Laura Pikkarainen  ,
Harri Strandman,
Eerik Vento,
Aaron Petty,
Olli-Pekka Tikkanen,
Antti Kilpeläinen,
Heli Peltola
,
Harri Strandman,
Eerik Vento,
Aaron Petty,
Olli-Pekka Tikkanen,
Antti Kilpeläinen,
Heli Peltola
Effects of forest conservation and management on timber, ecosystem carbon, dead wood and habitat suitability area in a boreal forest under climate change
Pikkarainen L., Strandman H., Vento E., Petty A., Tikkanen O.-P., Kilpeläinen A., Peltola H. (2024). Effects of forest conservation and management on timber, ecosystem carbon, dead wood and habitat suitability area in a boreal forest under climate change. Silva Fennica vol. 58 no. 2 article id 23045. https://doi.org/10.14214/sf.23045
Highlights
- Increasing forest conservation areas increased all other ecosystem services except timber yield. Intensive forest management enhanced this increase
- Increased conservation area decreased timber yield, but intensive forest management reduced this effect and even overcompensated it at the end of simulation period in the 10% conservation scenario with intensified forest management
- Climate change increased all other ecosystem services, except carbon stocks.
Abstract
We used forest ecosystem model simulations to study how forest conservation and management intensity affected timber yield, ecosystem carbon stocks, amount of dead wood, and habitat suitability area in a middle boreal forest region of Finland under changing climate over a 90-year simulation period. We used the following forest conservation and management scenarios: baseline forest management (BM), BM with 10 or 20% increase of conservation area with or without intensified forest management (i.e. improved forest regeneration material and forest fertilization). The simulations were done under current climate (reference period of 1981–2010), and Representative Concentration Pathway (RCP) climate change projections under the RCP2.6 and RCP4.5 forcing scenarios. Overall, increasing the forest conservation area decreased timber yield and increased the ecosystem carbon stock, the amount of dead wood and consequently the area of suitable habitat for saproxylic species. The use of intensified forest management reduced the loss of timber yield, increased ecosystem carbon stock, the amount of dead wood and area of suitable habitat for saproxylic species. At the end of simulation period, the use of intensified forest management even overcompensated (4–6% higher) the timber loss from 10% increase of conservation area. Under changing climate, timber yield, the amount of dead wood and the area of suitable habitats for saproxylic species increased. To conclude, with intensified forest management it is possible, in the short term, to decrease the loss of timber yield through increased forest conservation area and in the long term maintain or even increase it compared to baseline forest management.
Keywords
saproxylic species;
simulation;
carbon storage;
forest fertilization;
Fennoscandia;
forest ecosystems services;
improved regeneration material
-
Pikkarainen,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
 https://orcid.org/0000-0001-5301-3639
E-mail
laura.pikkarainen@uef.fi
https://orcid.org/0000-0001-5301-3639
E-mail
laura.pikkarainen@uef.fi
-
Strandman,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
https://orcid.org/0000-0002-9400-6424
E-mail
harri.strandman@uef.fi
- Vento, School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland E-mail eerik.vento@gmail.com
-
Petty,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
https://orcid.org/0009-0006-6595-1386
E-mail
aaron.petty@uef.fi
-
Tikkanen,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
https://orcid.org/0000-0002-3875-2772
E-mail
olli-pekka.tikkanen@uef.fi
-
Kilpeläinen,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
https://orcid.org/0000-0003-4299-0578
E-mail
antti.kilpelainen@uef.fi
- Peltola, School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland E-mail heli.peltola@uef.fi
Received 22 August 2023 Accepted 25 February 2024 Published 13 March 2024
Views 85783
Available at https://doi.org/10.14214/sf.23045 | Download PDF

1 Introduction
Boreal forests provide in addition to timber also various other ecosystem services to society (Hetemäki et al. 2022). Forests sequester and store large amounts of carbon, therefore supporting climate change mitigation efforts. In addition, dead wood of boreal forests accommodates numerous saproxylic species (Tikkanen et al. 2006, 2007). In Finland, the demand of wood for the forest industry has been increasing, which has raised concerns about the sustainability of commonly utilized forest practices (Triviño et al. 2017; Hetemäki et al. 2022). At the same time, an increase in forest conservation areas have been suggested for Finland and elsewhere in Europe (European Commission 2020).
Forest management and harvesting intensity may have large impact on ecosystem services and their trade-offs (Felton et al. 2016; Pohjanmies et al. 2017; Heinonen et al. 2017; Diaz et al. 2020, 2021; Triviño et al. 2023). Especially, intensive timber harvesting decreases ecosystem carbon stocks and biodiversity values of forests (Heinonen et al. 2017; Triviño et al. 2017). On the other hand, forest management practices, such as use of improved forest regeneration material and fertilization (Saarsalmi and Mälkönen 2001; Hedwall et al. 2014; Ruotsalainen 2014; Jansson et al. 2017; Heinonen et al. 2018b; Routa et al. 2019), might simultaneously enhance carbon sequestration and timber yield, and consequently make possible to set aside more forests, e.g. for conservation.
Climate change and the related increases in natural disturbances affect the functioning of forests and consequently the provisioning of different ecosystem services (Gauthier et al. 2015; Seidl et al. 2014; Reyer et al. 2017; Venäläinen et al. 2020). However, forest management has a greater impact on ecosystem services than gradual climate change (Alrahahleh et al. 2016, 2018a, 2018b; Triviño et al. 2023). This is although forest growth and carbon sequestration are mostly expected to increase under climate change in boreal conditions (Kellomäki et al. 2008; Poudel et al. 2011, 2012; Kellomäki et al. 2018). However, a warming climate may also make growing conditions sub-optimal for some tree species and increase the mortality of trees under such conditions (Kellomäki et al. 2018).
Nearly half of the threatened forests species live in old forests or in forests rich in dead wood (Hyvärinen et al. 2019). Forest management practices have altered the structure and age composition of managed forests and reduced existence of old and primary forests along with the amount and quality of dead wood (Hyvärinen et al. 2019; Korhonen et al. 2020). In Finland, the mean volume of decayed dead wood and standing and fallen dead trees on forest land as measured by the latest national inventory (NFI13) is currently approximately 6.4 m3 ha–1 (Kulju et al. 2023). According to previous research saproxylic species prefer sites where there is > 20 m3 ha–1 of dead wood available (Penttilä et al. 2004; Müller and Bütler 2010; Junninen and Komonen et al. 2011; Gossner et al. 2013). In natural forests there can be about 60–120 m3 ha–1 of dead wood, depending on site-specific conditions (Peuhu and Siitonen 2011).
The aim of the EU Biodiversity Strategy is to stop and reverse the loss of biodiversity by placing minimum of 30% EU land (and sea) areas under legal conservation of which 10% must be under strict conservation by 2030. Priority should be given to primary and old forests (European commission 2020). In Finland, about 2.4 Mha (10.6%) of forest land and poorly productive forests are legally protected of which about 2.2 Mha is strictly protected (Kotiaho et al. 2021). In strictly protected areas ecological processes are left undisturbed from human activities, while statutory protected areas entail legal protection with the possibility of moderate logging (Kotiaho et al. 2021; European commission 2022). The share of conservation area is overall higher in the northern part of Finland and in poorly productive forests. According to Kotiaho et al. (2021) strong bias in conservation to northern and/or unproductive forests does not maintain biological diversity in the whole of Finland. Sufficient protection benefit is achieved when the 10% strict conservation target is reached in each province in Finland. In the simulation study of Räty et al. (2023), implementation of EU Biodiversity Strategy 2030 resulted in lower total growing stock and increased proportion of younger forests in managed forests when the target harvest levels remained on current level under current climate.
