Nelson Thiffault  ,
Alain Paquette,
Christian Messier
,
Alain Paquette,
Christian Messier
Early silvicultural guidelines for intensive management of hybrid larch plantations on fertile sub-boreal sites
Thiffault N., Paquette A., Messier C. (2017). Early silvicultural guidelines for intensive management of hybrid larch plantations on fertile sub-boreal sites. Silva Fennica vol. 51 no. 2 article id 1716. https://doi.org/10.14214/sf.1716
Highlights
- Satisfactory growth can be obtained using a wide range of site preparation intensities
- There is a net advantage of performing two motor-manual release treatments over a single release
- A second release treatment cannot be replaced by more intensive site preparation
- Planting depth had no influence on planted seedling growth after 6 years.
Abstract
Use of fast-growing tree plantations on dedicated areas is proposed as a means of reconciling fibre production with conservation objectives. Success of this approach however requires fine-tuning silvicultural scenarios so that survival and growth are optimized while management and environmental costs are minimized. This is particularly challenging for hybrid larch (Larix × marschlinsii Coaz), a shade-intolerant species planted on fertile sites in Quebec (Canada) where legislation prevents the use of chemical herbicides. In this context, multiple motor-manual release treatments are often required, with high impacts on costs and social issues related to the scarcity of a qualified workforce. We established a split-split-plot design on a recently harvested site to assess the main and interaction effects of mechanical site preparation (MSP) intensity (five modalities of trenching or mounding), motor-manual release scenario (one or two treatments) and planting depth (0–3 cm or 3–10 cm) on hybrid larch seedling growth and survival six years after planting. Mechanical site preparation intensity and planting depth did not influence seedling growth after 6 years. The lack of significant interaction between MSP and release scenarios indicates that these operations should be planned independently. A more intensive MSP treatment cannot replace a second motor-manual release on fertile sites, as proposed to reduce costs. Our results also show the significant advantage of performing two motor-manual release treatments two years apart (the first one early in the scenario), over performing a single treatment. Our study provides silvicultural guidelines for the establishment of high-yield exotic larch plantations.
Keywords
vegetation management;
survival;
growth;
scarification;
exotic tree species;
intensive silviculture;
forest functional zoning (TRIAD)
-
Thiffault,
Direction de la recherche forestière, Ministère des Forêts, de la Faune et des Parcs du Québec, 2700 rue Einstein, Québec, QC, Canada G1P 3W8; Centre d’étude de la forêt, CP 8888, Succursale Centre-ville, Montréal, QC, Canada H3C 3P8
 http://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@mffp.gouv.qc.ca
http://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@mffp.gouv.qc.ca
-
Paquette,
Centre d’étude de la forêt, CP 8888, Succursale Centre-ville, Montréal, QC, Canada H3C 3P8; Université du Québec à Montréal, Département des sciences biologiques, CP 8888, Succursale Centre-ville, Montréal, QC, Canada H3C 3P8
http://orcid.org/0000-0003-1048-9674
E-mail
alain.paquette@gmail.com
- Messier, Centre d’étude de la forêt, CP 8888, Succursale Centre-ville, Montréal, QC, Canada H3C 3P8; Université du Québec à Montréal, Département des sciences biologiques, CP 8888, Succursale Centre-ville, Montréal, QC, Canada H3C 3P8; Institut des Sciences de la Forêt tempérée (ISFORT), 58 rue Principale, Ripon, QC, Canada JOV 1V0 E-mail nelson.thiff E-mail christian.messier@uqo.ca
Received 12 October 2016 Accepted 20 February 2017 Published 2 March 2017
Views 92068
Available at https://doi.org/10.14214/sf.1716 | Download PDF

1 Introduction
Plantation forestry plays an increasing role in forest management worldwide, as it enables responding to the pressure on forests to deliver environmental, social, and economic services (Paquette and Messier 2010). For example, the use of fast-growing species in elite silvicultural scenarios (Bell et al. 2008) on dedicated forest areas (functional zoning) is proposed as an effective means of reconciling wood fibre production with conservation objectives at the landscape level (Messier et al. 2003). Exotic species are of particular interest in that context, especially in northern ecosystems where their productivity is usually much higher than that of indigenous species (Kjær et al. 2014; Tullus et al. 2012). In Quebec (Canada), hybrids of poplar (Populus spp.) and larch (Larix spp.) form the basis of intensive plantation forestry within forest functional zoning projects that test this management approach at the operational scale (Messier et al. 2009). Genetic gains obtained from recent breeding cycles for hybrid larch (Larix × marschlinsii Coaz) are expected to yield mean annual increments of ~ 10 to 12 m3 ha–1 y–1 in Quebec (Perron 2008; Perron 2010), which is higher than that of native larch (~ 6 to 8 m3 ha–1 y–1 for Larix laricina (Du Roi) K.Koch; M. Perron, pers. comm.), and about eight times that of the average provincial mean annual increment (estimated to 1.2 m3 ha–1 y–1 for the 2015–2018 period; Bureau du forestier en chef 2015).
