Anneli Viherä-Aarnio  ,
Pirkko Velling
,
Pirkko Velling
Growth, wood density and bark thickness of silver birch originating from the Baltic countries and Finland in two Finnish provenance trials
Viherä-Aarnio A., Velling P. (2017). Growth, wood density and bark thickness of silver birch originating from the Baltic countries and Finland in two Finnish provenance trials. Silva Fennica vol. 51 no. 4 article id 7731. https://doi.org/10.14214/sf.7731
Highlights
- Baltic origins of silver birch had significantly thicker bark than the Finnish ones
- In terms of wood density, no consistent difference was detected between the Baltic and Finnish origins
- Incidence of darkened core wood increased with increasing seed origin latitude
- Frost cracks were most common in south Latvian origins grown in central Finland.
Abstract
Silver birch (Betula pendula Roth) seed origins from the Baltic countries and from Finland were compared in terms of growth, wood density, bark thickness and the incidence of darkened core wood, frost cracks and decay, and the effect of seed origin latitude was examined in two Finnish provenance trials. The material consisted of 21 stand and single tree origins ranging from latitudes 54° to 63°N from the Baltic countries and Finland. The trials, measured at the age of 22 years, were located at Tuusula (60°21´N), southern Finland and at Viitasaari (63°11´N), central Finland. The Baltic origins were superior to the Finnish ones in the southern trial regarding height, whereas in central Finland the Finnish origins grew better. There was no consistent difference between the Baltic and the Finnish group of origins in wood density. Bark thickness decreased with increasing seed origin latitude. The Baltic origins had significantly thicker bark than the Finnish origins. A moderate positive correlation was detected between the seed origin latitude and the incidence of darkened core wood in the southern trial, where the darkened core wood was more common in the Finnish origins than in the Baltic ones. The highest proportion of trees with frost cracks was detected in the south-western Latvian origins growing in central Finland. Seed transfers from the Baltic would have an increasing effect on the bark thickness of birch logs, but no or only minor effects on wood density. Based on our results, there is no reason to recommend the use of non-native Baltic seed origins in Finland instead of the native locally adapted seed sources.
Keywords
Betula pendula;
provenance;
bark thickness;
frost crack;
seed transfer;
wood basic density;
wood discolouration
-
Viherä-Aarnio,
Natural Resources Institute Finland (Luke), Green technology, P.O. Box 2, FI-00791 Helsinki, Finland
E-mail
anneli.vihera-aarnio@luke.fi
- Velling, E-mail pike.velling@phnet.fi
Received 19 May 2017 Accepted 18 August 2017 Published 4 September 2017
Views 118579
Available at https://doi.org/10.14214/sf.7731 | Download PDF

1 Introduction
Silver birch (Betula pendula Roth) is a pioneer hardwood species occupying various habitats over a wide area of distribution in the temperate and cold regions of the Eurasian continent (Hulten and Fries 1986; Jonsell 2000). In Finland, silver birch is appreciated as a valuable material in both mechanical and chemical forest industries (Verkasalo et al. 2017). Large-sized birch timber is mainly used in the plywood and sawmilling industry, while sawn timber is further processed in joinery as well as for panels and parquets. Birch wood is generally considered to be a highly valuable raw material for a range of wood products (Luostarinen and Verkasalo 2000; Verkasalo et al. 2017). Small-sized birch is used in the pulp industry, which is, by volume, the most important consumer of birch wood in Finland. Nevertheless, the production of large-sized birch timber for the plywood and sawmilling industry is a central aim of forest management (Hynynen et al. 2010).
In the mechanical forest industry, the value of birch as a raw material is first of all determined by the size of the logs and the external quality of the stem, which affect the amount and quality of veneer and sawn timber obtained (Heiskanen 1966; Kärkkäinen 1986). However, wood density is also an important quality trait in mechanical processing, because it affects practically all mechanical strength properties and the hardness of birch timber (Heräjärvi 2004ab). Decay and discoloration in the wood are severe quality defects, especially in sawmilling, and soft decay also in veneer processing (Luostarinen and Verkasalo 2000). Discolorations around the pith are extremely common in planted birches (Hallaksela and Niemistö 1998). From the point of view of the pulping industry, wood density, bark content and incidence of decay are among the most important characteristics of birch wood (Tammisola et al. 1995). Higher density provides better pulp yields, but decay is detrimental to both the pulping process and the pulp quality (Tammisola et al. 1995). Trunks are debarked before pulping, but on the other hand, the content and properties of bark can be interesting also in terms of bioactive extracts, such as betulin, with beneficial pharmacological effects (Alakurtti et al. 2006; Krasutsky 2006). In general, interest in the different chemical properties of bark and wood is increasing in the industries (Verkasalo et al. 2017).
Birch wood is of medium density compared to other European hardwoods. The average basic density of silver birch wood is 500 ± 20 kg m–3 (Heräjärvi 2002 and references therein). Within a tree, the basic density decreases slightly vertically from the base to the tree top and increases horizontally from the pith outwards, i.e. the juvenile wood near the pith row is the lightest and the wood near the surface is the heaviest (Walldén 1934; Jalava 1945; Kujala 1946; Hakkila 1966; Repola 2006). The wood density depends on the proportions of fibres and vessels and the thickness of the fibre cell wall in relation to the lumen (Bhat 1983). The proportion of fibres decreases as the number of vessels and parenchyma increases from breast height to the crown (Bhat and Kärkkäinen 1981); in the radial direction, the proportion of fibres increases from pith to surface, which explains the corresponding increase in density (Bhat and Kärkkäinen 1981). The higher the age of cambium, the denser the wood produced (Hakkila 1979; Bhat 1980; Nepveu and Velling 1983). Densest wood is obtained from mature trees, with younger trees having 5–20% lighter wood (Heräjärvi 2002). Reaction (tension) wood, which is common in birches, is heavier than average normal wood because it contains fewer vessels than normal wood; in addition, the fibres are thinner and have thicker walls and a smaller lumen than in normal wood (Ollinmaa 1956).
Cultivation of birch by planting has been practiced in Finland since the 1960s (Raulo 1978) and it is based on genetically improved seeds of native origin, produced in multi-clonal polythene greenhouse seed orchards (Hagqvist 1991; Ahtikoski 2000). Practically no imported seed origins have been used for cultivation, because genetically improved seed orchard seeds of native origin and high quality have been available in abundance.