To ensure sustainable management and utilization of forests, we need a better understanding on how different ecosystem services (e.g. timber production, carbon sequestration/stocks and biodiversity indicators like the amount of dead wood) are affected by the intensity of forest management, forest conservation and the severity of climate change (Seidl et al. 2007; Triviño et al. 2023). In the above context, we studied based on forest ecosystem model simulations how forest conservation and management intensity affect to timber yield, ecosystem carbon stocks, and the amount of dead wood with implications on the area of suitable habitats for saproxylic species under different climate projections. We conducted 90-year simulations for the province of north Karelia using five management scenarios: baseline forest management (BM); BM with increased conservation area by 10 and 20%; BM with increased conservation area combine with intensive forest management (improved forest regeneration material and forest fertilization). The simulations were done under the current climate (reference period of 1981–2010), and Representative Concentration Pathway (RCP) projections under the RCP2.6 and RCP4.5 forcing scenarios. We hypothesized that increased forest conservation area will decrease the timber yield if intensive forest management is not used, whereas its use may help to maintain or even increase timber yield in conservation scenarios compared to the baseline forest management.
2 Material and methods
2.1 Outlines of the forest ecosystem model (SIMA) and its simulations
This study used a gap-type forest ecosystem model (SIMA) (Kellomäki et al. 2008, 2019). The model simulates forest regeneration, growth and mortality of main boreal tree species growing on upland forests throughout Finland. The growth and dynamics of forests are controlled by forest management, and climatic and site conditions. In the model, the growth of tree species is calculated as a function of the maximum growth, which is affected by temperature sum (degree days with +5 C° threshold), soil water availability, light conditions, nitrogen supply and atmospheric carbon dioxide (CO2) concentration. Furthermore, the growth is affected by initial diameter of trees and genotype. The tree height, and mass (or carbon) of foliage, branches, stem, and roots (carbon content 50% of mass), are calculated based on tree diameter. The probability of tree mortality (and further the amount of dead wood) is affected by stand basal area and density, and the maximum age of trees, as well as by very low diameter growth for two consecutive years.
The site fertility and regional mean temperature sum under current climate determine the initial amount of soil organic matter (carbon) and nitrogen (N) content (Kellomäki et al. 2008). Decay of litter and humus, as well as dead wood (on ground), affect the amount of soil carbon (50% of biomass) and nutrients available for the growth of trees, together with possible N fertilization (and atmospheric deposition of nitrogen). Soil moisture is affected by precipitation and evaporation, as well as by soil texture.
In the model, forest management control includes precommercial and commercial thinning, N fertilization and final felling with planting of seedlings (also natural regeneration) for the following production (rotation) cycle. In thinning and final felling, harvested timber assortments (pulpwood and sawlogs) are calculated using minimum stem diameters of 16.5 and 6 cm for sawlog and pulpwood, respectively. The stem volume is calculated based on height and diameter at breast height using Laasasenaho (1982) function. Model simulations are done with an area of 100 m2 and a time step of 1 year for over a 90-year period (2010–2099). Simulations are conducted with a pre-determined number of iterations (50 within our study) due to stochastic processes of birth and mortality of trees. Mean value of results for different iterations are used in data analysis.
The habitat suitability indices were calculated for dead wood demanding, forest saproxylic species groups, based on the simulated amount and type of dead wood and stand basal area. Calculations are based on Kouki and Tikkanen (2007) and Tikkanen et al. (2007) habitat suitability index (HSI) models including 27 saproxylic species group (both invertebrate and fungal). In each group, species were dependent on dead wood of the same host tree species, decay level, type (standing/fallen) and size. The availability of suitable dead wood and microclimatic conditions of the stand (indifferent, moist/shady and sunny) affected HSI values of each saproxylic species group. In the model microclimatic conditions are affected by the total basal area of the stand and the proportion of Norway spruce (Picea abies (L.) Karst.). The presence of Norway spruce increases the habitat suitability for moist and shade demanding species. If stand (plot) HSI is greater than 0.5, it is considered as suitable habitat for the species group. Area of suitable habitats (HSA) is calculated based on total area of stands with HSI averages of indifferent, moist/shady and sunny species groups greater than 0.5.
2.2 Forest data and simulations
The regional level simulations were conducted on upland mineral soils in Finland on forest land used for timber production in the province of North Karelia (62°50´N, 29°55´E) of which total surface area is 21 584 km2. The forest data used to initialize simulations was based on national forest inventory data (NFI10, data represents circular sample plots measured in stands with relascope; one sample stand of each permanent sample stand cluster), obtained from the Natural Resources Institute Finland. We used in total of 190 forest stands (sample plots) in the simulations. Initial forest composition consisted of 48% Scots pine, 27% Norway spruce, 23% silver birch and 2% other broadleaved species.
All simulations were conducted using the climate data and scenarios obtained from the Finnish Meteorological Institute for the current climate (CU) representing a period of 1981–2010 and multi-model Global Climate Model (GCM) projections of RCP2.6 and RCP4.5 forcing scenarios over a 90-year period (period of 2010–2099). Under the RCP2.6 and RCP4.5 scenarios, the mean annual temperature is expected to increase by 1.9 °C and 3.3 °C and the precipitation by 6% and 11% by the 2080s when compared to the period of 1981–2010, respectively (Ruosteenoja et al. 2016; Venäläinen et al. 2020). The climate change projections used in this study are quite similar also with the recently updated projections SSP1-2.6 and SSPs-4.5 by the Shared Socio-economic Pathways (SSPs) greenhouse gas (GHG) scenarios, with temperature and precipitation increases of 2.4–3.7 °C and 8–11% (Ruosteenoja and Jylhä 2021). We did not consider more severe climate change scenarios in our simulations, such as RCP8.5. This was because our simulation model cannot simulate natural disturbances (windstorms and spruce bark beetles), which are projected to increase strongly under RCP8.5, but also to some extent under RCP4.5, in southern and central Finland towards end of this century (Venäläinen et al. 2020).
Stand level management scenarios used in simulations were: Baseline management (BM); BM with increases in forest conservation area by 10 or 20% (CONS10, CONS20); Intensive management scenarios using CONS10 and CONS20 combined with use of improved seedlings and fertilization in managed forest stands (CONS10+INT and CONS20+INT). Inventory stands with a higher basal area and/or trees with larger diameter had a higher probability of selection for conservation in the simulations. Thinning treatments (thinning from below) were based on current Finnish forest management recommendations (Äijälä et al. 2019). The basal area weighted diameter at breast height, defined the timing of final felling.
