no significant correlations between neither the number of moose damage per 1000 ha of development classes 2–4 and the proportion of development classes 2–4 of private land (Spearman’s rho = 0.221, p = 0.395, n = 17) nor with the proportion of compensated area of development classes 2–4 and the proportion of development classes 2–4 of private land (Spearman’s rho = 0.267, p = 0.300, n = 17).
3.2 Forest type and the main tree species of compensated and NFI-stands
The main forest site types of all stands studied were mesic and sub-xeric heath forests: 84–98% of stands belonged to these two types (Table 1). The proportion of mesic heath was higher than that of sub-xeric heath in all stand groups. The proportion of mesic heath was the lowest in the NFI-control stands (44%), second highest in the compensated damage stands (54%) and the highest in NFI-damage stands (62%). The proportion of sub-xeric heaths, in turn, was higher in compensated damage stands than in either of the NFI-stand groups. The overall distribution of the forest types significantly differed between NFI-control and compensated damage stands (χ2 = 630; P < 0.001); between NFI-control and NFI-damage stands (χ2 = 35; P < 0.001) and between compensated damage and NFI-damage stands (χ2 = 655; P < 0.001). Overall, there were more both the most nutrient-rich as wells as nutrient-poor site types in NFI-damage and NFI-control stands than there were in compensated damage stands (Table 1).
| Table 1. The proportions of forest site types and the main tree species among compensated moose damage stands and NFI-stands in Finnish Lapland. | |||
| Forest type | Control NFI-stands | Compensated stands | Damaged NFI-stands |
| Herb-rich | 1.7a | 0.0b | 1.4a |
| Herb-rich heath | 10.2a | 1.8b | 7.2a |
| Mesic heath | 44.4a | 54b | 62.4c |
| Sub-xeric heath | 39.3a | 43.8b | 25c |
| Xeric heath | 4.2a | 0.4b | 3.6a |
| Barren heath | 0.2a | 0.0b | 0.4a |
| Main tree species | |||
| Scots pine | 77.3a | 99.2b | 86c |
| Norway spruce | 10.7a | 0.4b | 7.5a |
| Birch (Betula sp.) | 11.9a | 0.0b | 4.3c |
| Aspen | 0.0a | 0.3b | 2.2c |
| Alder (Alnus sp.) | 0.0a | 0.1a | 0.0a |
| Other | 0.1a | 0.0a | 0.0a |
| a,b,c Statistically significant differences between treatment groups. | |||
The major tree species in compensated damage stands was Scots pine: over 99% of stands were pine-dominated (Table 1). Scots pine was also the dominant tree species in the majority of NFI-control (77%) and NFI-damage stands (86%), but they contained higher proportions of spruce- and birch-dominated stands than did the compensated damage stands (Table 1). The proportions of a stand’s main tree species significantly differed between NFI-control and compensated damage stands (χ2 = 1298; P < 0.001); NFI-control and NFI-damage stands (χ2 = 89; P < 0.001) and between compensated damage and NFI-damage stands (χ2 = 433; P < 0.001).
3.3 Bedrock composition in compensated and NFI-stands
When compared to bedrock composition on private land, the distribution of different rock classes in NFI-control stands followed well the bedrock composition of the study area (Fig. 3). On contrast, the bedrock composition of compensated damage stands significantly differed (χ2 = 655; P < 0.001) from that of NFI-control stands (Fig. 3). Multiple comparisons showed that the moose-damaged stands had significantly larger proportions of carbonate rocks, mafic and ultramafic rocks and smaller proportions of gneiss class rocks and felsic and intermediate rocks than the control stands (Fig. 3). Interestingly, the proportion of felsic quartzite class rocks was also a significantly higher in the compensated damage stands than in the control stands, whereas the schist and phyllite-class rocks was the only rock class that did not differ between NFI-control and compensated damage stands (Fig. 3).
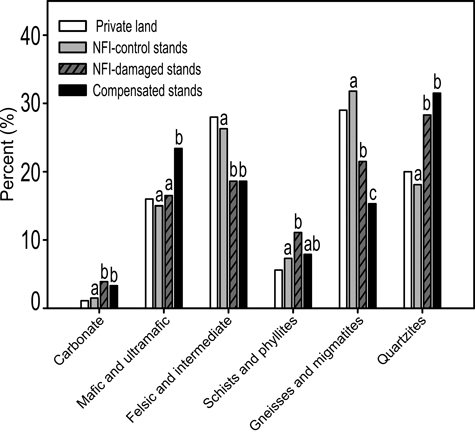
Fig. 3. Proportion of different rock classes in private land, NFI-control stands, NFI-damage stands and compensated moose damage stands. Statistically significant (P ≤ 0.05) differences are marked with different letters. The proportions of private land were not tested but they were used as a control for the proportions of NFI-control stands.
NFI-damage stands had a more similar bedrock composition with compensated damage stands than with NFI-control stands (Fig. 3). NFI-damage stands had, however, a higher proportion of gneiss-class rocks than the compensated damage stands, which, in turn, had a significantly higher proportion of mafic and ultramafic rocks (Fig. 3). Otherwise, differences to NFI-control plots showed a similar pattern with compensated stands, although all the differences were not significant (Fig. 3).
When the bedrock types were grouped into two classes (excluding quartzites), nutrient-rich (carbonate, mafic and ultramafic, schists and phyllite) and nutrient-poor rocks (felsic and intermediate, gneisses and migmatites), the proportion of nutrient-rich rocks was markedly higher in compensated damage stands than in the NFI-control stands (χ2 = 623; P < 0.001), whereas the proportion of nutrient-poor rocks was smaller in compensated damage stands (Fig. 3). This difference was found for all forest site types, which also significantly (χ2 = 128; P < 0.001) differed from each other in their proportions of nutrient-rich and nutrient-poor rocks (Fig. 4).