However, the free trade of forest cultivation material within the EU enables – and may give rise to – efforts to introduce birch seed from abroad, e.g. from the Baltic countries. It has even been suggested that slightly more southern seed origins should be used in forestry in order to take advantage of the lengthening growing season as a means of adaptation to climate change (Marttila et al. 2005). In fact, the yield of silver birch can be improved by transferring seed from the south within certain limits, which, on the other hand, may entail weaker external stem quality (Viherä-Aarnio and Velling 2008; Viherä-Aarnio et al. 2013) and higher risk of browsing by moose (Viherä-Aarnio and Heikkilä 2006). The wood quality traits, such as wood density and discoloration, as well as the properties of bark of non-native birch origins grown in Finland and the effects of seed transfers on these traits are, however, poorly known. Along with the increasing need for wood biomass in bio-refineries, the diverse properties of the biomass used and their characterisation are getting more and more important (Verkasalo et al. 2017). Studies on the genetic variation of wood quality have been carried out on the progeny (Velling 1979a; Nepveu and Velling 1983; Tammisola et al. 1995) and the clone level (Stener and Hedenberg 2003), but the variation on the provenance level is practically unknown.
The aim of this study was to compare Baltic and native Finnish seed origins of pulp-tree- sized silver birch grown in Finland and to examine the effects of seed origin on growth, wood density and bark thickness as well as on incidences of darkened core wood, frost cracks and decay.
2 Material and methods
2.1 Seed origins and field trials
The material for this study was collected from two silver birch provenance trials located at Tuusula, southern Finland (60°21´N, 25°02´E, 50 m a.s.l.) and at Viitasaari, central Finland (63°11´N, 26°07´E, 180 m a.s.l.) (Fig. 1). The trials included stand seed origins from Latvia (7), Estonia (2), Russia (2) and Finland (6) (Table 1, Fig. 1). In addition, the trials included two full-sib families from controlled crosses and two half-sib families from open pollination of south Finnish plus trees, as well as a progeny from open pollination of one Lithuanian plus tree.
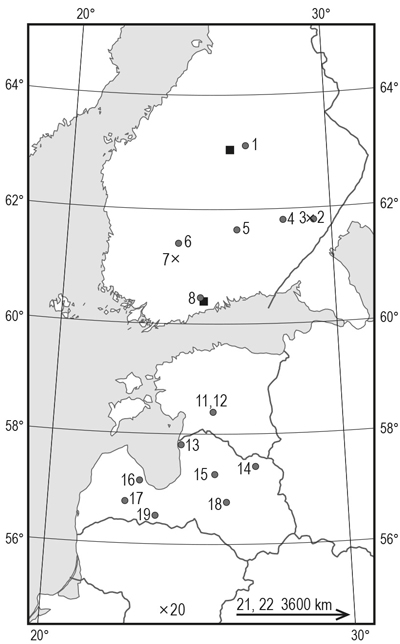
Fig. 1. Silver birch seed origins and location of the two field trials included in this study. ● = stand seed orgin, × = single tree origin, ■ = field trial. Numbering of origins refers to Table 1.
| Table 1. Silver birch seed origins included in this study. FIN = Finland, EST = Estonia, LVA = Latvia, LTU = Lithuania, RUS = Russia. Numbering of origins refers to Fig. 1. | ||||
| Origin | Latitude (N) | Longitude (E) | Altitude, m | |
| 1 | FIN, Pielavesi | 63°18´ | 26°49´ | 165 |
| 2 | FIN, Punkaharju | 61°48´ | 29°18´ | 92 |
| 3a | FIN, E1987 Punkaharju | 61°48´ | 29°18´ | 80 |
| 4 | FIN, Sulkava | 61°48´ | 28°10´ | 110 |
| 5 | FIN, Joutsa | 61°40´ | 26°15´ | 110 |
| 6 | FIN, Kangasala | 61°25´ | 24°09´ | 50 |
| 7a | FIN, E2812 Valkeakoski | 61°12´ | 24°00´ | 90 |
| 8 | FIN, Tuusula | 60°27´ | 24°58´ | 50 |
| 9a | FIN, E3012 Tuusula × E2378 Punkaharju | 60°22´ / 61°48´ | 25°03´ / 29°19´ | 60 / 80 |
| 10a | FIN, E3013 Tuusula × E2812 Valkeakoski | 60°22´ / 61°12´ | 25°03´ / 24°00´ | 60 / 90 |
| 11 | EST, Viljandi | 58°20´ | 25°30´ | |
| 12 | EST, Viljandi | 58°20´ | 25°30´ | |
| 13b | LVA, Salacgrīva | 57°49´ | 24°21´ | 10 |
| 14 | LVA, Alūksne | 57°28´ | 26°59´ | 180 |
| 15 | LVA, Liepa | 57°22´ | 25°30´ | 100 |
| 16 | LVA, Dursupe | 57°11´ | 22°58´ | 20 |
| 17 | LVA, Saldus | 56°42´ | 22°27´ | 120 |
| 18 | LVA, Jaunkalsnava | 56°41´ | 25°55´ | 100 |
| 19 | LVA, Zaļenicki | 56°31´ | 23°29´ | 30 |
| 20a | LTU, U8863 Girionys | 54°50´ | 24° | 72–100 |
| 21 | RUS, Novosibirskaya obl., Maslyanino | 54°30´ | 84° | |
| 22 | RUS, Novosibirskaya obl., Maslyanino | 54°30´ | 84° | |
| a single tree progeny b mixture of Betula pendula and Betula pubescens; excluded from the analysis | ||||
The trials were established on moist, upland forest sites, classified as a Myrtillus site type (MT) at Tuusula and as Oxalis-Myrtillus site type (OMT) at Viitasaari, according to the Finnish site type classification (Cajander 1949). Before planting, the sites were prepared by ploughing. The trials were established with two-year-old bare-rooted seedlings in May 1981. A randomised complete block design with four blocks of 22 plots each was used. Each plot consisted of 5 × 5 m (Viitasaari) and 7 × 7 m (Tuusula) trees of the same origin, in a spacing of 2 × 2 m.
2.2 Wood samples and measurements
The trials were measured when the trees were 22 years old. Tree height (dm) and diameter (mm) over bark at 1.3 m (dbh) were measured for all living trees.
For wood quality studies, samples were collected from all seed origins and blocks.
Two sample trees from each experimental plot were taken systematically (the second and the fourth tree of the middle row) for collection of wood samples, totalling eight sample trees per origin (4 blocks × 2 trees) in each trial. If the tree in the given location was missing, dead or clearly deviating from other trees in the plot regarding growth, then the nearest tree in the nearby row on either side was taken. Trees were felled and stem disks (5 cm in thickness) were taken at heights 1.3 m (breast height), 3 m and 6 m at both trials. In Tuusula, an additional disc was taken at stump height, but not at Viitasaari due to the high frequency of trees with basal sweep. Altogether, sample disks were taken from 168 and 151 trees from Tuusula and Viitasaari, respectively.
The diameter above and below bark was measured in two directions perpendicularly; bark thickness was calculated by subtraction and the average bark thickness and average diameters (above and below bark) for each sample disc were calculated. Incidence of darkened core wood, frost cracks, and decay was assessed as a 0or 1 trait (disc containing or not containing a defect). The sample disks were debarked and the basic density of wood (kg m–3) was determined according to the water displacement method by Olesen (1971). From here on, wood density refers to basic density, which denotes the dry-matter weight per unit volume of green weight.