The clear-cut sites were planted with either Scots pine (Pinus sylvestris L.) (2000 seedlings ha–1), Norway spruce (2000 seedlings ha–1), or silver birch (Betula pendula Roth) (1600 seedlings ha–1) depending on the dominant species prior to final felling of the initial stand based on inventory stands. Forest regeneration under CONS10+INT and CONS20+INT scenarios used improved seedlings with 20% better potential growth than non-improved seedlings. However, realized growth with improved seedling is an approximation and is to some extent lower (Saarsalmi and Mälkönen 2001; Ruotsalainen 2014; Jansson et al. 2017).
Nitrogen fertilization was applied using 150 kg N ha−1 at the time of thinning on upland sub-xeric (Scots pine) and mesic sites (Scots pine and Norway spruce). In managed forests, an annual N deposition of 10 kg ha−1 was applied, based on Kellomäki et al. (2005). In practice, the management recommendations are not strictly followed and there exist harvest savings in the forests. To avoid harvesting of all harvest savings in the first years of simulations, we used a mean delay of 13 years in the thinning and final felling as was done in previous simulation studies by Alrahahleh et al. (2016, 2018a). The lack of initial amount of dead wood in forest inventory data affected the simulated amount of dead wood and further habitat suitability area over time.
2.3 Data analyses
Based on the forest ecosystem model simulations, we analyzed how timber yield (m3 ha–1), ecosystem carbon stock (trees and soil, Mg ha–1), amount of dead wood (standing and fallen, Mg ha–1), and total area of suitable habitat for saproxylic species were affected by forest conservation and management intensity, in addition to climate change, over a 90-years simulation period (periods of 2010–2039, 2040–2069 and 2070–2099). The results of management scenarios were compared to the BM. Similarly, the results under RCP2.6 and RCP4.5 were compared to CU.
3 Results
3.1 Harvested timber yield
Under the current climate (CU), timber yield decreased in the first 30-year period among different forest conservation scenarios by 17–38% compared to BM, the least in CONS10+INT and the most under CONS20. In the second and third 30-years periods, the corresponding decreases were in a range of 11–32% and 13–20%, respectively, excluding the CONS10+INT in the third 30-years period, in which timber yield was 5% higher compared to BM (Fig. 1).
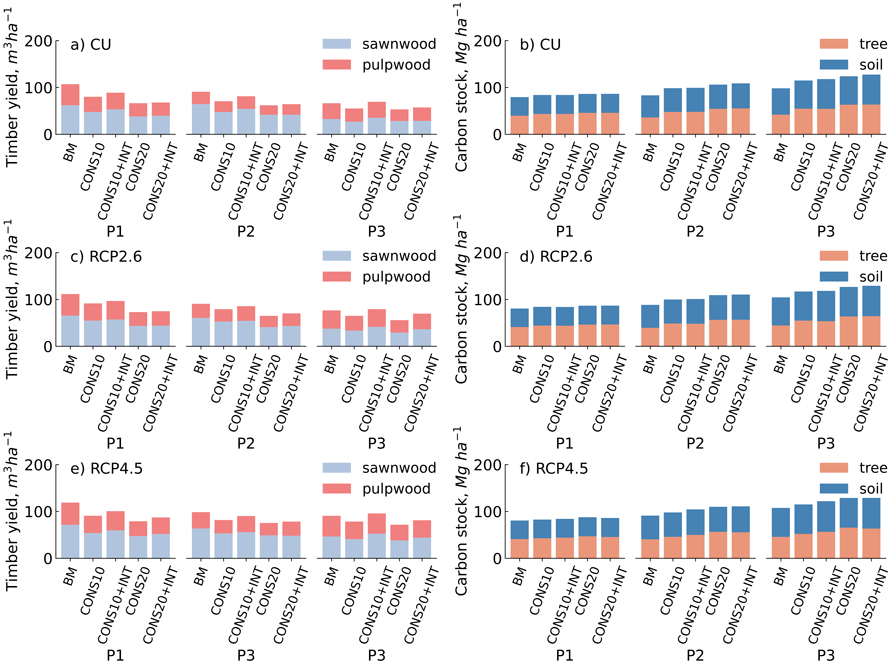
Fig. 1. The total timber yield (m3 ha–1) (a,c,e) and ecosystem carbon stock (Mg ha–1) (b,d,f) over three 30-year periods (P1,P2,P3) for different management scenarios under the current climate (CU), Representative Concentration Pathway RCP2.6 and RCP4.5 climate change projections in eastern Finland.
Under changing climates represented by the RCPs, the change in conservation area and/or management intensity affected timber yield in a similar way as under CU. Under RCP2.6 and RCP4.5, timber yield decreased in the first 30-year-period under different forest conservation scenarios by 13–35% and 15–34%, respectively, compared to BM, the least in CONS10+INT and the most with CONS20. In the second and third 30-year-periods, the decreases under RCP2.6 were in a range of 6–28% and 9–27%, while under RCP4.5 decreases of 9–23% and 11–13% occurred, under different conservation scenarios (Excluding CONS10+INT), respectively. However, under RCPs, 4–6% increases in timber yield were found in CONS10+INT under the third 30-years period compared to BM (Fig. 1).
Under the RCP2.6, timber yield was in the first, second and third 30-year-periods 4–14%, 0–12%, and 5–18% higher compared to CU, respectively, depending on conservation and management intensity. Under the RCP4.5, the corresponding increases were 11–33%, 9–26% and 35–49%, respectively, compared to CU (Fig. 1).
3.2 Forest ecosystem carbon stock
Under the current climate, forest ecosystem carbon stocks (trees and soil) increased in the first 30-year-period by 6% in both CONS10 and CONS10+INT scenarios and 9% in both CONS20 and CONS20+INT scenarios compared to BM. In the second and third 30-year-periods, the corresponding increases were in a range of 18–31% and 17–30%, respectively, the least in CONS10 and the most under the CONS20+INT (Fig. 1).
Under the RCPs, carbon stocks increased in the first 30-year-period across all forest conservation scenarios by 3–9%, respectively, the least in CONS10 and CONS10+INT scenarios and the most in CONS20 and CONS20+INT scenarios compared to the BM. In the second and third 30-year-periods, the increases were under RCP2.6 in a range of 13–25% and 12–24% and under RCP4.5 in a range of 8–22% and 7–20%, respectively, the least in CONS10 and the most in CONS20+INT (Fig. 1).
Under RCP2.6 the carbon stocks were in the first, second and third 30-year-periods in range of –2–1%, 1–6%, and 0–6% lower/higher compared to CU, respectively, depending on conservation and management intensity. Under RCP4.5, the corresponding changes were –2–2%, 0–10% and 0–10%, respectively, compared to the current climate (Fig. 1).
3.3 Amount of dead wood and area of suitable habitats for saproxylic species
On average, the share of fallen dead wood from the total amount of dead wood (fallen + standing) decreased from 54% to 34% over study period under all management and climate scenarios. Under the CU, the total amount of dead wood increased in the first 30-year-period across all forest conservation scenarios by 10–19%, the least in CONS10+INT and the most under the CONS20+INT scenarios compared to BM. In the second and third 30-year periods, the corresponding increases were in a range of 20–39% and 22–35%, respectively, the least in CONS10+INT (second period) and CONS10 (third period) and the most under CONS20+INT (Fig. 2).