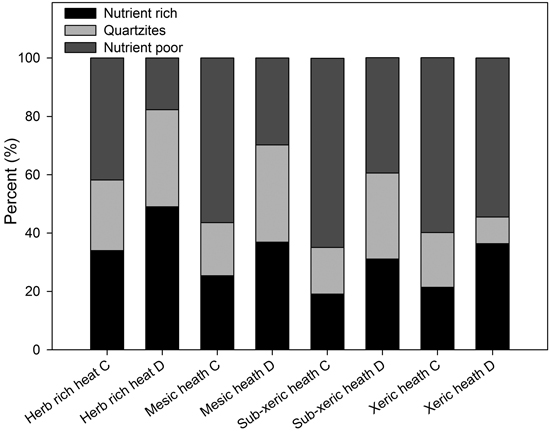
Fig. 4. The proportions of nutrient-rich (carbonate, mafic and ultramafic rocks, schists and phyllite), nutrient-poor (felsic and intermediate rocks, gneisses and migmatites) and quartzites in different site types representing different fertility class and moose damage status (C = NFI-control stands, D = compensated moose damage stands).
The proportion of nutrient-rich rocks showed a decreasing tendency with a decrease in the fertility of the site type both in the NFI-control stands and compensated damage stands (Fig. 4). Oppositely, the proportion of nutrient-poor, acidic rocks increased when the fertility of the site type decreased (Fig. 4). Interestingly, the proportion of quartzite rocks also decreased with the fertility of site type and this decrease was clearer in the damaged stands than in the control stands (Fig. 4).
3.4 Composition of superficial deposits
The proportions of superficial deposits significantly differed between NFI-control stands and compensated damage stands (surface soil: χ2 = 359; P < 0.001; subsoil: χ2 = 313; P < 0.001) but not between NFI-control stands and NFI-damage stands; thus, damaged NFI-stands were not examined in more detail. The proportion of till was significantly higher in the compensated damage stands than in the control stands (Table 2). In contrast, the proportions of grained soils, peatlands and rocky soils (boulder fields, bedrock openings) were significantly lower in the compensated damage stands than in the control stands (Table 2). Similar results were detected in the case of subsoil composition (Table 2).
| Table 2. Distribution of superficial deposits in undamaged NFI-stands (control) and compensated moose damage stands in Finnish Lapland. | ||||
| Proportion of superficial deposits (%) | ||||
| Surface soil | Subsoil | |||
| Soil type | NFI-Control stands | Compensated damage stands | NFI-Control stands | Compensated damage stands |
| Fine-grained stratified | 1.2a | 0.5b | 1.8a | 0.6b |
| Coarse-grained stratified | 10.4a | 6.5b | 10.8a | 6.8b |
| Rocky | 4.0a | 2.2b | 4.0a | 2.1b |
| Non-stratified | 63.7a | 81.5b | 70.3a | 85.2b |
| Peat deposits | 20.6a | 9.3b | 13.1b | 5.2b |
| a,b Statistically significant differences between treatment groups. | ||||
3.5 Stand location in relation to altitude and subaquatic and supra-aquatic areas
Significantly (χ2 = 220; P < 0.001) more compensated damage stands (82.5%) than NFI-control stands (70.3%) were located in the so-called sub-aquatic area formerly under the ancient phases of the Baltic Sea or under northern ice lakes after the last deglaciation ca. 7000−8000 years ago (Fig. 5). Also, a significantly (χ2 = 9,317; P = 0.002) larger part of NFI-damage stands were located in the subaquatic area than NFI-control stands, whereas the difference compared to compensated damage stands was non-significant (χ2 = 2.870; P = 0.090). In northern and NE Lapland compensated damage stands were located in a lower elevation than the NFI-control stands but in SW Lapland the situation was the opposite (Fig. 6).
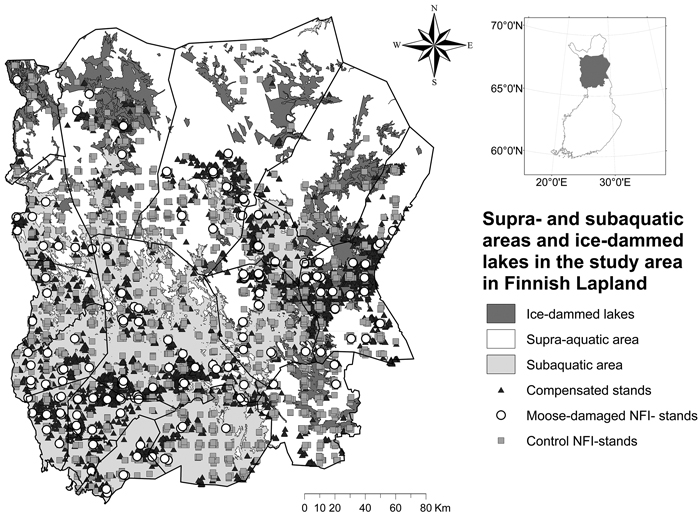
Fig. 5. Location of compensated moose damage, NFI-control and NFI-damage stands in relation to the location of subaquatic and supra-aquatic areas in Finnish Lapland. Subaquatic areas represent the highest shoreline ca. 7000–9000 BP (Ojala et al. 2013). Ancient Shoreline Database©, Geological Survey of Finland; Basemap©, the National Land Survey of Finland.