2.3 Statistical methods and calculations
Statistical analyses were performed on the plot means, i.e. the means calculated from the two sample trees taken from each plot. Relative bark thickness (%) was calculated as the proportion of double bark thickness of the stem diameter over the bark value. Individual percentage values of relative bark thickness were arcsin √p transformed before analysis. The transformation had no effect on the test results and, thus, original values of the variables were used in the final analyses.
The pairwise linear dependences between tree height, dbh, wood density, bark thickness and relative bark thickness were examined with Pearson’s linear correlation coefficients calculated on the plot means.
Analysis of variance was used to test the significance of differences among the seed origins in tree height, diameters (above and below bark), wood density, bark thickness and relative bark thickness. The seed origins were divided into the Finnish and the Baltic (Estonia, Latvia, Lithuania) group, and the significance of differences in tree height, diameters (above and below bark), wood density, bark thickness and relative bark thickness between these two groups was tested using analysis of variance.
A linear mixed model analysis was used separately for the two trials to examine the effects of the latitude of seed origin on tree height and diameter (above bark) at breast height (dbh):
![]()
where y is the response variable, m is the general mean, Bi is the random effect of block i (i = 1,…,4), L is the fixed effect of seed origin latitude and ε is the random error term. Latitude (L) and the square of latitude (L2) were used as covariates.
A linear mixed model analysis was used separately for the two trials to examine the effects of latitude of seed origin and the tree height and diameter on wood density, bark thickness and relative bark thickness:
![]()
where y is the response variable, m is the general mean, Bi is the random effect of block i (i = 1,…,4) L is the fixed effect seed origin latitude, H is the average plot height (mean of the two sample trees), D is the average plot dbh (mean of the two sample trees) and ε is the error term. Latitude (L), the square of latitude (L2), average plot height (H) and average plot dbh (D) were used as covariates.
The linear mixed model analyses for testing the effect of seed origin latitude on height, dbh, wood density, bark thickness and relative bark thickness and the correlation analysis were performed only on the Finnish and Baltic seed origins. The two entries from Novosibirsk, Russia were excluded from the analyses because they are origins from a climatically different, continental area in Siberia (Table 1).
The coefficient of determination for seed origin latitude was computed as:

where ![]() is the residual variance of the model (1) or (2) and
is the residual variance of the model (1) or (2) and ![]() is the residual variance of the reduced model in which the seed origin latitude variables are not included.
is the residual variance of the reduced model in which the seed origin latitude variables are not included.
The proportion of trees (%) within each origin with darkened core wood, frost cracks and decay was calculated. Since eight trees/origin/trial were assessed, one tree was equivalent to 12.5%. Linear Pearson correlation coefficients were calculated between seed origin latitude, wood density (at 1.3 m), bark thickness (at 1.3 m), relative bark thickness (at 1.3 m) and the proportion of trees with darkened core wood, frost cracks and decay, excluding the Novosibirsk origins. Origin means of wood density, bark thickness and relative bark thickness were used in this correlation analyses. The significance of the differences in the proportions of trees with darkened core wood, frost cracks and decay between the Finnish and the Baltic group was tested using analysis of variance.
All analyses were performed using the statistical package SPSS 22.0 (SPSS, Chicago, IL).
3 Results
3.1 Height and diameter
Significant differences were detected in tree height among the seed origins in both trials, whereas the differences in dbh (above and below bark) were not significant (Table 2). The average height of the seed origins varied from 11.9 to 15.2 m at Viitasaari and from 12.4 to 16.7 m at Tuusula (for diameters see Table 2). At Viitasaari, best performing origins were found among the southern Finnish and Estonian ones, and at Tuusula among the Estonian and northern Latvian origins (Fig. 2). The origins from Lithuania and Novosibirsk performed poorly in both trials (Fig. 2). For example at Tuusula, the average height of the Novosibirsk origins was 13% lower than the trial mean and 27% lower than the mean height of the tallest origin. The effect of seed origin latitude on tree height was significant and curvilinear at both trials and on dbh at Tuusula (Fig. 2, Table 3). According to the model, the largest average height could be found in origins that were about 2 degrees of latitude south of the location of the trial. At Tuusula, trees of the Baltic group were taller than the trees of the Finnish group regarding height and dbh above bark, whereas at Viitasaari, the Finnish group was superior to the Baltic group regarding height, but the differences in breast height diameters were not significant (Table 2).
| Table 2. Average height and diameter at breast height (dbh) of the sample trees and their variation at the Viitasaari (63°11´N) and Tuusula (60°21´N) trials. The local origin at Viitasaari is Pielavesi (63°18´N) and that at the Tuusula trial is Tuusula (60°27´N). p-values indicate the significance of differences among origins and between the Finnish and Baltic groups of origins. Eight trees /origin /trial were sampled. N total = 168 (Tuusula) and 151 (Viitasaari). | ||||||||||||
| Trait | Trial | Average | SD | Min | Max | Lowest origin mean | Highest origin mean | Local origin mean | p origins | Baltic group mean | Finnish group mean | p groups |
| Height (m) | Viitasaari | 13.9 | 1.43 | 8.9 | 16.7 | 11.9 | 15.2 | 14.1 | <0.001 | 13.7 | 14.3 | 0.028 |
| Tuusula | 14.8 | 1.88 | 9.1 | 18.6 | 12.4 | 16.7 | 13.8 | <0.001 | 15.6 | 14.5 | <0.001 | |
| Dbh (cm) above bark | Viitasaari | 12.5 | 2.58 | 6.3 | 18.4 | 10.9 | 14.9 | 12.6 | ns | 12.7 | 12.7 | ns |
| Tuusula | 11.3 | 2.65 | 5.4 | 17.7 | 9.2 | 12.9 | 10.9 | ns | 11.9 | 11.0 | 0.044 | |
| Dbh (cm) below bark | Viitasaari | 11.6 | 2.38 | 5.8 | 17.2 | 9.8 | 13.5 | 11.8 | ns | 11.9 | 11.9 | ns |
| Tuusula | 10.6 | 2.53 | 5.1 | 17.1 | 8.8 | 12.2 | 10.2 | ns | 11.2 | 10.4 | ns | |
| SD = standard deviation. | ||||||||||||
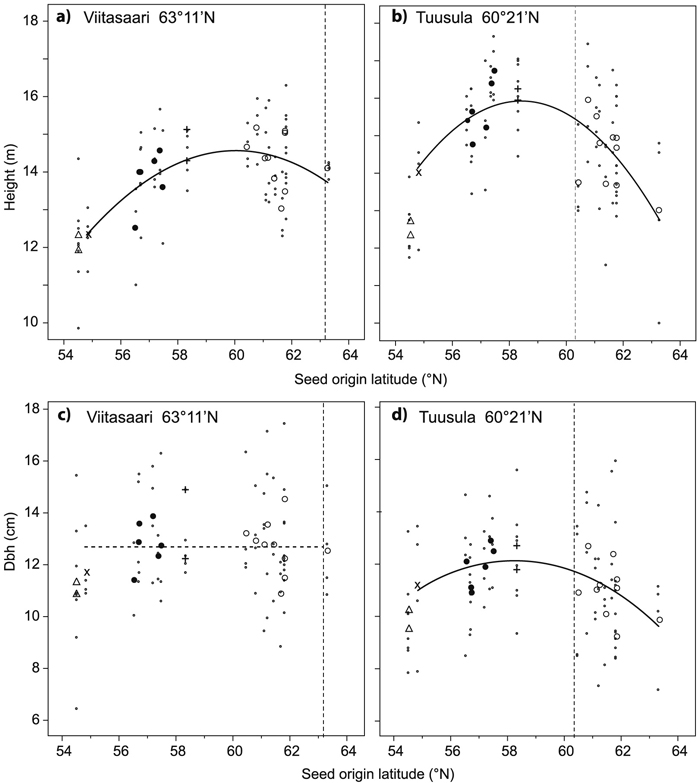
Fig. 2. Average height (ab) and diameter at breast height (dbh) (cd) of the silver birch origins in relation to seed origin latitude for the Viitasaari (ac) and Tuusula (bd) trials. The second order curves are based on the plot means, excluding Novosibirsk. ∙ = plot mean (mean of the two sample trees in a plot), bigger symbols are origin means (means of four plots) from different countries: ○ = Finland, + = Estonia, ● = Latvia, × = Lithuania, ∆ = Novosibirsk, Russia. A solid curve indicates a significant effect of latitude and a dashed curve a nonsignificant one. Latitude of the field trial is indicated by the vertical dashed line.