The successful establishment and growth of tree plantations are highly sensitive to management practices (Savill et al. 1997). Exotic tree plantations are no exception; the need for adequate silviculture can even be greater than for native species plantations given the high sensitivity of some exotic species to soil conditions, competition and resource availability (e.g. Fortier et al. 2012). For example, adequate mechanical site preparation is critical to the successful establishment and growth of the hybrid Populus balsamea × P. maximowiczii due to the high sensitivity of its roots to belowground competition (Bilodeau-Gauthier et al. 2011). The success of functional zoning based on intensively managed exotic plantations relies on their capacity to provide expected yields, which requires fine-tuning of silvicultural scenarios so that survival and growth are optimized while management costs are minimized (Salmón Rivera et al. 2016). This is particularly challenging for hybrid larch plantations, a shade-intolerant species, planted on fertile sites in sub-boreal Quebec, where legislation prevents the use of chemical herbicides for forestry purposes on public lands (92% of the forest land base) (Thiffault and Roy 2011). The actual strategy for such plantations is based on the early planting of large stock seedlings produced in containers > 300 cm3, following harvest. Seedlings are planted on sites mechanically prepared by disc trenching, and submitted to motor-manual release treatments using brushsaws. Most fertile sites require at least two release treatments within the first 4–5 growing seasons. These motor-manual release treatments are three times more expensive than herbicide applications for vegetation management (Dampier et al. 2006), and require a considerable qualified workforce. There is thus an incentive to evaluate if more intensive mechanical site preparations could reduce the need for multiple vegetation release treatments. Planting depth is also a significant issue for plantation establishment (Paquette et al. 2011); based on tradition, deep planting is sometimes financially penalized, with significant but not always justified impacts on the profitability of exotic plantations. The real impacts of planting depth on hybrid larch seedling growth and survival remain to be verified.
Our objective was thus to assess the main and interaction effects of site preparation, motor-manual release scenarios and planting depth on sixth year dimensions, growth and survival of hybrid larch seedlings planted on a fertile sub-boreal site. More specifically, we tested the hypothesis that a silvicultural scenario comprising two motor-manual release treatments can be replaced by a single-release scenario combined with a more intensive site preparation treatment than the normally used single-pass trenching. We also posited that planting depth does not have a significant impact on seedling dimensions, growth and survival. This paper follows a previous study (Buitrago et al. 2015) that presented results after the first two years and thus could not consider the effect of the release treatments.
2 Materials and methods
2.1 Study region
We used an experimental plantation (47°37´N, 72°49´W) established near the city of La Tuque in Quebec (Canada). Details on the study region, experimental design and treatments are available in Buitrago et al. (2015). In summary, the site is located within the balsam fir (Abies balsamea (L.) Mill.) – yellow birch (Betula alleghaniensis Britton) bioclimatic domain described by Saucier et al. (2009). The region is characterized by a cold continental climate with temperate summers and a growing season of 170 days. Average annual temperature and precipitation are 3.4 °C and 940 mm, respectively. The podzolic soil formed from a coarse-textured glacial till 50–100 cm thick with moderate drainage. The previous stand (~ 30 ha) was dominated by balsam fir, paper birch (Betula papyrifera Marshall), yellow birch, red maple (Acer rubrum L.) and black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.). It was harvested in October 2009 with careful logging around advanced regeneration. Advanced regeneration was scarce though (low density and low stocking). It mainly consisted in balsam fir, paper birch, yellow birch, and red maple. Patches where regeneration density and height were high were not scarified and kept as biological legacies.
2.2 Experimental design and seedling measurements
The experimental setup established on the clearcut consisted of a split-split-plot design with three replicate blocks. Each block contained five main plots of ~ 1 ha each, to which we randomly applied one of the following mechanical site preparation treatments (Buitrago et al. 2015): (i) simple-pass trenching (T) performed with a T26 Bracke disk trencher; (ii) double adjacent trenching (T-Da), implying two passes over the same trench, producing a deeper and wider area of exposed mineral soil than (i); (iii) double intensive trenching (T-Di) using two passes over the same row but with amplified disk angle, thus producing deeper and wider trenches than (i) and (ii); (iv) mounding (M), performed with an excavator equipped with a 60 cm wide bucket that excavated and upturned the mineral soil, creating a roughly conical elevated mass beside the hole; and (v) inverting (M-inv), a treatment in which the bucket excavated, upturned the material and replaced it in its original hole. The experiment did not include a non-scarified control, as empirical observations have already demonstrated the need for soil perturbation when planting hybrid larch in this region. Conventional trenching (i.e. treatment i) was considered as the minimal base treatment in this context.