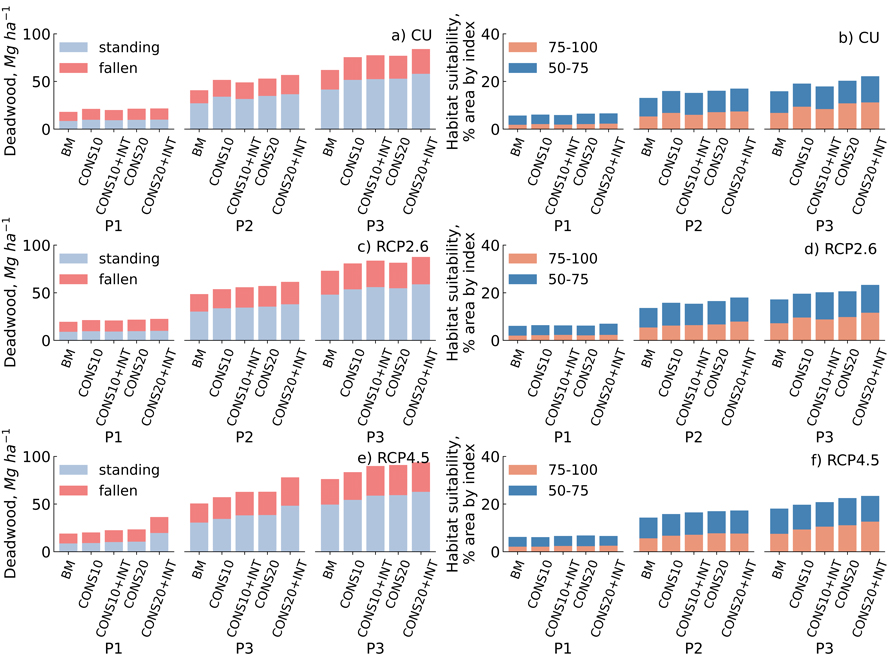
Fig. 2. The total amount of dead wood (Mg ha–1) (a,c,e) and suitable habitat area for saproxylic species (%) (b,d,f) over three 30-year simulation period (P1,P2,P3) for different management scenarios under the current climate (CU), RCP2.6 and RCP4.5 climate change projections in central Finland. (50–75) Means that the habitat is suitable and (75–100) means that the habitat is very suitable for the species.
Under RCP2.6 and RCP4.5, the total amount of dead wood increased in the first 30-year-period under conservation scenarios by 7–16% and 6–92%, respectively, the least in CONS10+INT (for RCP2.6) and CONS10 (for RCP4.5) and the most under CONS20+INT compared to BM. In the second and third 30-year-periods, the corresponding increases under RCP2.6 were in a range of 11–26% and 11–20% and under RCP4.5 in a range of 13–54% and 10–23%, respectively, the least in CONS10 and the most in CONS20+INT (Fig. 2).
Under RCP2.6 the total amount of dead wood was in the first, second and third 30-year-periods in range of 1–7%, 4–19%, and 4–18% higher compared to CU, respectively, depending on conservation and management intensity. Under RCP4.5, the corresponding changes were –5–68%, 11–37% and 11–23%, respectively, compared to CU (Fig. 2).
3.4 Area of suitable habitats for saproxylic species
Under the current climate, the habitat suitability area increased in the first 30-year-period across all forest conservation scenarios by 3–16%, the least in CONS10+INT and the most under CONS20 and CONS20+INT when compared to BM. In the second and third 30-year periods, the corresponding increases were in a range of 16–30% and 13–40%, respectively, the least in CONS10+INT and the most under CONS20+INT (Fig. 2).
Under RCP2.6, habitat suitability area increased in the first 30-year-period in forest conservation scenarios by 4–16%, the least in CONS20 and the most under the CONS20+INT compared to BM. Correspondingly, under RCP4.5, the change in habitat suitability area was in a range of –2–9%, the lowest under CONS10 and highest under CONS20 when compared to BM. In the second and third 30-year-periods, the habitat suitability area increased under RCP2.6 in a range of 14–33% and 14–36%, respectively, the least in CONS10 scenarios and the most under CONS20+INT. Correspondingly under RCP4.5 the increase was in the second and third period in a range of 11–21% and 9–30%, respectively, the least in CONS10 and the most under CONS20+INT (Fig. 2).
Under RCP2.6 the habitat suitability area was in the first, second and third 30-year-periods in range of –5–8%, –1–6%, and 2–13% lower or higher compared to CU, respectively, depending on conservation and management intensity. Under RCP4.5, the corresponding changes were –1–13%, 0–9% and 3–16%, respectively, compared to the CU (Fig. 2).
4 Discussion
Previous studies have shown that timber yield usually compete with other ecosystem services (Schwenk et al. 2012; Triviño et al. 2015; Alrahahleh et al. 2016; Pohjanmies et al. 2017; Díaz-Yáñez et al. 2020). Our results from solely increasing conservation area align with previous studies, displaying competing ecosystem services through decreases in timber yield on average by 15–30%. However, compared to baseline forest management, the integration of enhanced forest regeneration materials and nitrogen fertilization, coupled with expanded conservation areas, resulted in reduction in timber yield on average from 5% to 26%. Intensive management did not fully compensate the loss of timber within the first and second period. However, by the third period, the CONS10+INT scenario was able to increase timber yield by 4–6% across all climates when compared to BM.
Heinonen et al. (2018b) and Routa et al. (2019) found the use of improved regeneration material and N fertilization to increase long-term timber yield, similar to our findings. Based on Heinonen et al. (2018b), at a regional level, the benefits of forest fertilization will realize quicker and be also higher than those of improved regeneration material. This is because the latter one is dependent on the area of forest at the final felling phase, whereas fertilization may be done even a couple of times over a rotation. Whereas in individual stands their simultaneous use will increase timber yield remarkably (Routa et al. 2019). The effect of improved regeneration material in the study of Heinonen et al. (2018b) was more pronounced in southern Finland, while fertilization had a larger effect in central and northern Finland. These regional differences may partially be explained by differences in forest structure, which affects areas of regeneration and fertilization. In our simulations, we did not employ management scenarios that solely included use of forest fertilization or improved forest regeneration.
Under mild (RCP2.6) and moderate (RCP4.5) climate change, timber yield increased compared to the current climate over the study period by on average 7–15% and 19–36%, respectively. The harvested amount of timber in the first period was much larger compared to other periods, as large amount of harvesting savings were harvested at the beginning of the simulations. In a simulation study Heinonen et al. (2018a), it was observed that under mild (RCP2.6) and moderate (RCP4.5) climate change with intensified forest management (improved regeneration material, fertilization, and ditch network maintenance) the maximum sustainable harvest level in Finland would be up to 80 million m3 yr–1 over next 100 years. As of 2021, total annual roundwood removals in Finland were approximately 76 million m3 (Kulju et al. 2023). In previous simulation studies, use of more severe RCP-scenarios (e.g. RCP8.5 by 2100) were found to reduce the volume growth of forests due to increase in tree mortality, as growing conditions become suboptimal for conifer species, especially in southern Finland (Alrahahleh et al. 2018a, 2018b; Heinonen et al. 2018a; Kellomäki et al. 2018).