| Table 3. The parameter values of models (Eq. 1 and 2) for six response variables in the Viitasaari and Tuusula trials and p-values for the fixed factors included in the linear mixed models from which the parameter values of the models are determined. R2 is the coefficient of determination (%) for seed origin latitude (Eq. 3). | ||||||||||||
| Response variable | Trial | Intercept | Latitude | Latitude2 | Height | Dbh | Residual variance | R2 | ||||
| Coefficient | p | Coefficient | p | Coefficient | p | Coefficient | p | |||||
| Height a | Viitasaari | –278.25 | 9.75 | 0.002 | –0.081 | 0.002 | 0.964 | 21 | ||||
| Tuusula | –441.82 | 15.66 | <0.001 | –0.134 | <0.001 | 1.344 | 30 | |||||
| Dbh a | Viitasaari | 12.7 | 3.577 | - | ||||||||
| Tuusula | –318.62 | 11.36 | 0.048 | –0.098 | 0.045 | 3.725 | 8 | |||||
| Basic density 1.3 m b | Viitasaari | 366.45 | 7.618 | 0.001 | 364.950 | - | ||||||
| Tuusula | 692.22 | –3.55 | <0.001 | 309.190 | 19 | |||||||
| Basic density 3 m b | Viitasaari | 467.98 | 450.468 | - | ||||||||
| Tuusula | 3716.74 | –107.69 | 0.038 | 0.892 | 0.042 | 300.765 | 12 | |||||
| Bark thickness b | Viitasaari | 10.92 | –0.19 | <0.001 | 0.348 | <0.001 | 0.844 | 19 | ||||
| Tuusula | 11.34 | –0.19 | <0.001 | 0.306 | <0.001 | 0.749 | 21 | |||||
| Relative bark thickness b | Viitasaari | 21.69 | –0.26 | <0.001 | 1.736 | 17 | ||||||
| Tuusula | 23.48 | –0.29 | <0.001 | 1.767 | 21 | |||||||
| a Eq. 1 was used. b Eq. 2 was used. Dbh = diameter at breast height. | ||||||||||||
3.2 Wood density
Average wood density measured at 1.3 m height was 473 and 481 kg m–3 at the Viitasaari and Tuusula trials, respectively (Table 4, Fig. 3). Significant differences among the seed origins in wood density at 1.3 m height were detected at Tuusula. At Viitasaari, there was a moderate positive correlation between wood density at 1.3 m and tree height (r = 0.411, p = 0.001) and a weak positive correlation between wood density and dbh (r = 0.296, p = 0.015). Thus, at Viitasaari the tree height, but not the seed origin latitude, had a significant effect on wood density (Table 3). At Tuusula, no significant correlation between wood density and tree height or dbh was found, whereas the seed origin latitude had a significant effect on wood density at 1.3 m (Table 3). At Tuusula, wood density at the height of 1.3 m decreased linearly with increasing seed origin latitude (Fig. 3).