Each main plot was divided into two sub-plots to which we randomly assigned one of two scenarios of motor-manual release from competing vegetation (dominated by Rubus idaeus L. and Prunus pensylvanica L. f.): a single motor-manual release done in August of the third growing season after planting (Single Release 3), or two release treatments performed in August of the second and fourth growing seasons after planting (Double Release 2–4). Release was performed over the entire sub-plot area using motor-manual brushsaws. Due to operational constraints, the two sub-plots of one of the replicates were both submitted to the ‘Double Release 2–4’ scenario, thus resulting in an unbalanced randomized block design.
In April 2010, we further divided each sub-plot into two sub-sub-plots and planted large stock seedlings of hybrid larch produced in 320 cm3 containers over 2 years, with their root collar positioned at either 0–3 cm or 3–10 cm depth relative to the soil surface. Larix × marschlinsii seedlings (clone MEH-C2-ALO-2-1) were obtained from a governmental nursery (Saint-Modeste, Quebec, Canada); this hybrid results from the crossing of European larch (Larix decidua Mill.) with Japanese larch (Larix kaempferi (Lamb.) Carrière). The seedling lot was (mean ± SD) 42.6 cm ± 9.6 in height and 6.0 mm ± 1.0 in diameter at the nursery, immediately prior shipping to the site. Seedlings were planted 2 m apart in the trenching treatments (planting density of 2000 seedlings ha–1) and 3 m apart in the mounding treatments (planting density of 1100 seedlings ha–1). We established a 200 m2 circular sampling unit in the centre of every sub-sub-plot, in which we measured all seedlings for height (cm) and ground-level diameter (mm) at the time of planting (respectively 38.5 cm ± 10.0 and 4.6 mm ± 1.0) and after their first, second, fourth and sixth growing season. Survival was also assessed.
2.3 Statistical analyses
We submitted sixth-year height, ground-level diameter and absolute growth rates over the 2013–2015 period (calculated as [dimensionautumn 2015 – dimensionautumn 2013] / 2) to analyses of variance (ANOVA) with respect to the experimental design using the function lmer of the package ‘lme4’ (Bates et al. 2015) in R v3.0.2 (R Core Team 2013). We focused on the 2013–2015 period for growth assessment as it better represents seedling responses to the scenario of mechanical release (initiated in 2011 and terminated in 2013) than the average growth since planting. We considered mechanical site preparation treatments (5 levels), scenarios of motor-manual release (2 levels), planting depth (2 levels), and their interactions as fixed effects. Blocks and interactions with blocks were considered as random effects. The statistical model was thus:

where yijkh is the measure of variable y in block i, submitted to mechanical site preparation j, scenario of motor-manual release k, and planting depth h, µ is the general mean, βi, τj, γk and δh are random block, fixed mechanical site preparation, fixed scenarios of motor-manual release and fixed planting depth effects, respectively, and εijkh is the error term.
We used Type III ANOVAs to take into account the unbalanced nature of the design (Steel et al. 1997). Data were verified for ANOVA prerequisites using standard graphical procedures. We calculated denominator degrees of freedom using the Satterthwaite method (Spilke et al. 2005). Survival after six growing seasons was analyzed with a generalized linear mixed-effect model using the glmer function of lme4, specifying a binomial distribution and a logit link.
3 Results
Site preparation intensity and planting depth did not significantly influence seedling height and diameter measured six years after planting (Table 1, Fig. 1). These treatments did not significantly affect seedling growth over the 2013–2015 period (Table 1).