When examining ecosystem carbon stocks, increases in forest conservation area increased carbon stocks by on average 6–21%, while intensive management slightly enhanced this effect (on average, 11–23%). Increase in soil carbon stocks can be explained by the increased forest growth and production of litter and its accumulation in the soil (Lindroos et al. 2023; Mäkipää et al. 2023). However, in our study climate change had no clear impact on carbon stocks in the forest (varying by 1 to 7%), because increased growth due to climate change enhanced production of timber and its harvest.
In North Karelia, approximately 51 000 ha (3%) of forest land and 8000 ha (18%) of poorly productive forests are strictly protected. Increasing the area of forest land under strict protection to 10% in each Finnish province, favouring the oldest forests, would mean an increase of 60 000 ha in North Karelia (Kotiaho et al. 2021). In the study of Kniivilä et al. (2022) 10% strict protection in each Finnish province (allocated to old forests) resulted in significant reduction of harvesting opportunities, especially in North Karelia. In their study the simulated use of timber and wood for energy either exceeded or was very close to the estimate of the maximum sustained harvesting level in Finland (Maanavilja et al. 2021; Kniivilä et al. 2022). Therefore, based on our simulations, use of improved regeneration material and fertilization, as represented by intensive management scenarios could help in reducing the timber deficit due to increased conservation area and, in the long term, even achieve better timber yield compared to current forest management.
The total amount of available dead wood per period was relatively low, particularly at the start of the simulations, as the model did not consider dead wood already present. As forest conservation area increased, the amount of dead wood (DW) rose by on average 10–24% and the area of suitable habitat for saproxylic species (HSA) increased (on average, 8–24%) compared to baseline forest management over the study period. In conservation areas, self-thinning increases tree mortality and thereby also the amount of dead wood (Tikkanen et al. 2007; Mazziotta et al. 2014). The use of improved regeneration material and fertilization enhanced this effect in managed forest stands (on average, 12–56% for DW, 12–32% for HSA). Compared to the current climate, the amount of dead wood and the area of suitable habitats increased under warmer climates (on average, 4–39% for DW, 1–13% for HSA) and therefore forest management had a higher impact on the amount of dead wood and habitat suitability area compared to climate change.
In cases where there is a small amount of dead wood, additional increases of dead wood will improve biodiversity values significantly until a certain point, after which biodiversity will no longer benefit from additional increases of dead wood (Díaz-Yáñez et al. 2021). However, biodiversity is also influenced by tree species composition in forest stands; for species requiring spruce dead wood, substantial amount of pine dead wood does not create an optimal environment and vice versa, especially when the habitat suitability index (HSI) models favor spruce dependent species (Tikkanen et al. 2006). The habitat suitability area in our study did not increase at the same rate as the total amount of dead wood under RCP4.5. According to Mazziotta et al. (2016) vulnerability of saproxylic species to climate change is greatly dependent on species-habitat associations. Climate change is expected to increase the availability of dead wood, but increased decomposition rate may further reduce availability of dead wood for saproxylic species (Mazziotta et al. 2014, 2016).
The diversity and future availability of dead wood are more important than the total dead wood volume (Similä et al. 2003; Lassauce et al. 2011). In our study, the amount of fallen dead wood from total amount of dead wood decreased from half to a third and the amount of very suitable habitat area (75‒100) from total amount of suitable habitat areas (50‒100) increased from one third to almost half over the study periods in all management and climate scenarios. This is understandable, if the amount of dead wood for saproxylic species decreased in poor sites and increased in ideal sites. For example, in the optimizations of Tikkanen et al. (2007) HSI values differentiated depending on the use of the forest stand (conservation or wood production). The quality of the matrix in managed and conservation forests is important, therefore should be chosen efficiently (Lindenmayer and Franklin 2002; Juutinen et al. 2004; Tikkanen et al. 2007; Mönkkönen et al. 2014).
Our results show that slight increase in conservation area in conjunction with intensive forest management (use of improved growing stock and fertilization) may increase all the studied ecosystem variables over the rotation period of forest stands. However, our simulations could not consider multiple natural disturbances, which are expected to increase along with the severity of climate change (Reyer et al. 2017; Seidl et al. 2017; Netherer et al. 2019; Venäläinen et al. 2020). Increasing natural disturbances, such as windstorms and spruce bark beetles in southern and central Finland, and especially under RCP8.5 but also to some extent under RCP4.5, affect also provisioning of ecosystem services (Venäläinen et al. 2020). In addition, our study used only saproxylic species as biodiversity indicator and recognized the limitations of the analyses based on calculated HSI averages of indifferent, moist/shady and sunny species groups.
Rotational forestry is the most common forest management method in Finland (Mason et al. 2022), and therefore it was used in our study. However, continuous cover forestry relies on natural regeneration and thus the benefits of improved forest regeneration material are not obtained. Use of fertilization may result in adverse environmental impacts both in rotational and continuous cover forestry, e.g. through changes in species and organism composition of forest ecosystem (Strengbom and Nordin 2008; Högberg et al. 2010; Entwistle et al. 2018; Strengbom et al. 2018; Jörgensen et al. 2021). Too intensive fertilization may also increase greenhouse gas (GHG) emissions and nutrient export to water courses (Bergh et al. 2008; Hedwall et al. 2014; Ojanen et al. 2019; Minkkinen et al. 2020; Håkansson et al. 2021). Typically, the fertilization dose has been about 150 kg N ha–1 in Scots pine and Norway spruce stands in practical forestry, i.e. as we used in model simulations (Hedwall et al. 2014).
5 Conclusion
Trade-offs exist between the harvested amount of timber and other ecosystem services utilized in this study. No single management scenario allowed for maximizing all study variables simultaneously. The increase in forest conservation area resulted in higher ecosystem carbon stocks, quantities of dead wood and area of suitable habitats for saproxylic species, but a lower amount of harvested timber. The combined use of improved regeneration material and fertilization enhanced all studied variables and reduced the timber yield loss in the conservation simulations. As an exception to other simulation cases, the 10% forest conservation scenario with intensified forest management increased the benefits of ecosystem services and even increased the harvested amount of timber compared to baseline forest management in the last 30-year period. In addition, the warming climate increased timber yield, amount of dead wood and through it also habitat suitability area for saproxylic species.
Declaration on the availability of research materials
Data available on request from the corresponding author.
Authors’ contributions
Conceptualization (HP), data curation (HS), data analysis (LP, HS, EV), writing – original draft preparation (LP, EV), writing – Review & Editing (AP, O-PT, AK, HP), visualization (LP, AP), project administration (HP). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Academy of Finland [grant numbers 337127 and 357906 for UNITE flagship].
Acknowledgement
The National Forest Inventory data is provided by the Natural Resources Institute Finland and climatic data (for current and changing climate scenarios) is provided by the Finnish Meteorological Institute.
References
Äijälä O, Koistinen A, Sved J, Vanhatalo K, Väisänen P (2019) Metsänhoidon suositukset- METSÄNHOITO. [Finnish forest management recommendations]. Metsätalouden kehittämiskeskus Tapion julkaisuja. ISBN 978-952-5632-75-0.
Alrahahleh L, Ikonen V-P, Kilpeläinen A, Torssonen P, Strandman H, Asikainen A, Kaurola J, Venäläinen A, Peltola H (2016) Effects of forest conservation and management on volume growth, harvested amount of timber, carbon stock, and amount of dead wood in Finnish boreal forests under changing climate. Can J For Res 47: 215–225. https://doi.org/10.1139/cjfr-2016-0153.