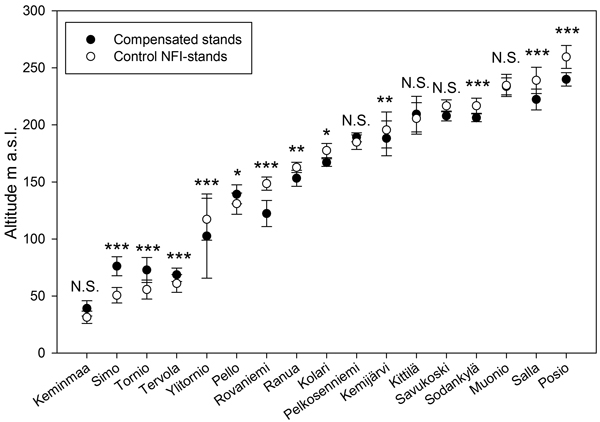
Fig. 6. The average altitudes (m a.s.l.) and S.D. of compensated moose damage and NFI-control stands in the municipalities (see Fig. 1) of the study area in Finnish Lapland. Statistical significances between groups: *** = p < 0.001, ** = p < 0.01, * = p < 0.05 and N.S = p > 0.05.
4 Discussion
4.1 Largest number and area of moose damage stands located in SW and eastern Lapland
Moose damage in the private forests in Finnish Lapland comprised a total area of 18 500 ha; altogether 5400 moose damage reports were registered in years 1997−2010. The majority of these damage stands were located in SW Lapland (Tervola, Keminmaa, Tornio, Rovaniemi) and eastern Lapland (Salla, Pelkosenniemi), whereas the proportion of moose damage was the smallest in the municipalities of Savukoski and Muonio. The occurrence of damage generally coincide with the study of Jalkanen et al. (2005).
SW Lapland is renowned for its alkaline, Ca-rich, bedrock and rich, often calcifuge vegetation (Kalliola 1973). The bedrock of SW Lapland is mainly composed of basic and mafic rocks of the Peräpohja Schist Belt (PSB) (Fig. 1; Noukka 1987; Silvennoinen 1998) but some dolomite (Ca-Mg-carbonate) deposits also exist in the area (Fig. 1; Noukka 1987). In the municipality of Rovaniemi moose damage were concentrated to southern parts that belong to PSB, whereas there were less damage in the northern part which is composed of nutrient-poor felsic granitoides of the Central Lapland Granitoid Complex (CLGC) (Noukka 1987) (Fig. 1).
The municipalities of Pelkosenniemi and Salla, in turn, are located within the precincts of the Lapland Greenstone Belt, LGB (Fig. 1; Noukka 1987). The bedrock of Pelkosenniemi is entirely composed of LGB rocks, whereas the bedrock of Salla is partly composed of mafic rocks of LGB and partly of felsic granitoides of CLGC (Fig. 1; Noukka 1987). Dolomite deposits also exist in the eastern part of Salla. In contrast, in the areas of low moose damage the bedrock is largely composed of nutrient-poor felsic granites, like in Savukoski and felsic granitoides and gneisses but also quartzites and mafic volcanites, like in Muonio (Fig. 1.; Noukka 1987).
4.2 Moose damage was concentrated in the areas of nutrient-rich rock types
The bedrock of compensated damage stands and NFI-damage stands contained significantly higher proportions of nutrient-rich rocks; carbonate rocks (mainly dolomite), mafic and ultramafic rocks, than did the control stands. Evidently, the composition of bedrock also affected the fertility of habitats, since the proportion of nutrient-rich rock types were the highest in herb-rich heath forests gradually decreasing with the decrease in the fertility of forest type. Intriguingly, the proportion of nutrient-rich rocks was clearly higher in compensated moose damage stands than in control stands in all forest site types, reflecting the variation in the fertility within the forest site type (see Hotanen et al. 2008).
It is well-known that the composition of the parental bedrock has a determinative role in the composition of the aboveground vegetation (e.g. Kalliola 1973; Sutinen et al. 2002, 2012; Närhi et al. 2011) and the concentrations of plant elements generally reflect the element composition of the bedrock beneath the growing site (Närhi et al. 2013). It has been shown that the forest vegetation communities of Finnish Lapland are intimately related to soil chemistry; Ca-concentration being the major variable in determining plant diversity (Närhi et al. 2011). For example, herb-rich forests grow in alkaline soils originated from Ca-rich rocks like dolomite and greenstones and many demanding calcifuge species colonize these fertile herb-rich forests (Kalliola 1973).
The Lapland greenstones are composed of Mg-, Fe- and Ca-rich minerals (Koljonen 1992b) and till that has originated from the rocks of greenbelt stones, is nutrient-rich. Nutrients are also available to plants due to easy dissolving of mafic minerals (Koljonen 1992c; Sutinen et al. 2002, 2011a,b). Till that has been formed from Svekokarelian schists and gneisses has lower level of nutrients than greenstones but the minerals are also available for plants due to the fine-grained mineral composition of rocks and easy leaching of nutrients with water (Koljonen 1992b). Oppositely, the concentration of nutrients in glacial till formed from granitoids of Central Lapland (CLGC) and Archean gneisses is so low that nutritional deficiencies of many elements are widespread in these areas (Koljonen 1992c).