| Table 1. ANOVA results for hybrid larch dimensions six growing seasons after planting, and growth over the last two growing seasons. | ||||||||||
| Effect (fixed) | ndf | ddf | Height 2015 | Absolute growth rate in height 2013–2015 | Ground-level diameter 2015 | Absolute growth rate in ground-level diameter 2013–2015 | ||||
| F-value | Pr > F | F-value | Pr > F | F-value | Pr > F | F-value | Pr > F | |||
| Site Preparation (SP) | 4 | 7.7 | 0.657 | 0.639 | 0.289 | 0.876 | 0.783 | 0.553 | 0.518 | 0.725 |
| Release scenario (Rel) | 1 | 28.9 | 48.502 | < 0.001 | 44.959 | < 0.001 | 43.303 | < 0.001 | 18.409 | 0.002 |
| SP×Rel | 4 | 27.4 | 0.083 | 0.987 | 0.263 | 0.899 | 0.383 | 0.817 | 0.265 | 0.894 |
| Planting depth (PD) | 1 | 27.2 | 0.267 | 0.609 | 0.018 | 0.894 | 0.493 | 0.491 | 0.363 | 0.554 |
| SP×PD | 4 | 26.4 | 1.358 | 0.275 | 0.619 | 0.653 | 1.611 | 0.213 | 1.508 | 0.240 |
| Rel×PD | 1 | 27.2 | 0.710 | 0.407 | 0.018 | 0.893 | 1.591 | 0.222 | 0.861 | 0.365 |
| SP×Rel×PD | 4 | 26.4 | 1.019 | 0.416 | 0.935 | 0.459 | 0.801 | 0.540 | 1.215 | 0.338 |
| ndf = numerator degrees of freedom; ddf = denominator degrees of freedom. ddf were calculated using the Satterthwaite method (Spilke et al. 2005) and thus vary depending on the response variables. For brevity, ddf are presented for ‘Height 2015’ only. | ||||||||||
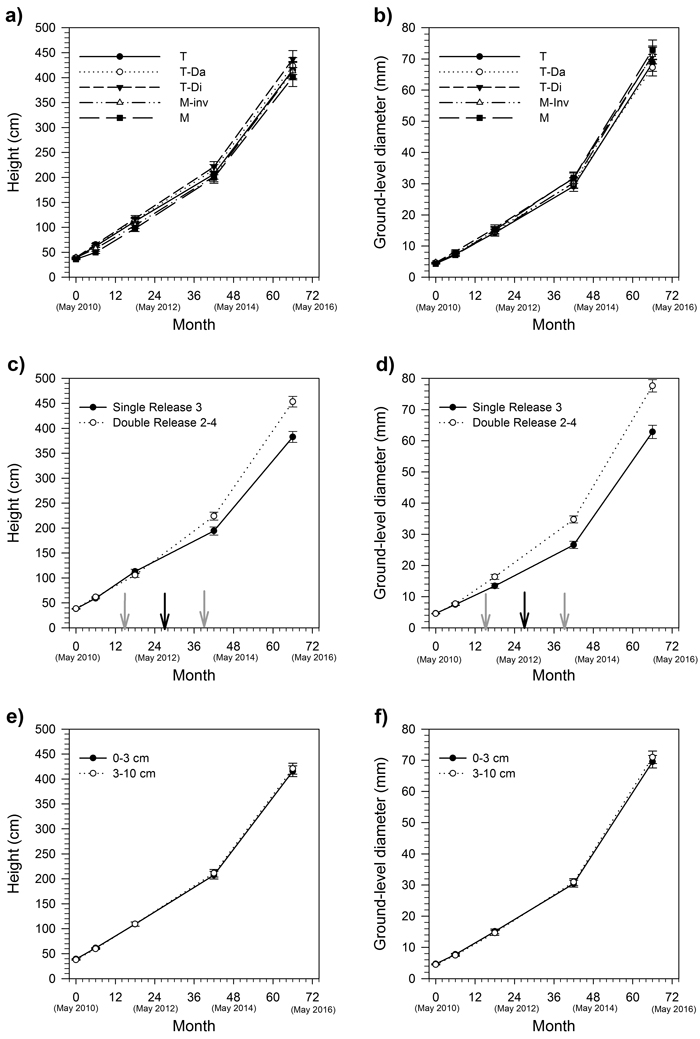
Fig. 1. Height (a, c, e) and diameter (b, d, f) profiles of hybrid larch seedlings as influenced by site preparation (a, b), motor-manual release scenario (c, d) and planting depth (e, f) over six growing seasons. Data are presented as mean ± standard error. In c and d, black arrows indicate the timing of the ‘Single Release 3’ treatment; grey arrows indicate the timing of the ‘Double Release 2–4’ treatments. Analyses were conducted on final dimensions and on absolute growth rates over the last 24 months; full profiles are provided for information only. T = simple trenching; T-Da = double adjacent trenching; T-Di = double intensive trenching; M = mounding; M-inv = inverting.
The ‘Double Release 2–4’ scenario significantly increased seedling height and ground-level diameter by 18% and 23%, respectively, compared to the ‘Single Release 3’ scenario (Table 1, Fig. 1). Differences in seedling dimensions between release scenarios are expected to continue increasing as the treatments also had a significant effect on height growth (p < 0.001) and diameter growth (p = 0.002) over the 2013–2015 period. Seedlings submitted to the ‘Single Release 3’ scenario presented an annual growth of 93 cm y–1 and 18 mm y–1 in height and diameter, respectively. Over the same two-year period, seedlings planted in the ‘Double Release 2–4’ plots presented an annual growth of 115 cm y–1 and 22 mm y–1 in height and diameter growth, respectively.