| Table 4. Average basic density (kg m–3) and its variation at different heights of the stem at the Viitasaari (63°11´N) and Tuusula (60°21´N) trials. The local origin at Viitasaari is Pielavesi (63°18´N) and that at the Tuusula trial is Tuusula (60°27´N). p-values indicate the significance of differences among origins and between the Finnish an Baltic groups of origins. Eight trees /origin /trial were sampled. N total = 168 (Tuusula) and 151 (Viitasaari). | ||||||||||||
| Measuring height | Trial | Average | SD | Min | Max | Lowest origin mean | Highest origin mean | Local origin mean | p origins | Baltic group mean | Finnish group mean | p groups |
| 0 m | Viitasaari | - | - | - | - | - | - | - | - | - | - | - |
| Tuusula | 511 | 26.6 | 428 | 576 | 487 | 531 | 494 | ns | 514 | 508 | ns | |
| 1.3 m | Viitasaari | 473 | 29.9 | 388 | 554 | 456 | 497 | 467 | ns | 468 | 478 | 0.044 |
| Tuusula | 481 | 25.7 | 407 | 546 | 458 | 510 | 458 | <0.001 | 489 | 475 | 0.001 | |
| 3 m | Viitasaari | 468 | 30.6 | 394 | 543 | 446 | 490 | 469 | ns | 464 | 471 | ns |
| Tuusula | 470 | 24.3 | 418 | 538 | 448 | 496 | 448 | <0.001 | 475 | 467 | ns | |
| 6 m | Viitasaari | 463 | 28.1 | 408 | 545 | 442 | 483 | 456 | ns | 459 | 466 | ns |
| Tuusula | 462 | 26.9 | 396 | 538 | 436 | 483 | 447 | <0.001 | 470 | 460 | 0.021 | |
| SD = standard deviation. | ||||||||||||
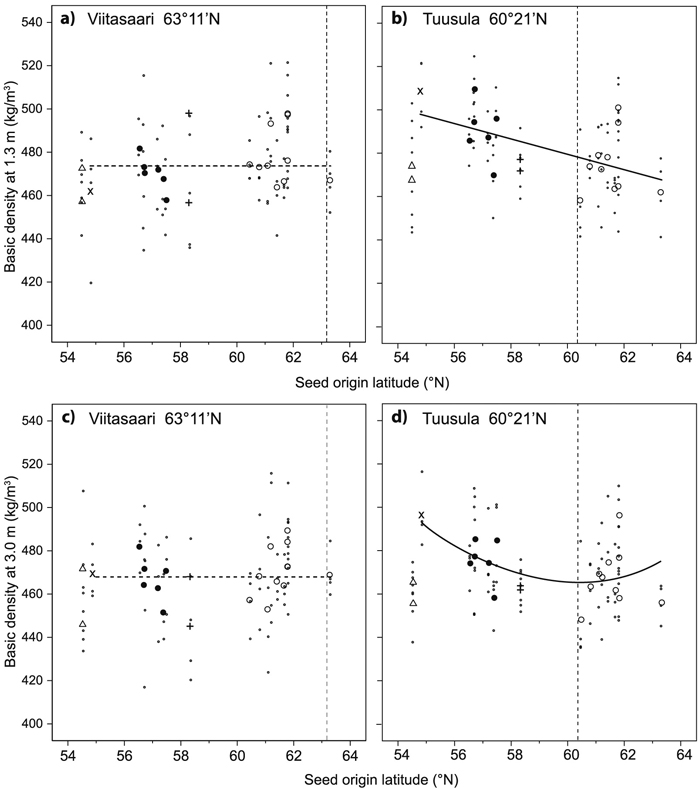
Fig. 3. Wood density at height 1.3 m (ab) and at height 3 m (cd) of silver birch origins in relation to seed origin latitude for the Viitasaari (ac) and Tuusula (bd) trials. The first order lines (abc) and second order curve (d) are based on the plot means, excluding Novosibirsk. ∙ = plot mean (mean of the two sample trees in the plot), bigger symbols are origin means (mean of four plots) from different countries: ○ = Finland, + = Estonia, ● = Latvia, × = Lithuania, ∆ = Novosibirsk, Russia. A solid line indicates a significant effect of latitude and a dashed line a nonsignificant one. Latitude of the field trial is indicated by the vertical dashed line.
In both trials the average wood density decreased systematically from the base of the stem upwards (Table 4). Significant differences among the seed origins in wood density at heights of 3 and 6 m were detected at Tuusula. No significant correlation was found between wood density at 3 m and tree height or dbh in either of the trials. At Viitasaari, neither the seed origin latitude nor the tree height had a significant effect on wood density at 3 m (Table 3). At Tuusula, the effect of seed origin latitude on wood density at 3 m was significant and curvilinear (Table 3, Fig. 3).
Wood density at the height of 1.3 m in the Baltic group was higher than that in the Finnish group at Tuusula (Table 4). At Viitasaari, the opposite was observed; the Finnish group had a significantly higher wood density at 1.3 m. No significant difference was detected between the two origin groups in wood density at 3 m in either of the trials.
3.3 Bark thickness
Average bark thickness measured at a height of 1.3 m was 4.2 and 3.6 mm at the Viitasaari and Tuusula trials, respectively (Table 5). Average bark thickness decreased systematically from the base of the stem upwards in both trials. Significant differences in bark thickness among the origins were detected at heights 0and 1.3 m at Tuusula (Table 5). At both trials, there was a moderate negative correlation between bark thickness and seed origin latitude (Viitasaari r = –0.368, p = 0.002, Tuusula r = –0.490, p < 0.001) and a moderate positive correlation with dbh (Viitasaari r = 0.537, Tuusula r = 0.596, both p < 0.001); at Tuusula there was also a weak positive correlation with tree height (r = 0.275, p = 0.016). In the linear mixed model analysis, both seed origin latitude and dbh had a significant effect on bark thickness in both trials (Table 3, Fig. 4). Bark thickness decreased with increasing seed origin latitude and increased with increasing tree dbh. At Tuusula, the Baltic group had thicker bark compared to the Finnish group at all heights and at Viitasaari at a height of 1.3 m. (Table 5).
| Table 5. Average bark thickness (mm) and its variation at different heights of the stem at the Viitasaari (63°11´N) and Tuusula (60°21´N) trials. The local origin at Viitasaari is Pielavesi (63°18´N) and that at Tuusula trial is Tuusula (60°27´N). p-values indicate the significance of differences among origins and between the Finnish and Baltic groups of origins. Eight trees /origin /trial were sampled. N total = 168 (Tuusula) and 151 (Viitasaari). | ||||||||||||
| Measuring height | Trial | Average | SD | Min | Max | Lowest origin mean | Highest origin mean | Local origin mean | p origins | Baltic group mean | Finnish group mean | p groups |
| 0 m | Viitasaari | - | - | - | - | - | - | - | - | - | - | |
| Tuusula | 11.4 | 4.51 | 2.5 | 24.3 | 6.7 | 15.5 | 9.8 | 0.023 | 12.6 | 10.0 | 0.001 | |
| 1.3 m | Viitasaari | 4.2 | 1.64 | 0.2 | 12.5 | 3.3 | 7.1 | 3.8 | ns | 4.6 | 3.8 | 0.005 |
| Tuusula | 3.6 | 1.64 | 1.0 | 11.3 | 2.2 | 5.4 | 3.7 | 0.003 | 4.1 | 3.1 | <0.001 | |
| 3 m | Viitasaari | 2.9 | 0.96 | 1.3 | 6.7 | 2.4 | 3.8 | 2.4 | ns | 3.1 | 2.9 | ns |
| Tuusula | 2.7 | 0.86 | 1.0 | 5.5 | 2.2 | 3.3 | 2.3 | ns | 3.0 | 2.5 | <0.001 | |
| 6 m | Viitasaari | 2.4 | 0.85 | 1.0 | 6.3 | 2.0 | 3.5 | 2.2 | ns | 2.6 | 2.3 | ns |
| Tuusula | 2.2 | 0.82 | 0.0 | 4.7 | 1.6 | 2.9 | 1.7 | ns | 2.4 | 2.1 | 0.033 | |
| SD = standard deviation. | ||||||||||||
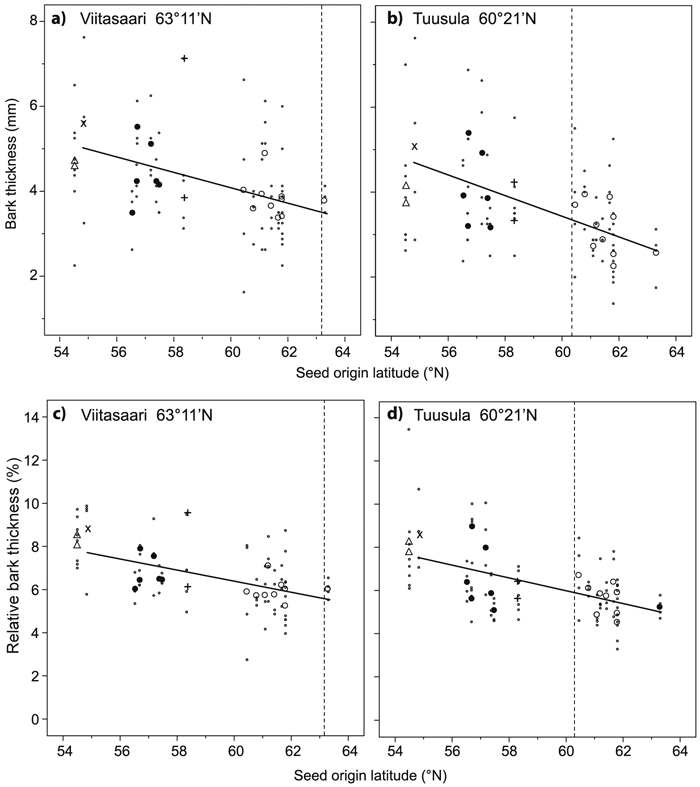
Fig. 4. Bark thickness (ab) and relative proportion of the bark from diameter over bark (cd) in relation to seed origin latitude for the Viitasaari (ac) and Tuusula (bd) trials. The first order lines are based on the plot means, excluding Novosibirsk. ∙ = plot mean (mean of the two sample trees in the plot), bigger symbols are origin means (mean of four plots) from different countries: ○ = Finland, + = Estonia, ● = Latvia, × = Lithuania, ∆ = Novosibirsk, Russia. A solid line indicates a significant effect of latitude. Latitude of the field trial is indicated by the vertical dashed line.