Quartzites are mostly nutrient-poor rocks (e.g. Kalliola 1973). However, quartzites of the study area are stratified with Svekokarelian schists and gneisses containing, e.g. Ca- and Mg-rich dolomites and graphite-rich schists (Koljonen 1992b), which explains why otherwise nutrient-poor quartzites acted similarly than nutrient-rich rock types: their proportion was highest in fertile site types and decreased with the fertility of the site type. Their proportion was also higher in moose-damaged stands than in the control stands. In addition, it has been suggested that geological nitrogen (N) bound in sedimentary, metasedimentary and metamorphic rocks such as sand stones, schists and quartzites, may comprise a considerable proportion of total N budget in arctic and subarctic areas, wherein N deficiency invariably limits the growth of plants due to the climatic and soil constrains (Dixon et al. 2012).
4.3 Moose damage was the highest in pine-dominated mesic and sub-xeric heath forests
The main tree species in the compensated damage stands was Scots pine (99%) and the most common (98%) forest types were mesic and sub-xeric heath forests. The proportion of mesic forest type was higher among moose-damaged stands (both compensated, 54% and NFI-damage stands, 62% ) than among controls stands (44%), which may either, reflect the active food selection of moose or be simply due to forest owners’ higher interest in growing pine on more productive site types. The economic value of the fast growing pine stands of fertile habitats is much higher than the value of the stands growing in less fertile growth sites. Thus, also a threshold to follow the condition of the economically valuable stands and readiness to report on the damage might be lower than in the case of economically less important stands. However, it is well-documented that moose prefer to forage on young pine-dominated stands growing in fertile habitats (e.g. Niemelä and Danell 1988; Heikkilä 1990; Heikkilä and Härkönen 1993; Ball and Dahlgren 2002; Månsson 2009; Nikula et al. 2008). Fertile habitats offer moose not only better quality and a larger quantity of food but also a variety of alternative food plants (Niemelä and Danell 1988; Heikkilä 1990; Heikkilä and Härkönen 1993; Månsson 2009; Nikula et al. 2008). Pine stands mixed with deciduous seedlings seem to be attractive pastures for moose (Heikkilä 1990; Vehviläinen and Koricheva 2006), especially when birches are overtopping the pine seedlings (Nikula et al. 2008).
In the study of Jalkanen et al. (2005), the moose damage in cultivated pine stands was the most serious in mesic heath composed of fine-sorted mineral soil and in drained peatland, when all moose browsing, also non-detrimental was counted. In contrast, Nikula et al. (2008) reported that the soil type had no significant effect on damage risk, except that the paludification of soil decreased the risk for the moose-damage, when only the broken main shoot was counted as moose damage. In our study, the major portion of the surficial deposits of NFI-control and compensated moose damage stands was composed of non-stratified till, which was expected since over 75% of mineral soil in northern Scandinavia is composed of glacial till (Sutinen et al. 2007). The proportion of till-covered soil was, however, significantly higher among compensated damage stands (79%) than among undamaged stands (62%), whereas the proportions of stratified soils, peatlands, boulder fields and bedrock openings were lower in compensated damage stands.
To conclude, our results indicate that the risk for moose damage is high in pine plantations growing on fertile soils and especially over nutrient-rich bedrock. Because bedrock type can vary also within each site type this can partly obscure the effect of site type but also soil type on moose damage risk, which might be an explanation for discrepancies among earlier studies.
4.4 Are moose-damaged stands located on suboptimal growth sites?
In Lapland, a clear difference exists in the natural dispersal of Scots pine and Norway spruce: Norway spruce inhabit areas of fine-grained, Mg- and Ca-rich till that is derived from the weathering of mafic rocks, whereas Scots pine typically grows in coarse-grained, dry, acidic and nutrient-poor soil that is derived from felsic granitoides (Sutinen et al. 2002; 2011a,b). It has been suggested that low soil moisture is a prerequisite for the growth of Scots pine (Sutinen et al. 2002) and the reforestation of previously spruce-dominated fine-grained, moist habitats with pine has often failed (Sutinen et al. 2007). In addition, pines planted in mesic sites often suffer from high incidences of fungal diseases, e.g. those caused by Gremmeniella abietina (Lagerb.) Morelet (Sutinen et al. 2002).
Scots pine is the most important tree species from an economic perspective due to its high timber value and productivity. Thus, it has been largely used in the reforestation of clear cuts in Finland, also in sites that were formerly spruce-dominated (Sutinen et al. 2002, 2007). In our study, over half of the moose-damaged stands located on moist heaths and in sub-xeric heaths and further in the most fertile parts of these forest types, if bedrock composition is pored over (Fig. 5). Thus, a portion of compensated damage stands are probably located in areas formerly occupied by Norway spruce, i.e. in suboptimal growth site for Scots pine (Sutinen et al. 2007). Heavy mechanical site preparation (MSP) methods like ploughing have been often used to improve the performance of young pine saplings in formerly spruce-occupied sites and the positive effect of MSP on the height growth of Scots pine has been found to continue throughout the time that the leader shoots of pine are available for moose (Mäkitalo 1999). However, MSP methods expose soil minerals to excess leaching (especially P, Ca and Mg) and the soil-drainage effects of MSP also start to vanish after some 12–20 years in Finnish Lapland, depending on the original water content of the soil (Sutinen et al. 2002, 2005, 2007). Thus, pines planted in too moist and fine-grained soil may suffer from nutrient stress and/or imbalance in addition to excess moisture, especially in the advanced stages of the plantation; thus their resistance to plant-eating animals might also be lowered (Phelan et al. 1996). Intriguingly, it has been shown that heavy soil scarification is associated with an increased moose damage probability compared to lighter scarification methods (Nikula et al. 2008).