Alrahahleh L, Kilpeläinen A, Ikonen V-P, Strandman H, Asikainen A, Venäläinen A, Kaurola J, Kangas J, Peltola H (2018a) Effects of using certain tree species in forest regeneration on volume growth, timber yield, and carbon stock of boreal forests in Finland under different CMIP5 projections. Eur J For Res 137: 573–591. https://doi.org/10.1007/s10342-018-1126-z.
Alrahahleh L, Kilpeläinen A, Ikonen V-P, Strandman H, Venäläinen A, Peltola H (2018b) Effects of CMIP5 projections on volume growth, carbon stock and timber yield in managed scots pine, Norway spruce and silver birch stands under southern and northern boreal conditions. Forests 9, article id 208. https://doi.org/10.3390/f9040208.
Bergh J, Nilsson U, Grip H, Hedwall O, Lundmark T (2008) Effects of frequency of fertilisation on production, foliar chemistry and nutrient leaching in young Norway spruce stands in Sweden. Silva Fenn 42: 721–733. https://doi.org/10.14214/sf.225.
Díaz-Yáñez O, Pukkala T, Packalen P, Peltola H (2020) Multifunctional comparison of different management strategies in boreal forests. Forestry 93: 84–95. https://doi.org/10.1093/forestry/cpz053.
Díaz-Yáñez O, Pukkala T, Packalen P, Lexer MJ, Peltola H (2021) Multi-objective forestry increases the production of ecosystem services. Forestry 94: 386–394. https://doi.org/10.1093/forestry/cpaa041.
Entwistle EM, Romanowicz KJ, Argiroff WA, Freedman ZB, Morris JJ, Zak DR (2018) Anthropogenic N deposition alters the composition of expressed class II fungal peroxidases. Appl Environ 84, article id e02816-17. https://doi.org/10.1128/AEM.02816-17.
European commission (2020) Communication from the commission to the European parliament, the council, the European economic and social committee and the committee of the regions. EU Biodiversity Strategy for 2030. Bringing nature back into our lives. Brussel 20.5.2020. COM(2020) 380 final. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A52020DC0380.
European commission (2022) Criteria and guidance for protected areas designations – staff working document. Directorate-General for Environment. Brussels 28.1.2022. SWD(2022) 23 final. https://environment.ec.europa.eu/publications/criteria-and-guidance-protected-areas-designations-staff-working-document_en.
Felton A, Gustafsson L, Roberge J-M, Ranius T, Hjältén J, Rudolphi J, Lindbladh M, Weslien J, Rist L, Brunet J, Felton AM (2016) How climate change adaptation and mitigation strategies can threaten or enhance the biodiversity of production forests: insights from Sweden. Biol Conserv 194: 11–20. https://doi.org/10.1016/j.biocon.2015.11.030.
Gauthier S, Bernier P, Kuuluvainen T, Shvidenko AZ, Schepaschenko DG (2015) Boreal forest health and global change. Sci 349: 819–822. https://doi.org/10.1126/science.aaa9092.
Gossner MM, Lachat T, Brunet J, Isacsson G, Bouget C, Brustel H, Brandl R, Weisser WW, Mueller J (2013) Current near-to-nature forest management effects on functional trait composition of saproxylic beetles in beech forests. Conserv Biol 27: 605–614. https://doi.org/10.1111/cobi.12023.
Håkansson C, Hedwall P-O, Strömgren M, Axelsson M, Bergh J (2021) Effects of fertilization on soil CH4 and N2O fluxes in young Norway spruce stands. For Ecol Manag 499, article id 119610. https://doi.org/10.1016/j.foreco.2021.119610.
Hedwall P-O, Gong P, Ingerslev M, Bergh J (2014) Fertilization in northern forests – biological, economic and environmental constraints and possibilities. Scand J Forest Res 29: 301–311. https://doi.org/10.1080/02827581.2014.926096.
Heinonen T, Pukkala T, Mehtätalo L, Asikainen A, Kangas J, Peltola H (2017) Scenario analyses for the effects of harvesting intensity on development of forest resources, timber supply, carbon balance and biodiversity of Finnish forestry. For Policy Econ 80: 80–98. https://doi.org/10.1016/j.forpol.2017.03.011.
Heinonen T, Pukkala T, Kellomäki S, Strandman H, Asikainen A, Venäläinen A, Peltola H (2018a) Effects of forest management and harvesting intensity on the timber supply from Finnish forests in a changing climate. Can J For Res 48: 1124–1134. https://doi.org/10.1139/cjfr-2018-0118.
Heinonen T, Pukkala T, Asikainen A, Peltola H (2018b) Scenario analyses on the effects of fertilization, improved regeneration material, and ditch network maintenance on timber production of Finnish forests. Eur J Forest Res 137: 93–107. https://doi.org/10.1007/s10342-017-1093-9.
Hetemäki L, Kangas J, Peltola H (eds) (2022) Forest bioeconomy and climate change. Managing Forest Ecosystems 42, Springer Cham. https://doi.org/10.1007/978-3-030-99206-4.
Högberg MN, Briones MJI, Keel SG, Metcalfe DB, Campbell C, Midwood AJ, Thornton B, Hurry V, Linder S, Näsholm T, Högberg P (2010) Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytol 187: 485–493. https://doi.org/10.1111/j.1469-8137.2010.03274.x.
Hynynen J, Salminen H, Ahtikoski A, Huuskonen S, Ojansuu R, Siipilehto J, Lehtonen M, Eerikäinen K (2015) Long-term impacts of forest management on biomass supply and forest resource development: a scenario analysis for Finland. Eur J Forest Res 134: 415–431. https://doi.org/10.1007/s10342-014-0860-0.
Hyvärinen E, Juslén A, Kemppainen E, Uddström A, Liukko U-M (eds) (2019) The 2019 Red List of Finnish species. Ympäristöministeriö & Suomen ympäristökeskus, Helsinki. http://hdl.handle.net/10138/299501.
Jansson G, Kehlet Hansen J, Haapanen M, Kvaalen H, Steffenrem A (2017) The genetic and economic gains from forest tree breeding programmes in Scandinavia and Finland. Scand J For Res 32: 273–286. https://doi.org/10.1080/02827581.2016.1242770.
Jörgensen K, Granath G, Lindahl BD, Strengbom J (2021) Forest management to increase carbon sequestration in boreal Pinus sylvestris forests. Plant Soil 466: 165–178. https://doi.org/10.1007/s11104-021-05038-0.
Junninen K, Komonen A (2011) Conservation ecology of boreal polypores: a review. Biol Conserv 144: 11–20. https://doi.org/10.1016/j.biocon.2010.07.010.
Juutinen A, Mäntymaa E, Mönkkönen M, Salmi S (2004) A cost-efficient approach to selecting forest stands for conserving species: a case study from Northern Fennoscandia. For Sci 50: 527–539.
Kellomäki S, Strandman H, Nuutinen T, Peltola H, Korhonen KT, Väisänen H (2005) Adaptation of forest ecosystems, forests and forestry to climate change. FINADAPT Working Paper 4, Finnish Environmental Institute Mimeographs 334, Helsinki.