3.4 Relative bark thickness
Average relative bark thickness, which denotes the percentage of the doubled bark thickness from the dbh above bark, was 6.7 and 6.3% at the Viitasaari and Tuusula trials, respectively. Variation among the origins in relative bark thickness at a height of 1.3 m was significant at both trials (both p < 0.001). Similar patterns of variation in relative bark thickness were detected as for absolute bark thickness. Average relative bark thickness decreased from the base of the stem upwards (not presented). In the linear mixed model analysis, the latitude of seed origin had a significant effect on the relative thickness of the bark in both trials (Table 3, Fig. 4). Relative bark thickness decreased with increasing seed origin latitude. The relative bark thickness at a height of 1.3 m of the Baltic group was higher than that of the Finnish group in both trials (both p < 0.001) and at Tuusula, relative bark thickness of the Baltic group was also higher a height of 3 m (p = 0.034).
3.5 Incidence of darkened core wood, frost cracks and decay
Darkened core wood was detected in 54 and 81% of the trees at Viitasaari and Tuusula, respectively (Table 6). At Tuusula, a moderate positive correlation (r = 0.517, p = 0.023) was detected between the proportion of trees with darkened core wood and the seed origin latitude in the Finnish and Baltic origins. However, also in the Novosibirsk origin, darkened core wood was detected in all sample trees. A moderate negative correlation was also detected between darkened core wood and bark thickness (r = –0.529, p = 0.020) and relative bark thickness (r = –0.506, p = 0.027) at Tuusula. The proportion of trees with darkened core wood was significantly (p = 0.033) higher in the Finnish (85%) group of origins than in the Baltic (74%) group in Tuusula.
| Table 6. Proportion of trees (%) with darkened core wood, frost crack or decay and their variation at the Viitasaari (63°11´N) and Tuusula (60°21´N) trials. The local origin at Viitasaari is Pielavesi (63°18´N) and that at the Tuusula trial is Tuusula (60°27´N). Eight trees /origin /trial were sampled. Here one sample tree is equivalent to 12.5%. N total = 168 (Tuusula) and 151 (Viitasaari). | ||||||
| Trait | Trial | Average | SD | Lowest origin | Highest origin | Local origin |
| Black pith | Viitasaari | 54 | 30 | 0 | 100 | 88 |
| Tuusula | 81 | 12 | 63 | 100 | 75 | |
| Frost crack | Viitasaari | 15 | 14 | 0 | 50 | 13 |
| Tuusula | 5 | 8 | 0 | 25 | 0 | |
| Decay | Viitasaari | 6 | 8 | 0 | 25 | 0 |
| Tuusula | 5 | 6 | 0 | 13 | 0 | |
| SD = standard deviation. | ||||||
The proportion of trees with frost cracks was 15 and 5% at Viitasaari and Tuusula, respectively (Table 6). At Viitasaari, there was a tendency of the southern origins to have more trees with frost cracks, but the correlation was not significant (r = –0.408, p = 0.083). Highest proportion of trees with frost cracks was detected in the south-western Latvian origins Zalenicki (50%) and Dursupe (38%).
Decay was detected in 6 and 5% of the sample trees at Viitasaari and Tuusula, respectively, but no significant correlation with the seed origin latitude was found (Table 6). At Viitasaari, the highest proportion of trees with decay was detected in the south-western Latvian origins Dursupe (25%) and Saldus (17%). A moderate positive correlation (r = 0.502, p = 0.023) was detected between the incidence of frost cracks and decay at Tuusula.
4 Discussion
Variations in growth, wood density, bark thickness, darkened core wood, frost cracks and decay of 22-year-old trees were studied among north European silver birch origins, and non-native Baltic origins were compared with the native Finnish ones in two Finnish provenance trials. The results of this study can provide some cues for the feasibility and use of non-native southern seed origins regarding these biomass properties. So far, this is the first study on pulp-sized trees representing different provenances from a wide range of latitudes, since earlier studies on genetic variation of wood quality are based on younger trees or have been carried out on either progeny or clonal level within a more restricted geographical area (Velling 1979a; Nepveu and Velling 1983; Tammisola et al. 1995; Stener and Hedenberg 2003).
The average tree height in both trials and the average dbh at Tuusula varied curvilinearly in relation to the latitude of seed origin, which is well in accordance with the pattern of variation in growth reported earlier by e.g. Eriksson and Johnsson (1986), Viherä-Aarnio and Velling (2008) and Viherä-Aarnio et al. (2013). According to the models, the highest mean height and dbh values would be attained by origins that are transferred ca. 2 degrees of latitude from the south, whereas long transfers either from the north or from the south would result in weaker growth. On average, the growth of the Baltic group was superior to that of the Finnish origins at Tuusula, southern Finland, but no more at Viitasaari, where they had been transferred as long as 5–7 degrees of latitude to the north. The reasons for the different performance of the seed origins and the effects of seed transfer on their performance lay in their different climatic adaptation and their response to different photoperiodic conditions (Håbjørg 1972, 1978; Viherä-Aarnio et al. 2006) and, consequently, differences in the length of their growth periods (Velling 1979b; Viherä-Aarnio et al. 2005).