The use of proportional damage as a measure of the degree of damage might at least partly obscure the interpretation of the effect of regeneration methods on moose damage risk. For example, the moose damage level in planted pine stands has been reported to be markedly higher (20.8%) than in the seeded stands (8.4%) in northern Finland (Jalkanen et al. 2005). Planting methods may also explain the concentration of moose damage in fertile sites. The recommended regeneration methods for Scots pine in moist and fine-grained soils are planting of artificially cultivated saplings instead of seeding. However, the density of the saplings remains much lower in the planted stands than in seeded stands and thus, the proportional level of damage of planted stands also can be much higher than in seeded stands (Heikkilä 2000), although the absolute number of browsed trees may be the same.
4.5 Moose-damaged stands mainly located in subaquatic areas and at low elevation
The different phases of the Baltic Sea basin from 11 600–9000 years BP on are closely related to sedimentary deposits in current land areas (Ojala et al. 2013). In our study, the majority of moose damage stands are located in a so-called subaquatic area, which were either under ancient phases of the Bothnian Sea in SW Lapland or under ice-dammed lakes in Eastern Lapland (Johansson 2005). Most of the area in SW Lapland was under water after the last glacial period, thus part of the results are probabaly just an anomaly. However low-lying terrain of previously subaquatic areas often contains fine-grained material deposited on the surface of non-stratified till (Sarala and Rossi 2006) because the finest particles of soil leached away from fell slopes and were deposited at lower altitudes (Lindroos 2003; Sarala and Rossi 2006). A typical feature for these shore deposits is that under the well-drained surface soil a water-retaining soil layer exists composed of silt, clay or fine-grained till. The vegetation in these areas is atypically fresh for a superficially coarse-grained soil (Lindroos 2003) making these probably favoured habitats by moose.
Compensated damage stands generally were located at lower altitudes than control stands which is in line with Nikula et al. (2008) who found that moose damage were the most serious in lower altitudes. One explanation for this can be that the till in the upper slopes of fells have lower concentrations of Ca and Mg than lower slopes (Sutinen et al. 2012). This partly co-incides with the finding of Nikula et al. (2008) who found that the concentration of soil-exchangeable Mg was a significant variable in the assessment of moose damage risk. In addition, it has been proposed that high Ca- concentration, together with the high protein and P content of food plants, have a decisive role in the food selection of moose (Andersson and Markkula 1974).
5 Conclusions
Compensated moose damage stands located in nutrient-rich habitats, and their bedrock contained significantly higher proportions of carbonate, mafic and ultramafic rocks than control stands. Damaged stands mostly located in SW and eastern Lapland in the area of the Peräpohja Schist Belt and Lapland Greenstone Belt that are composed of Ca- and Mg-rich rocks. Calcium plays a pivotal role in soil fertility by decreasing soil pH, and thus improving the nutrient uptake of plants. Especially, the growth of Scots pine benefits from the high soil Ca concentration (Tamminen and Mälkönen 2003). The moose is known to prefer nutrient-rich sites and it has been suggested that Ca and Mg could positively direct the food selection of the moose. Our results suggest that pine stands growing in Ca- and Mg-rich soils are especially vulnerable to moose damage. In addition, a part of pine stands were probably planted in moist and fertile sites formerly occupied by Norway spruce that are suboptimal habitats for Scots pine, which may suffer from diseases, hypoxia, nutrient imbalance and further a high risk for moose damage. Altogether, our results suggest that fertile habitats in the areas of BPS and LGB are especially vulnerable to moose damage and the use of Norway spruce in the regeneration of clear cuts in these areas could decrease the damage since spruce is the least preferred food plant for the moose and spruce naturally occupies moist and fertile habitats.
Acknowledgements
We thank Olli Lehtonen and Mikko Nivala for the guidance in the use of ArcMap-program and Olli-Pekka Tikkanen for comments concerning the MS. The analyses and writing work was financially supported by Ministry of Employment and the Economy as providing salary for TR (subsidiary work). We also thank two anonymous reviewers for their valuable comments.
References
Andersson E., Markkula A. (1974). Hirven talviravinnon kemiallisesta koostumuksesta. Suomen Riista 25: 15–19.
Ball J.P., Dahlgren J. (2002). Browsing damage on pine (Pinus sylvestris and P. contorta) by a migrating moose (Alces alces) population in winter: Relation to habitat composition and road barriers. Scandinavian Journal of Forest Research 17(5): 427–435. http://dx.doi.org/10.1080/028275802320435441.
Bergqvist G., Bergström R., Edenius L. (2003). Effects of moose (Alces alces) rebrowsing on damage development in young stands of Scots pine (Pinus sylvestris). Forest Ecology and Management 176(1–3): 397–403. http://dx.doi.org/10.1016/S0378-1127(02)00288-8.
Bergqvist G., Bergström R., Wallgren M. (2012). Browsing by large herbivores on Scots pine (Pinus sylvestris) seedlings in mixture with ash (Fraxinus excelsior) on silver birch (Betula pendula). Scandinavian Journal of Forest Research 27(4): 372–378. http://dx.doi.org/10.1080/02827581.2011.635155.
Bergqvist G., Bergström R., Wallgren M. (2014). Recent browsing damage by moose on Scots pine, birch and aspen in young commercial forests – effects of forage availability, moose population density and site productivity. Silva Fennica 48(1): 1−13. http://dx.doi.org/10.14214/sf.1077.
Cassing G., Greenberg L.A., Mikusinski G. (2014). Moose (Alces alces) browsing in young forest stands in central Sweden: a multiscale perspective. Scandinavian Journal of Forest Research 21(3): 221−230. http://dx.doi.org/10.1080/02827580600673535.