Kellomäki S, Peltola H, Nuutinen T, Korhonen KT, Strandman H (2008) Sensitivity of managed boreal forests in Finland to climate change, with implications for adaptive management. Phil Trans R Soc B363: 2339–2349. http://doi.org/10.1098/rstb.2007.2204.
Kellomäki S, Strandman H, Heinonen T, Asikainen A, Venäläinen A, Peltola H (2018) Temporal and spatial change in diameter growth of boreal Scots pine, Norway spruce and birch under recent-generation (CMIP5) global climate model projections for the 21st century. Forests 9, article id 118. https://doi.org/10.3390/f9030118.
Kniivilä M, Hirvelä H, Lintunen J, Mutanen A, Vatanen E, Viitanen J, Kurttila M (2022) Metsien tiukan lisäsuojelun hakkuumahdollisuus-, arvonlisäys- ja työllisyysvaikutusten arviointi: Skenaariotarkastelu EU:n biodiversiteettistrategiasta Suomessa. [Assessment of the harvesting, value increase and employment impact of the additional strict forest protection: Scenario review of the EU Biodiversity Strategy in Finland]. Luonnonvara- ja biotalouden tutkimus 64/2022. Luonnonvarakeskus, Helsinki. http://urn.fi/URN:ISBN:978-952-380-480-7.
Korhonen KT, Ihalainen A, Kuusela S, Punttila P, Salminen O, Syrjänen K (2020) Metsien monimuotoisuudelle merkittävien rakennepiirteiden muutokset Suomessa vuosina 1980–2015. [Changes in significant structural features for forest biodiversity in Finland in 1980–2015]. Metsätieteen aikakauskirja, article id 10198. https://doi.org/10.14214/ma.10198.
Kotiaho JS, Ahlvik L, Bäck J, Hohti J, Jokimäki J, Kallio KP, Ketola T, Kulmala L, Lakka H-K, Lehikoinen A, Oksanen E, Pappila M, Sääksjärvi I, Peura M (2021) Metsäluonnon turvaava suojelun kohdentaminen. [Allocation of forest protection in Finland]. Suomen Luontopaneelin julkaisuja 4/2021.
Kouki J, Tikkanen OP (eds) (2007) Uhanalaisten lahopuulajien elinympäristöjen turvaaminen suojelualueilla ja talousmetsissä. [Securing habitats for threatened saproxylic species in protected and managed forests]. Suomen Ympäristö – The Finnish Environment 24/2007, Ympäristöministeriö, Helsinki,
Kulju I, Niinistö T, Peltola A, Räty M, Sauvula-Seppälä T, Torvelainen J, Uotila E, Vaahtera E (2023) Finnish statistical yearbook of forestry 2022. Luonnonvarakeskus, Helsinki. http://urn.fi/URN:ISBN:978-952-380-584-2.
Laasasenaho J (1982) Taper curve and volume functions for pine, spruce and birch. Commun Inst for Fenn 108. http://urn.fi/URN:ISBN:951-40-0589-9.
Lassauce A, Paillet Y, Jactel H, Bouget C (2011) Dead wood as a surrogate for forest biodiversity: meta-analysis of correlations between dead wood volume and species richness of saproxylic organisms. Ecol Ind 11: 1027–1039. https://doi.org/10.1016/j.ecolind.2011.02.004.
Lindenmayer DB, Franklin JK (2002) Conserving forest biodiversity – a comprehensive multiscaled approach. Island Press.
Lindroos AJ, Mäkipää R, Merilä P (2023) Soil carbon stock changes over 21 years in intensively monitored boreal forest stands in Finland. Ecol Ind 144, article id 109551. https://doi.org/10.1016/j.ecolind.2022.109551.
Maanavilja L, Tuomainen T, Aakkula J, Haakana M, Heikkinen J, Hirvelä H, Kilpeläinen H, Koikkalainen K, Kärkkäinen L, Lehtonen H, Miettinen A, Mutanen A, Myllykangas JP, Ollila P, Viitanen J, Vikfors S, Wall A (2021) Hiilineutraali Suomi 2035: Maankäyttö- ja maataloussektorin skenaariot. [Carbon neutral Finland 2035 – scenarios for LULUCF and agriculture Sectors]. Valtioneuvoston selvitys- ja tutkimustoiminnan julkaisusarja 2021:63, Valtioneuvoston kanslia. http://urn.fi/URN:ISBN:978-952-383-263-3.
Mäkipää R, Abramoff R, Adamczyk B, Baldy V, Biryol C, Bosela M, Casals P, Curiel Yuste J, Dondini M, Filipek S, Garcia-Pausas J, Gros R, Gömöryová E, Hashimoto S, Hassegawa M, Immonen P, Laiho R, Li H, Li Q, Luyssaert S, Menival C, Mori T, Naudts K, Santonja M, Smolander A, Toriyama J, Tupek B, Ubeda X, Verkerk PJ, Lehtonen A (2023) How does management affect soil C sequestration and greenhouse gas fluxes in boreal and temperate forests? – A review. For Ecol Manag 529, article id 120637. https://doi.org/10.1016/j.foreco.2022.120637.
Mason WL, Diaci J, Carvalho J, Valkonen S (2022) Continuous cover forestry in Europe: usage and the knowledge gaps and challenges to wider adoption. Forestry 95: 1–12. https://doi.org/10.1093/forestry/cpab038.
Mazziotta A, Mönkkönen M, Strandman H, Routa J, Tikkanen O-P, Kellomäki S (2014) Modeling the effects of climate change and management on the dead wood dynamics in boreal forest plantations. Eur J Forest Res 133: 405–421. https://doi.org/10.1007/s10342-013-0773-3.
Mazziotta A, Triviño M, Tikkanen O-P, Kouki J, Strandman H, Mönkkönen M (2016) Habitat associations drive species vulnerability to climate change in boreal forests. Clim Change 135: 585–595. https://doi.org/10.1007/s10584-015-1591-z.
Minkkinen K, Ojanen P, Koskinen M, Penttilä T (2020) Nitrous oxide emissions of undrained, forestry-drained, and rewetted boreal peatlands. For Ecol Manag 478, article id 118494. https://doi.org/10.1016/j.foreco.2020.118494.
Mönkkönen M, Juutinen A, Mazziotta A, Miettinen K, Podkopaev D, Reunanen P, Salminen H, Tikkanen O-P (2014) Spatially dynamic forest management to sustain biodiversity and economic returns. J Environ Manage 134: 80–89. https://doi.org/10.1016/j.jenvman.2013.12.021.
Müller J, Bütler R (2010) A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur J For Res 129: 981–992. https://doi.org/10.1007/s10342-010-0400-5.
Netherer S, Panassiti B, Pennerstorfer J, Matthews B (2019) Acute drought is an important driver of bark beetle infestation in Austrian Norway spruce stands. Front For Global Change 2, article id 39. https://doi.org/10.3389/ffgc.2019.00039.
Ojanen P, Penttilä T, Tolvanen A, Hotanen J-P, Saarimaa M, Nousiainen H, Minkkinen K (2019) Long-term effect of fertilization on the greenhouse gas exchange of low-productive peatland forests For Ecol Manag 432: 786–798. https://doi.org/10.1016/j.foreco.2018.10.015.