Site preparation, release scenario and planting depth interacted to influence seedling survival, but the release treatment effect was by far the strongest based on an F value > 22 (Table 2). The lowest survival was observed for seedlings planted 3–10 cm deep in the double adjacent site preparation treatment and submitted to a single motor-manual release treatment in 2012 (Fig. 2). The ‘Double Release 2–4’ scenario was generally associated with treatment combinations that resulted in the highest survival (sometimes > 90%; Fig. 2), with the exception of the double intensive trenching plots where survival was highest in the ‘Single Release 3’ × ‘0–3 cm’ combination.
| Table 2. ANOVA results for hybrid larch survival six growing seasons after planting. | ||||
| Effect (fixed) | ndf | ddf | Survival | |
| F-value | Pr > F | |||
| Site Preparation (SP) | 4 | 8 | 3.244 | 0.052 |
| Release scenario (Rel) | 1 | 9 | 22.540 | 0.006 |
| SP×Rel | 4 | 9 | 3.113 | 0.047 |
| Planting depth (PD) | 1 | 19 | 1.723 | 0.063 |
| SP×PD | 4 | 19 | 4.295 | 0.006 |
| Rel×PD | 1 | 19 | 7.948 | 0.002 |
| SP×Rel×PD | 4 | 19 | 2.132 | 0.044 |
| ndf = numerator degrees of freedom; ddf = denominator degrees of freedom. | ||||
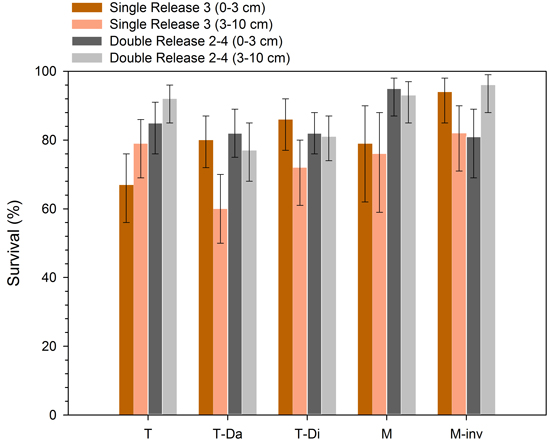
Fig. 2. Survival of hybrid larch seedlings after six growing seasons, as influenced by combinations of site preparation (T = simple trenching; T-Da = double adjacent trenching; T-Di = double intensive trenching; M = mounding; M-inv = inverting), motor-manual release scenario (Single Release 3; Double Release 2–4) and planting depth (0–3 cm; 3–10 cm). Data are presented as mean ± 95% CI, back-transformed from the logit scale.
4 Discussion
Our results have four main implications for intensive silviculture of hybrid larch in sub-boreal ecosystems. First, they confirm earlier studies that planting depth has little influence on planted seedling growth given that trees are located on adequate microsites, following proper site preparation and that basic vegetation management is carried out. This observation is in line with the growing body of literature describing the effects of planting depth on the growth of other boreal conifers such as black spruce, white spruce (Picea glauca (Moench.) Voss) and jack pine (Pinus banksiana L.) (Paquette et al. 2011; Thiffault 2016). For hybrid larch in particular, our results confirm that, just as planting depth had little influence after two years (Buitrago et al. 2015), it continued to have little effect on growth after 6 years. The effect of planting depth on seedling survival varied according to the mechanical site preparation treatment and the motor-manual release scenario, but the triple interaction was barely significant and weak compared to that of motor-manual release. In the worst case (Double adjacent trenching × Single Release 3 × 3–10 cm), we observed more than 40% mortality after six growing seasons. Competition, soil temperature, soil moisture and soil nutrient availability, as influenced by silvicultural practices, have been shown to interact in complex ways to influence planted seedling physiology (e.g. Trottier-Picard et al. 2016). The production of adventitious roots by larch potentially interacts with the chemical and physical characteristics of the rooting environment produced by the mechanical site preparation treatments and planting depths to influence seedling survival (Calvo-Polanco et al. 2012). Unfortunately, we did not perform detailed physiological or nutritional measurements on the seedlings; such information would have been necessary to explain the survival responses we observed and disentangle the driving mechanisms. Our results thus suggest that release from competition is the main driver of hybrid larch survival, but that minor adjustments to planting practices could, in certain cases, reduce early mortality (for example, favour deep planting in simple-pass trenching treatments; Fig. 2).
Secondly, our results show that similar growth of hybrid larch can be obtained using a wide range of site preparation intensities, and that no significant gains are obtained with more intensive mechanical site preparation. Although increased seedling growth is often correlated with soil treatment intensity, such as observed on less fertile boreal sites (e.g. Thiffault and Jobidon 2006), interactions with biophysical site characteristics (e.g. soil texture, humus thickness, dominant competing vegetation) can also lead to a limited effect of soil management intensity on planted seedling performance (e.g. Fleming et al. 2014). In such contexts, managers can adapt prescriptions based on environmental, economic or social constraints, with limited (if any) impact on productivity (Barrette et al. 2014). Seedling survival was shown to vary from 60% to more than 90%, depending on the treatment combination. Taking this into account, our study shows that mounding or inverting could be preferred to disking for the establishment of hybrid larch plantations, as these modalities favoured the highest survival rates. Other factors can however influence the choice of a mechanical site preparation treatment, such as the spatial distribution of soil disturbance and associated environmental effects (excavator mounding or inverting having little to no effect outside the treated area), the desire to favor the emergence of natural regeneration between the planted trees, treatment costs relative to the number of suitable microsites created, or the social acceptability of intensive treatments. If these concerns are preeminent, other treatment combinations such as single-pass trenching can be satisfactory alternatives in terms of seedling survival.