Danell K., Niemelä P., Varvikko T., Vuorisalo T. (1991). Moose browsing on Scots pine along a gradient of plant productivity. Ecology 72(5): 624–1633. http://dx.doi.org/10.2307/1940962.
Dixon J.C., Cambell S.W., Durham B. (2012). Geological nitrogen and climate change in the geochemical budget of Käkevagge, Swedish Lapland. Geomorphology 167–168: 70–76. http://dx.doi.org/10.1016/j.geomorph.2012.03.011.
Finlex (2014). Legislation of Finland: Game Animal Damages Act, http://www.finlex.fi/en/laki/kaannokset/2009/en20090105.pdf. [Cited 1 Dec 2014].
Finnish Statistical Yearbook of Forestry (2009). Finnish Forest Research Institute. Vammalan Kirjapaino Oy. Sastamala 2009. 452 p. ISBN 978-951-40-2204-3.
The Finnish Meteorological Institute (2014). Climate: normal period 1981–2010. http://en.ilmatieteenlaitos.fi/normal-period. [Cited 21 Nov 2014].
Haukioja E., Huopalahti R., Kotiaho J., Nygren K. (1983). Millaisia männyntaimia hirvi suosii? Summary: The kinds of pine preferred by moose? Suomen Riista 30: 22–27.
Heikkilä R. (1990). Effect of plantation characteristics on moose browsing on Scots pine. Silva Fennica 24(4): 341–351. http://doi.org/10.14214/sf.a15588.
Heikkilä R. (2000). Männyn istutustaimikoiden metsänhoidollinen tila hirvivahingon jälkeen Etelä-Suomessa. [The silvicultural condition of pine plantation after moose damage in southern Finland]. Metsätieteen aikakauskirja 2: 259–267.
Heikkilä R., Härkönen S. (1993). Moose (Alces alces L.) browsing in young Scots pine stands in relation to the characteristics of their winter habitats. Silva Fennica 27(2): 127–143. http://doi.org/10.14214/sf.a15667.
Hörnberg S. (2001). The relationship between moose (Alces alces) browsing utilization and the occurrance of different forage species in Sweden. Forest Ecology andManagement 149(1–3): 91–102. http://dx.doi.org/10.1016/S0378-1127(00)00547-8.
Hotanen J-P., Nousiainen H, Mäkipää R., Reinikainen A., Tonteri T. (2008). Metsätyypit – opas kasvupaikkojen luokitteluun. [Forest site types – a guide for classifying site types]. Metsäntutkimuslaitos, Metsäkustannus Oy, Hämeenlinna. 192 p. ISBN-13: 978-952-5694-22-2.
Jalkanen R., Aalto T., Hallikainen V., Hyppönen M., Mäkitalo K. (2005). Viljelytaimikoiden hirvituhot Lapissa ja Kuusamossa. [Moose damage in artificially regenerated plantations in Lapland and Kuusamo]. Metsätieteen Aikakausikirja 4: 399–411.
Järvitalo A., Erävuori L. (2011). Länsi-Lapin maakuntakaavan taustaselvitykset: maisema ja luonnonympäristö. [Background surveys of the West-Lapland regional land use plan: landscape and nature]. Lapin Liitto, Sito Oy, Espoo. 106 p.
Johansson P. (2005). Jääjärvet. In: Johansson P. Kujansuu R. (eds.). Pohjois-Suomen maaperä, maaperäkarttojen 1:400 000 selitys. [Summary: Quaternary deposits of Northern Finland, Explanation to the maps of Quaternary deposits 1:400 000]. Vammalan Kirjapaino Oy, Rovaniemi. p. 127–148. ISBN-13: 9789516909052.
Johnson D.H. (1980). The comparison of usage and availability measurements for evaluating resource preference. Ecology 61: 65–71. http://dx.doi.org/10.2307/1937156.
Kalliola R. (1973). Suomen Kasvimaantiede. [Phytogeography of Finland]. Werner Söderström Oy, Porvoo, Helsinki. 308 p. ISBN 978-9510057315.
Koljonen T. (1992a). Geochemical evolution of the Earth. In: Koljonen T. (ed.). The geochemical atlas of Finland, part 2: Till. Geological Survey of Finland, Espoo. p. 29–39. ISBN 951-690-379-7.
Koljonen T. (1992b). Bedrock. In: Koljonen T. (ed.). The geochemical atlas of Finland, part 2: Till. Geological Survey of Finland, Espoo. p. 51–60. ISBN 951-690-379-7.
Koljonen T. (1992c). Geochemical domains, provinces, and subprovinces. In: Koljonen T. (ed.). The geochemical atlas of Finland, part 2: Till. Geological Survey of Finland, Espoo. p. 107–122. ISBN 951-690-379-7.
Koljonen T., Tanskanen H. (1992). Queternary Sediments, In: Koljonen T. (ed.). The geochemical atlas of Finland, part 2: Till. Geological Survey of Finland, Espoo. p. 44–50. ISBN 951-690-379-7.
Korhonen K.T., Ihalainen A., Viiri H., Heikkinen J., Henttonen H.M., Hotanen J.-P., Mäkelä H., Nevalainen S., Pitkänen J. (2013). Suomen metsät 2004–2008 ja niiden kehitys 1921–2008. [Forests of Finland in 2004–2008 and their development in 1921–2008]. Metsätieteen aikakausikirja 3: 269–608.