Penttilä R, Siitonen J, Kuusinen M (2004) Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol Conserv 117: 271–283. https://doi.org/10.1016/j.biocon.2003.12.007.
Peuhu E, Siitonen J (2011) Ontot puistopuut ovat merkittävä elinympäristö monimuotoiselle lahopuueliöstölle. [Hollow park trees are a significant habitat for diverse saproxylic species]. Metsätieteen aikakausikirja 1/2011: 63–67. https://doi.org/10.14214/ma.5935.
Pohjanmies T, Triviño M, Le Tortorec E, Salminen H, Mönkkönen M (2017) Conflicting objectives in production forests pose a challenge for forest management. Ecosyst Serv 28: 298–310. https://doi.org/10.1016/j.ecoser.2017.06.018.
Poudel BC, Sathre R, Gustavsson L, Bergh J, Lundström A, Hyvönen R (2011) Effects of climate change on biomass production and substitution in north-central Sweden. Biomass Bioenergy 35: 4340–4355. https://doi.org/10.1016/j.biombioe.2011.08.005.
Poudel BC, Sathre R, Bergh J, Gustavsson L, Lundström A, Hyvönen R (2012) Potential effects of intensive forestry on biomass production and total carbon balance in north-central Sweden. Environ Sci Policy 15: 106–124. https://doi.org/10.1016/j.envsci.2011.09.005.
Pukkala T (2017) Does management improve the carbon balance of forestry? Forestry 90: 125–135. https://doi.org/10.1093/forestry/cpw043.
Räty M, Juutinen A, Korhonen KT, Syrjänen K, Kärkkäinen L (2023) EU wood production vs. biodiversity goals – possible reconciliation in Finland? Scand J For Res 38: 287–299. https://doi.org/10.1080/02827581.2023.2229732.
Reyer C, Bathgate S, Blennow K, Borges JG, Bugmann H, Delzon S, Faias SP, Garcia-Gonzalo J, Gardiner B, Gonzalez-Olabarria JR, Gracia C, Hernández JG, Kellomäki S, Kramer K, Lexer MJ, Lindner M, van der Maaten E, Maroschek M, Muys B, Nicoll B, Palahi M, Palma JHN, Paulo JA, Peltola H, Pukkala T, Rammer W, Ray D, Sabaté S, Schelhaas M-J, Seidl R, Temperli C, Tomé M, Yousefpour R, Zimmermann NE, Hanewinkel M (2017) Are forest disturbances amplifying or cancellingg out climate change-induced productivity changes in European forests? Environ Res Lett 12, article id 034027. https://doi.org/10.1088/1748-9326/aa5ef1.
Routa J, Kilpeläinen A, Ikonen V-P, Asikainen A, Venäläinen A, Peltola H (2019) Effects of intensified silviculture on timber production and its economic profitability in boreal Norway spruce and Scots pine stands under changing climatic conditions. Forestry 92: 648–658. https://doi.org/10.1093/forestry/cpz043.
Ruosteenoja K, Jylhä K (2021) Projected climate change in Finland during the 21st century calculated from CMIP6 model simulations. Geophys 56: 39–69.
Ruosteenoja K, Jylhä K, Kämäräinen M (2016) Climate projections for Finland under the RCP forcing scenarios. Geophys 51: 17–50.
Ruotsalainen S (2014) Increased forest production through forest tree breeding. Scand J For Res 29: 333–344. https://doi.org/10.1080/02827581.2014.926100.
Saarsalmi A, Mälkönen E (2001) Forest fertilization research in Finland: a literature review. Scand J Forest Res 16: 514–535. https://doi.org/10.1080/02827580152699358.
Schwenk WS, Donovan TM, Keeton WS, Nunery JS (2012) Carbon storage, timber production, and biodiversity: comparing ecosystem services with multi-criteria decision analysis. Ecol Soc Am 22: 1612–1627. https://doi.org/10.1890/11-0864.1.
Seidl R, Rammer W, Jäger D, Currie WS, Lexer MJ (2007) Assessing trade-offs between carbon sequestration and timber production within a framework of multi-purpose forestry in Austria. For Ecol Manag 248: 64–79. https://doi.org/10.1016/j.foreco.2007.02.035.
Seidl R, Schelhaas M-J, Rammer W, Verkerk PJ (2014). Increasing forest disturbances in Europe and their impact on carbon storage. Nat Clim Change 4: 806–810. https://doi.org/10.1038/nclimate2318.
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, Wild J, Ascoli D, Petr M, Honkaniemi J, Lexer MJ, Trotsiuk V, Mairota P, Svoboda M, Fabrika M, Nagel TA, Reyer CPO (2017) Forest disturbances under climate change. Nat Clim Change 7: 395–402. https://doi.org/10.1038/nclimate3303.
Similä M, Kouki J, Martikainen P (2003) Saproxylic beetles in managed and seminatural Scots pine forests: quality of dead wood matters. For Ecol Manag 174: 365–381. https://doi.org/10.1016/S0378-1127(02)00061-0.
Strengbom J, Nordin A (2008) Commercial forest fertilization causes long-term residual effects in ground vegetation of boreal forests. For Ecol Manag 256: 2175–2181. https://doi.org/10.1016/j.foreco.2008.08.009.
Strengbom J, Axelsson EP, Lundmark T, Nordin A (2018) Trade-offs in the multi-use potential of managed boreal forests. J Appl Ecol 55: 958–966. https://doi.org/10.1111/1365-2664.13019.
Tikkanen OP, Martikainen P, Hyvärinen E, Junninen K, Kouki J (2006) Red-listed boreal forest species of Finland: associations with forest structure, tree species, and decaying wood. Ann Zool Fenn 43: 373–383.
Tikkanen OP, Heinonen T, Kouki J, Matero J (2007) Habitat suitability models of saproxylic red-listed boreal forest species in long-term matrix management: cost-effective measures for multi-species conservation. Biol Conserv 140: 359–372. https://doi.org/10.1016/j.biocon.2007.08.020.
Triviño M, Juutinen A, Mazziotta A, Miettinen K, Podkopaev D, Reunanen P, Mönkkönen M (2015) Managing a boreal forest landscape for providing timber, storing and sequestering carbon. Ecosyst Serv 14: 179–189. https://doi.org/10.1016/j.ecoser.2015.02.003.
Triviño M, Pohjanmies T, Mazziotta A, Juutinen A, Podkopaev D, Le Tortorec E, Mönkkönen M (2017) Optimizing management to enhance multifunctionality in a boreal forest landscape. J Appl Ecol 54: 61–70. https://doi.org/10.1111/1365-2664.12790.
Triviño M, Morán-Ordoñez A, Eyvindson K, Blattert C, Burgas D, Repo A, Pohjanmies T, Brotons L, Snäll T, Mönkkönen M (2023) Future supply of boreal forest ecosystem services is driven by management rather than by climate change. Global Change Biol 29: 1484–1500. https://doi.org/10.1111/gcb.16566.
Venäläinen A, Lehtonen I, Laapas M, Ruosteenoja K, Tikkanen OP, Viiri H, Ikonen VP, Peltola H (2020) Climate change induces multiple risks to boreal forests and forestry in Finland: a literature review. Global Change Biol 26: 4178–4196. https://doi.org/10.1111/gcb.15183.
Total of 74 references.