Thirdly, our study demonstrates the significant advantage of performing two motor-manual release treatments two years apart (the first one being early in the scenario), over performing a single treatment at some point following planting. This effect was verified in terms of seedling size, growth and survival, which confirms that controlling competing vegetation is critical to ensure survival and growth of exotic Larix species in Canadian ecosystems (Gilmore and David 2002). Early vegetation management is usually beneficial for conifers; diameter growth typically increases in the years following cleaning treatments (e.g. Uotila and Saksa 2013). Whereas slow growing boreal species such as black spruce do not significantly respond to multiple release treatments compared to single vegetation control (Jobidon and Charette 1997), Larix benefits from repeated interventions because it is a light-demanding species with high potential rates of resource capture (Reich et al. 1998). Our results are of particular interest given the general trend in reducing the use of chemical herbicides for forest vegetation management in Europe and elsewhere (Ammer et al. 2011). They illustrate the critical need to further fine-tune management strategies in such contexts, especially the timing of motor-manual release operations (e.g. Thiffault et al. 2014). Indeed, the sooner the planted trees are released from herbaceous competition the better, in that they would require less investment over the whole rotation, and attain commercial maturity sooner, further increasing the profitability of early interventions.
Finally, our study shows that a more intensive mechanical site preparation cannot be used to replace a motor-manual release scenario based on two treatments on fertile sites such as the one studied here. Intensive mechanical site preparation is effective in controlling competing vegetation in some ecosystems, such as harvested boreal sites with understory layers dominated by ericaceous species (Thiffault and Jobidon 2006) or grass (Nilsson and Örlander 1999). However, sub-boreal mixedwoods are characterized by the rapid post-harvest establishment of early successional species, such as red raspberry (Rubus idaeus L.) and pin cherry (Prunus pensylvanica L. f.) (Laflèche et al. 2000), that reproduce through seed banks, root suckering and stump sprouting. Although the creation of large trenches or mounds potentially slows down the encroachment of planting microsites by competing species, the high level of solar radiation and its impact on soil temperature stimulates the emergence and growth of these competing species (Jobidon 1995). The general lack of significant interactions between site preparation treatments and motor-manual release scenarios indicates that these operations should be planned independently.
5 Conclusion
Our results have the following implications for the early silviculture of hybrid larch plantations established on fertile sub-boreal sites. They confirm that a wide range of mechanical site preparation intensities, from conventional simple-pass disk trenching to mounding and inverting, can be used to obtain satisfactory growth. It enables managers to select the best site preparation method given other operational, environmental, economic or social constraints. Our results show that after 6 years, more intensive mechanical site preparation treatments are not reducing the need for performing mechanical release from competing vegetation; the treatments should be planned independently. Performing two motor-manual release treatments significantly increases growth compared to performing a single release, and generally reduces mortality. Finally, planting depth has no influence on planted seedling growth even after 6 years clearly indicating that deeper planting should be tolerated, and possibly even encouraged when performing simple-pass trenching treatments, as it tends to increase survival.
Acknowledgements
We thank P. Gaucher, C. Jenkins, M. Hänsel, É. Rousselle, J. Carignan, S. Lemay, D. Létourneau, M. Brémont, T. André, Y. Gauthier, M. Buitrago, and E. Gaillard for their technical support and D. Lesieur for his help in database management. We are indebted to J. DeBlois for her constructive advice regarding the statistical analyses, as well as to M. Girard, N. Beaulieu and many others at Resolute Forest Products (Mauricie, QC). Larch hybrids used in this study are the result of continuous selection efforts from the Quebec provincial nurseries. We thanks the anonymous reviewers for providing helpful comments on an earlier version of the manuscript. This study is part of the TRIAD initiative, with the collaboration of the Direction de la recherche forestière of the Ministère des Forêts de la Faune et des Parcs du Québec and Resolute Forests Products.
References
Ammer C., Balandier P., Bentsen N.S., Coll L., Löf M. (2011). Forest vegetation management under debate: an introduction. European Journal of Forest Research 130(1): 1–5. https://doi.org/10.1007/s10342-010-0452-6.