Kujansuu R., Johansson P. (2005). Moreenikerrostumat. In: Johansson P., Kujansuu R. (eds.). Pohjois-Suomen maaperä, maaperäkarttojen 1:400 000 selitys. Summary: Quaternary deposits of Northern Finland, explanation to the maps of Quaternary deposits 1:400 000. Vammalan Kirjapaino Oy, Rovaniemi. p. 34−50. ISBN-13: 9789516909052.
Kujansuu R., Kejonen A. (2005). Rapautuminen ja sen synnyttämät pinnanmuodot. In: Johansson P., Kujansuu R. (eds.). Pohjois-Suomen maaperä, maaperäkarttojen 1:400 000 selitys. Summary: Quaternary deposits of Northern Finland, explanation to the maps of Quaternary deposits 1:400 000. Vammalan Kirjapaino Oy, Rovaniemi. p. 20−25. ISBN-13: 9789516909052.
Lavsund S. (1987). Moose relationships to forestry in Finland, Norway and Sweden. Swedish Wildlife Research, Supplement 1: 229–244.
Lehtinen M., Nurmi P., Rämö T. (1998). Suomen Kallioperä, 3000 Vuosimiljoonaa. [Bedrock of Finland, 3000 million years]. Suomen Geologinen Seura, Jyväskylä. 375 p. ISBN: 952-90-9260-1.
Lindroos E. (2003). Maaperä. [Soil]. In: Mälkönen E. (ed.). Metsämaa ja sen hoito. [Forest soil and its treatment]. Kustannusosakeyhtiö Metsälehti, Hämeenlinna. p. 7–38. ISBN-13: 9789525118575.
Löyttyniemi K. (1985). On repeated browsing of scots pine saplings by moose (Alces alces). Silva Fennica 19(4): 387–391. http://doi.org/10.14214/sf.a15431.
Löyttyniemi K., Hiltunen R. (1978). Monoterpenes in Scots pine in relation to browsing preference by moose (Alces alces L.). Silva Fennica 12(2): 85–87. http://doi.org/10.14214/sf.a14846.
Mäkitalo K. (1999). Effect of site preparation and reforestation method on survival and height growth of Scots pine. Scandinavian Journal of Forest Research 14(6): 512–525. http://dx.doi.org/10.1080/02827589908540816.
Mälkönen E. (2003). Metsämaan ravinteisuuden hoito. [Treatment of the soil nutrients]. In: Mälkönen E. (ed.). Metsämaa ja sen hoito. [Forest soil and its treatment]. Kustannusosakeyhtiö Metsälehti, Hämeenlinna. p. 175–198. ISBN-13: 9789525118575.
Markgren G. (1974). The moose in Fennoscandia. Le Naturaliste canadien 101: 185–194.
Metsäntutkimuslaitos (2002). Valtakunnan Metsien 9. Inventointi (VMI9). Maastotyön ohjeet 2002. Kuusamo ja Lappi. [9th National Forest Inventory (NFI9). Field work manual 2002. Kuusamo and Lapland]. Metsäntutkimuslaitos, Helsinki. 178 p.
Månsson J. (2009). Environmental variation and moose Alces alces density as determinants of spatio-temporal heterogeneity in browsing. Ecography 32(4): 601–612. http://dx.doi.org/10.1111/j.1600-0587.2009.05713.x.
Månsson J., Kalén C., Kjellander P., Andrén H., Smith H. (2007). Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scandinavian Journal of Forest Research 22(5): 407–414. http://dx.doi.org/10.1080/02827580701515023.
Månsson J., Bergström R., Danell K. (2009). Fertilization – effects on deciduous tree growth and browsing by moose. Forest Ecology and Management 258(11): 2450–2455. http://dx.doi.org/10.1016/j.foreco.2009.08.025.
Närhi P., Middleton M., Gustavsson N., Hyvönen E., Sutinen M.-L., Sutinen R. (2011). Importance of soil calcium for composition of understory vegetation in boreal forests of Finnish Lapland. Biogeochemistry 102(1): 239–249. http://dx.doi.org/10.1007/s10533-010-9437-2.
Närhi P., Gustavsson N., Sutinen M.-L., Mikkola K., Sutinen R. (2013). Long-term effect of site preparation on soil quality in Tuntsa, Finnish Lapland. Geoderma 192: 1–6. http://dx.doi.org/10.1016/j.geoderma.2012.07.007.
National Land Survey of Finland (2014). The topographic database. http://www.maanmittauslaitos.fi/en/digituotteet/topographic-database. [Cited 20 November 2014].
Nevalainen S. (1999). Gremmeniella abietina in Finnish Pinus sylvestris stands in 1986–1992: a study based on the National Forest Inventory. Scandinavian Journal of Forest Research 14(2): 111–120. http://dx.doi.org/10.1080/02827589950152836.
Niemelä P., Danell K. (1988). Comparison of moose browsing on Scots pine (Pinus sylvestris) and lodgepole pine (P. contorta). Journal of Applied Ecology 25(3): 761–775. http://dx.doi.org/10.2307/2403744.
Nikula A., Heikkinen S., Helle E. (2004). Habitat selection of adult moose Alces alces at two spatial scales in central Finland. Wildlife Biology 10: 121–135.
Nikula A., Hallikainen V., Jalkanen R., Hyppönen M., Mäkitalo K. (2008). Modelling the factors predisposing Scots pine to moose damage in artificially regenerated sapling stands in Finnish Lapland. Silva Fennica 42(4): 587–603. http://dx.doi.org/10.14214/sf.235.