The basic density values observed in the 22-year-old trees in our study (average 468 and 470 kg m–3 at 3 m height at Viitasaari and Tuusula, respectively) are well in line with e.g. the results of Hakkila (1979) and Tammisola et al. (1995). The basic density of birches younger than 25 years was over 450 kg m–3 (Hakkila 1979). The basic density of a 17-year-old southern Finnish silver birch progeny was 450 kg m–3 while that of 28-year-old silver birch progenies varied between 468 and 500 kg m–3 (Tammisola et al.1995). Tree age has an effect on wood density values, i.e. birch produces denser wood as the tree ages (Hakkila 1979; Bhat 1980). In our study, the average basic density decreased systematically from the stump height upwards, which is also in accordance with earlier studies (Walden 1934; Jalava 1945; Kujala 1946; Hakkila 1966; Velling 1979a).
The basic density of birch trees decreased in relation to the seed origin latitude at the Tuusula trial, but no such effect was found at Viitasaari. At Tuusula, the Baltic group had a higher basic density at 1.3 m than the Finnish group, whereas at Viitasaari, the opposite was observed. Further, in the basic density at 3 m, there was no significant difference between the two origin groups in either of the trials. This result at 3 m probably well represents the average wood density of the sample tree stems, because the fixed height of 3 m falls within the range of 25% relative height (2.2–4.7 m) in our sample trees, which, according to Hakkila, (1979) best represents the average wood density of the stem. Björklund (1984) reported decreasing basic density from south to north in Swedish birch pulpwood samples harvested from the original growing sites of the trees. In the study of Velling (1979a), progeny transfers of less than 250 km had no effect on wood density. In the end, the average effect of seed origin and seed transfer on basic density seems to be quite small and vague and may, after all, be partly hampered by the effect of tree height due to reaction wood formation (see discussion below).
The significant positive effect of tree height, which in our even-aged material indicates the growth rate, on wood density at 1.3 m in the Viitasaari trial is inconsistent with results from some of the earlier studies (Table 3, Velling 1979a; Nepveu and Velling 1983). No dependence was found between the growth characteristics of the stem, i.e. height, diameter at breast height and volume and wood density of silver birch by Velling (1979a) and Nepveu and Velling (1983). According to Tsoumis (1991), growth rate should have little direct influence on timber density of diffuse porous species such as birch, and a similar conclusion was presented by Dunham et al. (1999) on silver birch. However, Stener and Hedenberg (2003) found a significant intermediate negative genotypic correlation between wood density and diameter, which is contradictory to our result. The significant positive effect of tree height on basic density at 1.3 m at Viitasaari might be due to reaction wood, which is heavier than average normal wood (Ollinmaa 1956). The higher the trees, the stronger the influence of wind on the basal part of the stem, which in turn could explain the positive correlation between tree height and basic density at 1.3 m. Although not measured in this study, the reaction wood could be caused by basal sweep, which was common at the Viitasaari trial as a result of erroneous planting at the edge of the furrow in this ploughed experimental area. Basal sweep and crooks due to the leaning of the stem are commonly found in planted birches (Niemistö et al. 1997), and adequate planting techniques is stressed by Verkasalo and Paukkonen (1999). No significant correlation was found between tree size and wood density at 3 m at Viitasaari, and no significant correlation between tree size and wood density was observed at Tuusula. To conclude, neither seed origin nor tree height seems to have any clear and consistent effect on wood density.
Bark thickness as well as relative bark thickness decreased with increasing seed origin latitude. Although bark thickness increased significantly with increasing stem diameter, i.e. thicker trees have thicker bark, the effect of latitude was still highly significant (Table 3, Fig. 4). Our result is well in accordance with the observations of Lindquist (1947), who thoroughly elucidated the question of the regional variation of silver birch bark. Lindquist (1947) divided the European populations of silver birch into two varieties, i.e. var. saxatilis and var. lapponica, the former having its main distribution area in Central Europe, central and southern Russia, southern Sweden and southernmost Finland, whereas the latter grows in more northern parts of Fennoscandia. A markedly fissured, thick or stony bark in the lower part of the trunk is characteristic for the southern variety. The increasing bark thickness when moving towards south seems to hold true, although the division of the Finnish silver birch populations into varieties in the present taxonomy (Hämet-Ahti et al. 1992) differs from that of Lindquist (1947). Thus, the bark thickness of birch logs in Finland would be increased by using non-native Baltic seed origins transferred from the south.
Trees with darkened core wood were found in both trials, and at Tuusula, their incidence increased with increasing seed origin latitude. Stem discolorations around the pith were commonly found in planted birches with an age of 18–65 years in the study of Hallaksela and Niemistö (1998). Studies on their relation to the genetic background of the trees are, however, few. Bitenieks et al. (2012) discovered discoloration of wood already in 10-year-old silver birch trees, and variation in discoloration was found between families, indicating a possible genetic component. In a southern Finnish progeny test, discoloration was observed in all studied full-sib families of silver birch, and significant variation occurred also between individual trees (Tammisola et al. 1995). In the study of Tammisola et al. (1995), stem discoloration appeared to increase with the increasing moisture content of the site. In our study, trees with stem discoloration were found in both trials, but their proportion was somewhat higher at Tuusula, which may thus be related to the site, since the site at Tuusula was sporadically moister than at Viitasaari. According to Hallaksela and Niemistö (1998), stem discolorations are slightly more common on former agricultural land than on forest sites. The causal micro-organism and mechanism of discoloration have not been fully elucidated. Presumably, the invasion of microbes takes place via broken branches in connection with natural or artificial pruning (Hallaksela and Niemistö 1998; Bitenieks et al. 2012). Wood discoloration can also be caused by vole damages during the first years of the rotation.
The highest number of trees with frost cracks was found among the south-western Latvian origins. Frost cracks occur when the surface of the stem freezes and, consequently, shrinks more rapidly than the inner parts. According to Niemistö et al. (1997), low spacing of birch stands increased the number of frost cracks. The low survival of the southernmost seed origins (Viherä-Aarnio and Velling 2008) and, consequently, their low spacing may be an explanation for the higher number of frost cracks in these origins. In Norway spruce, frost cracks were more common in trees facing gaps and forest edges as well as in fast-growing trees of late hardening provenances (Persson 1994), which is also in line with our results. In our study, the number of trees with decay was low, as expected, since the trees were only 22 years old, but the incidence of decay was connected to the incidence of frost cracks.