Barrette M., Leblanc M., Thiffault N., Paquette A., Lavoie L., Bélanger L., Bujold F., Côté L., Lamoureux J., Schneider R., Tremblay J.-P., Côté S., Boucher Y., Deshaies M.-È. (2014). Issues and solutions for intensive plantation silviculture in a context of ecosystem management. The Forestry Chronicle 90(6): 748–762. https://doi.org/10.5558/tfc2014-147.
Bates D., Mächler M., Bolker B.M., Walker S.C. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1): 1–48. https://doi.org/10.18637/jss.v067.i01.
Bell F.W., Parton J., Stocker N., Joyce D., Reid D., Wester M., Stinson A., Kayahara G., Towill B. (2008). Developing a silvicultural framework and definitions for use in forest management planning and practice. The Forestry Chronicle 84(5): 678–693. https://doi.org/10.5558/tfc84678-5.
Bilodeau-Gauthier S., Paré D., Messier C., Bélanger N. (2011). Juvenile growth of hybrid poplars on acidic boreal soil determined by environmental effects of soil preparation, vegetation control, and fertilization. Forest Ecology and Management 261(3): 620–629. https://doi.org/10.1016/j.foreco.2010.11.016.
Buitrago M., Paquette A., Thiffault N., Bélanger N., Messier C. (2015). Early performance of planted hybrid larch: effects of mechanical site preparation and planting depth. New Forests 46(3): 319–337. https://doi.org/10.1007/s11056-014-9463-3.
Bureau du forestier en chef (2015). État de la forêt publique du Québec et de son aménagement durable. Bilan 2008–2013. [The state of the public forest in Quebec and of its sustainable management. 2008–2013 assessment]. Gouvernement du Québec, Roberval, QC. 382 p.
Calvo-Polanco M., Señorans J., Zwiazek J.J. (2012). Role of adventitious roots in water relations of tamarack (Larix laricina) seedlings exposed to flooding. BMC Plant Biology 12(99): 1–9. https://doi.org/10.1186/1471-2229-12-99.
Dampier J.E.E., Bell F.W., St-Amour M., Pitt D.G., Luckai N.J. (2006). Cutting versus herbicides: tenth-year volume and release cost-effectiveness of sub-boreal conifer plantations. The Forestry Chronicle 82(4): 521–528. https://doi.org/10.5558/tfc82521-4.
Fleming R.L., Leblanc J.D., Hazlett P.W., Weldon T., Irwin R., Mossa D.S. (2014). Effects of biomass harvest intensity and soil disturbance on jack pine stand productivity: 15-year results. Canadian Journal of Forest Research 44(12): 1566–1574. https://doi.org/10.1139/cjfr-2014-0008.
Fortier J., Truax B., Gagnon D., Lambert F. (2012). Hybrid poplar yields in Québec: implications for a sustainable forest zoning management system. The Forestry Chronicle 88(4): 391–407. https://doi.org/10.5558/tfc2012-075.
Gilmore D.W., David A.J. (2002). Current trends in management practices for European larch in North America. The Forestry Chronicle 78(6): 822–829. https://doi.org/10.5558/tfc78822-6.
Jobidon R. (1995). Autécologie de quelques espèces de compétition d’importance pour la régénération forestière au Québec. Revue de littérature. [Autecology of some important species competing with forest regeneration in Quebec. Literature review]. Mémoire de recherche forestière 117. Direction de la recherche forestière, Ministère des Ressources naturelles, Québec, QC. 180 p.
Jobidon R., Charette L. (1997). Effets, après 10 ans, du dégagement manuel simple ou répété et de la période de coupe de la végétation de compétition sur la croissance de l’épinette noire en plantation. [Effects, after 10 years, of single or repeated manual release and of time of cutting competing vegetation on the growth of planted black spruce]. Canadian Journal of Forest Research 27(12): 1979–1991. https://doi.org/10.1139/x97-166.
Kjær E.D., Lobo A., Myking T. (2014). The role of exotic tree species in Nordic forestry. Scandinavian Journal of Forest Research 29(4): 323–332. https://doi.org/10.1080/02827581.2014.926098.
Laflèche V., Ruel J.-C., Archambault L. (2000). Évaluation de la coupe avec protection de la régénération et des sols comme méthode de régénération de peuplements mélangés du domaine bioclimatique de la sapinière à bouleau jaune de l’est du Québec, Canada. [Evaluation of cutting with protection of regeneration and soils to regenerate mixed stands in the eastern balsam fir – yellow birch bioclimatic domain of Quebec]. The Forestry Chronicle 76(4): 653–663. https://doi.org/10.5558/tfc76653-4.
Messier C., Bigué B., Bernier L. (2003). Using fast-growing plantations to promote forest ecosystem protection in Canada. Unasylva 54: 59–63.