Noukka M. (1987). Lapin lääni, Tervola, luonto. [Province of Lapland, Tervola, nature]. In: Rikkinen K., Sihvo H. (eds.). Finlandia, Otavan iso maamme kirja. Kustannusosakeyhtiö Otava, Keuruu, Finland. p. 135–136.
Nuutinen T., Hirvelä H., Hynynen J., Härkönen K., Hökkä H., Korhonen K.T., Salminen O. (2000). The role of peatlands in Finnish wood production – an analysis based on large-scale forest scenario modelling. Silva Fennica 34(2): 131–153. http://dx.doi.org/10.14214/sf.636.
Ojala A.E.K., Palmu J.-P., Åberg A., Åberg S., Virkki H. (2013). Development of an ancient shoreline database to reconstruct the Litorina Sea maximum extension and the highest shoreline of the Baltic Sea basin in Finland. Bulletin of the Geological Society of Finland 85: 127–144.
Phelan P.L., Norris K.H., Mason J.F. (1996). Soil management history and host preference by Ostrinia nubillis: evidence for plant mineral balance mediating insect-plant interaction. Environmental Entomology 25(6): 1329–1336. http://dx.doi.org/10.1093/ee/25.6.1329.
Pulkkinen E., Sarala P. (2009). Till geochemical exploration for gold in Finnish Lapland. Geological survey of Finland, Pohjois-Suomen yksikkö, S41/2009/40, Rovaniemi. 85 p.
Sarala P., Rossi S. (2006). Rovaniemen – Tervolan alueen glasiaalimorfologiset ja -stratigrafiset tutkimukset ja niiden soveltaminen geokemialliseen malminetsintään. Summary: glacial geological and stratigraphical studies with applied geochemical exploration in the area of Rovaniemi and Tervola, southern Finnish Lapland. Geological Survey of Finland, Report of Investigation 161. 115 p.
Senft R.L., Coughenour M.B., Bailey D.W., Rittenhouse L.R., Sala O.E., Swift D.M. (1987). Large herbivore foraging and ecological hierarchies. Bioscience 37(11): 789−795. http://dx.doi.org/10.2307/1310545.
Silvennoinen A. (1998). Pohjois-Suomen liuskealueet, kerrosintruusiot ja granuliittialue. [Schist belts, layered intrusions and granulite area in northern Finland]. In: Lehtinen M., Nurmi P., Rämö T. (eds.). Suomen kallioperä, 3000 vuosimiljoonaa. Suomen Geologinen Seura, Jyväskylä. p. 141–163. ISBN: 952-90-9260-1.
Sutinen R., Teirilä A., Pänttäjä M., Sutinen M.-L. (2002). Survival of artificially regenerated Scots pine on till soils with respect to varying dielectric properties. Canadian Journal of Forest Research 32(7): 1151–1157. http://dx.doi.org/10.1139/x02-076.
Sutinen R., Hyvönen E., Ruther A., Ahl A., Sutinen M-L. (2005). Soil-driven timberline of spruce (Picea abies) in Tanaelv Belt Lapland Granulite transition, Finland. Arctic, Antarctic, and Alpine Research 37: 611–619. http://dx.doi.org/10.1657/1523-0430(2005)037[0611:STOSPA]2.0.CO;2.
Sutinen R., Kuoppamaa M., Hyvönen E., Hänninen P., Middleton M., Pänttäjä M., Teirilä A., Sutinen M.-L. (2007). Geological controls on conifer distributions and their implications for forest management in Finnish Lapland. Scandinavian Journal of Forest Research 22(6): 476–487. http://dx.doi.org/10.1080/02827580701672063.
Sutinen R., Kuoppamaa M., Hänninen P., Middleton M., Närhi P., Vartiainen S., Sutinen M.-L. (2011a). Tree species distribution on mafic and felsic fells in Finnish Lapland. Scandinavian Journal of Forest Research 26(1): 11–20. http://dx.doi.org/10.1080/02827581.2010.534109.
Sutinen R., Närhi P., Middleton M., Kuoppamaa M., Juntunen V., Hyvönen E., Hänninen P., Teirilä A., Pänttäjä M., Neuvonen S., Äikää O., Piekari M., Timonen M., Sutinen, M.-L. (2011b). Geological controls on subarctic conifer distribution. In: Nenonen K., Nurmi P.A. (eds.). Geoscience for society 125th anniversary volume. Geological Survey of Finland, Special Paper 49: 315–325. ISBN 978-952-217-136-8.
Sutinen R., Närhi P., Middleton M., Hänninen P., Timonen M., Sutinen, M-L. (2012). Advance of Norway spruce (Picea abies) onto mafic Lommoltunturi fell in Finnish Lapland during the last 200 years. Boreas 41: 387–377. http://dx.doi.org/10.1111/j.1502-3885.2011.00238.x.
Tamminen P., Mälkönen E. (2003). Metsämaiden viljavuus. [Fertility of forest soil]. In: Mälkönen E. (ed.). Metsämaa ja sen hoito. Kustannusosakeyhtiö Metsälehti, Hämeenlinna. p. 141–152. ISBN-13: 9789525118575.
Tikkanen M. (1987). Lapin lääni: luonto. [Lapland province: nature]. In: Rikkinen K., Sihvo H. (eds.). Finlandia, Otavan iso maamme kirja. Kustannusosakeyhtiö Otava, Keuruu, Finland. p. 18–22.
Vehviläinen H., Koricheva J. (2006). Moose and vole browsing patterns in experimentally assembled pure and mixed forest stands. Ecography 29(4): 497–506. http://dx.doi.org/10.1111/j.0906-7590.2006.04457.x.
Total of 65 references.