Messier C., Tittler R., Kneeshaw D.D., Gélinas N., Paquette A., Berninger K., Rheault H., Meek P., Beaulieu N. (2009). TRIAD zoning in Quebec : experiences and results after 5 years. The Forestry Chronicle 85(6): 885–896. https://doi.org/10.5558/tfc85885-6.
Nilsson U., Örlander G. (1999). Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Canadian Journal of Forest Research 29(7): 1015–1026. https://doi.org/10.1139/x99-071.
Paquette A., Messier C. (2010). The role of plantations in managing the world’s forests in the Anthropocene. Frontiers in Ecology and the Environment 8(1): 27–34. https://doi.org/10.1890/080116.
Paquette A., Girard J., Walsh D. (2011). Deep planting has no short- or long-term effect on the survival and growth of white spruce, black spruce, and jack pine. Northern Journal of Applied Forestry 28: 146–151.
Perron M. (2008). A strategy for the second breeding cycle of Larix × marschlinsii in Québec, Canada including experiments to guide interspecific tree breeding programme. Silvae Genetica 57: 282–291.
Perron M. (2010). Résultats de la sélection des parents en vue de la deuxième génération d’amélioration du mélèze hybride (Larix × marschlinsii Coaz.) au Québec. [Results of the selection of parents of hybrid larch (Larix × marschlinsii Coaz.) breeding populations for second generation]. Ministère des Ressources naturelles et de la Faune, Direction de la recherche forestière, Québec, QC. 41 p.
R Core Team. (2013). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Reich P.B., Tjoelker M.G., Walters B., Vanderklein D., Buschena C. (1998). Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light. Functional Ecology 12(3): 327–338. https://doi.org/10.1046/j.1365-2435.1998.00208.x.
Salmón Rivera B., Barrette M., Thiffault N. (2016). Issues and perspectives on the use of exotic species in the sustainable management of Canadian forests. Reforesta 1: 261–280. https://doi.org/10.21750/REFOR.1.13.13.
Saucier J.P., Robitaille A., Grondin P. (2009). Cadre bioclimatique du Québec. [Bioclimatic context of Quebec]. In: Doucet R., Côté M. (eds.). Manuel de foresterie, 2nd edition. Ordre des ingénieurs forestiers du Québec, Éditions Multimondes, Québec, QC. p. 186–205.
Savill P., Evans J., Auclair D., Falk J. (1997). Plantation silviculture in Europe. Oxford University Press, New York, NY. 297 p.
Spilke J., Piepho H.-P., Hu X. (2005). A simulation study on tests of hypotheses and confidence intervals for fixed effects in mixed models for blocked experiments with missing data. Journal of Agricultural, Biological, and Environmental Statistics 10(3): 374–389. https://doi.org/10.1198/108571105X58199.
Steel R.G.D., Torrie J.H., Dickey D.A. (1997). Principles and procedures of statistics. A biometrical approach, 3rd edn. WCB/McGraw-Hill, Boston, MA. 666 p.
Thiffault N. (2016). Short-term effects of organic matter scalping on the growth and nutrition of black spruce and jack pine seedlings planted in the boreal forest. The Forestry Chronicle 92(2): 221–231. https://doi.org/10.5558/tfc2016-041.
Thiffault N., Jobidon R. (2006). How to shift unproductive Kalmia angustifolia - Rhododendron groenlandicum heath to productive conifer plantation. Canadian Journal of Forest Research 36(10): 2364–2376. https://doi.org/10.1139/x06-090.
Thiffault N., Roy V. (2011). Living without herbicides in Québec (Canada): historical context, current strategy, research and challenges in forest vegetation management. European Journal of Forest Research 130(1): 117–133. https://doi.org/10.1007/s10342-010-0373-4.
Thiffault N., Hebert F., Charette L., Jobidon R. (2014). Large spruce seedling responses to the interacting effects of vegetation zone, competing vegetation dominance and year of mechanical release. Forestry 87(1): 153–164. https://doi.org/10.1093/forestry/cpt048.
Trottier-Picard A., Thiffault E., Thiffault N., DesRochers A., Paré D., Messier C. (2016). Complex impacts of logging residues on planted hybrid poplar seedlings in boreal ecosystems. New Forests 47(6): 877–895. https://doi.org/10.1007/s11056-016-9550-8.
Tullus A., Rytter L., Tullus T., Weih M., Tullus H. (2012). Short-rotation forestry with hybrid aspen (Populus tremula L.×P. tremuloides Michx.) in Northern Europe. Scandinavian Journal of Forest Research 27(1): 10–29. https://doi.org/10.1080/02827581.2011.628949.
Uotila K., Saksa T. (2013). Effects of early cleaning on young Picea abies stands. Scandinavian Journal of Forest Research 29(2): 111–119. https://doi.org/10.1080/02827581.2013.869349.
Total of 37 references.