Eight sample trees were taken per origin in each trial, which is a low number and may not represent the entire variation within a provenance. However, the trees sampled for this study showed a similar pattern of variation in height and diameter as detected in an earlier study including the entire, wider material of the same provenance trials (Viherä-Aarnio and Velling 2008). This indicates that the wood quality sample trees selected for this study well represented the entire material. Wood samples for this study were taken at fixed heights, i.e. at 0, 1.3, 3.0 and 6.0 meters instead of relative heights, which may possibly skew the comparisons in case the sample trees differ significantly in total height. However, for example, the fixed height of 3 m falls within the range of 25% relative height (2.2–4.7 m) in our sample trees, which according to Hakkila (1979) best represents the average wood density of the stem.
The effect of latitude of seed origin on the studied traits was small, since all the coefficients of determination for seed origin latitude were low. A clear and systematic effect of seed origin latitude was seen on bark thickness, whereas the effect on wood density was not consistent and would probably have no or minor practical significance. Long transfer of southern Baltic origins to Finland would probably increase the risk of frost cracks.
Although Baltic silver birch origins can have slightly higher yields compared to our native stand seed origins, the use of non-native Baltic seed origins in Finland would not provide definite advantage compared to our native locally adapted and genetically improved seed sources. The results of this study, and the findings of earlier studies regarding external stem quality, indicate that there is no reason to recommend the use of Baltic origins in Finland.
Acknowledgements
Erkki Kosonen, Veijo Hakamäki, Juhani Mäkinen and Heimo Tynkkynen measured the trees in the field and collected the sample discs. Jukka Lehtonen and Maija Pekkonen carried out wood sample analyses. Anne Siika and Sari Elomaa drew the final figures. Risto Häkkinen provided valuable advice in the statistical analysis. We wish to thank them all for their help and co-operation. Jyrki Hytönen is thanked for his valuable comments on the text. The Finnish Forest and Park Service is acknowledged for co-operation in establishment and early management of the trial at Viitasaari. We also wish to thank the two anonymous referees for their constructive comments on the manuscript. This study was financed by The Finnish Forest Research Institute and the Finnish Academy as a part of the Life2000 – Research programme for biological functions.
References
Ahtikoski A. (2000). The profitability of Scots pine (Pinus sylvestris L.) and silver birch (Betula pendula Roth) next-generation seed orchards in Finland. Academic dissertation. University of Helsinki. 148 p. + 41 app. http://urn.fi/URN:ISBN:951-91-2327-2.
Alakurtti S., Mäkelä T., Koskimies S.,Yli-Kauhaluoma J. (2006). Pharmacological properties of the ubiquitous natural product betulin. European Journal of Pharmaceutical Sciences 29(1): 1–13. https://doi.org/10.1016/j.ejps.2006.04.006.
Bhat K.M. (1980). Variation in structure and selected properties of Finnish birch wood: I. Interrelationships of some structural features, basic density and shrinkage. Silva Fennica 14(4): 384–396. https://doi.org/10.14214/sf.a15032.
Bhat K.M. (1983). A review of wood anatomy and selected properties of stems, branches and roots of birch species. IAWA Bulletin 4(2–3): 70.
Bhat K.M., Kärkkäinen M. (1981). Variation in structure and selected properties of Finnish birch wood: III. Proportion of wood elements in stems and branches in Betula pendula Roth. Silva Fennica 15(1): 1–9. https://doi.org/10.14214/sf.a15036.
Bitenieks K., Korica A., Bacāns Ž., Nikolajeva V., Baumanis I., Veinberga I., Ruņģis D. (2012). Identification of microorganisms found in discoloured birch (Betula pendula L.) wood. Mežzinātne 25(58): 180.
Björklund L. (1984). Massavedens torr-rådensitet och dess beroende av olika faktorer. Summary: Pulpwood basic density and it’s dependence on different factors. The Swedish University of Agricultural Sciences, Department of Forest Products. Report 155: 1–29.
Cajander A.K. (1949). Forest types and their significance. Acta Forestalia Fennica 56. 71 p. https://doi.org/10.14214/aff.7396.
Dunham R.A., Cameron A.D., Petty J.A. (1999). The effect of growth rate on the strength properties of sawn beams of silver birch (Betula pendula Roth). Scandinavian Journal of Forest Research 14(1): 18–26. https://doi.org/10.1080/02827589908540805.
Eriksson G., Jonsson A. (1986). A review of the genetics of Betula. Scandinavian Journal of Forest Research 1(1–4): 421–434. https://doi.org/10.1080/02827588609382434.
Håbjørg A. (1972). Effects of photoperiod and temperature on growth and development on three latitudinal and three altitudinal populations of Betula pubescens Ehrh. Meldinger fra Norges Landbrukshøgskole 51(2). 27 p.
Håbjørg A. (1978). Photoperiodic ecotypes in Scandinavian trees and shrubs. Meldinger fra Norges Landbrukshøgskole 57(33). 20 p.
Hagqvist R. (1991). Jalostetun koivunsiemenen tuotanto ja saatavuus. Summary: Production of genetically improved birch seed and micropropagated seedlings. Foundation for Forest Tree Breeding in Finland. Year Book 1991. p. 12–17, 31.
Hakkila P. (1966). Investigations on the basic density of Finnish pine, spruce, and birch wood. Communicationes Instituti Forestalis Fenniae 61(5). 98 p. http://urn.fi/URN:NBN:fi-metla-201207171093.
Hakkila P. (1979). Wood density survey and dry weight tables for pine, spruce and birch stems in Finland. Communicationes Instituti Forestalis Fenniae 96(3). 59 p. http://urn.fi/URN:ISBN:951-40-0470-1.
Hallaksela A.-M., Niemistö P. (1998). Stem discoloration of planted silver birch. Scandinavian Journal of Forest Research 13(1–4): 169–176. https://doi.org/10.1080/02827589809382973.
Hämet-Ahti L., Palmén A., Alanko P., Tigerstedt P.M.A. (1992). Suomen puu- ja pensaskasvio. [Woody flora of Finland]. Publications of the Finnish Dendrological Society 6. 373 p. Yliopistopaino, Helsinki. ISBN 951-96557-0-0.
Heiskanen V. (1966). Tutkimuksia koivujen vikaisuuksista, niiden vaikutuksesta sorvaustulokseen sekä niiden huomioonottamisesta laatuluokituksessa. Summary: Studies on the defects of birch, their influence on the quality and quantity of rotary cut veneer, and their consideration in veneer birch grading. Acta Forestalia Fennica 80(3). 128 p. https://doi.org/10.14214/aff.7165.
Heräjärvi H. (2002). Properties of birch (Betula pendula, B. pubescens) for sawmilling and further processing in Finland. Academic dissertation, University of Joensuu. Finnish Forest Research Institute, Research Papers 871. 52 p. + 5 original papers. http://urn.fi/URN:ISBN:951-40-1856-7.
Heräjärvi H. (2004a). Static bending properties of Finnish birch wood. Wood Science and Technology 37(6): 523–530. https://doi.org/10.1007/s00226-003-0209-1.
Heräjärvi H. (2004b). Variation of basic density and Brinell hardness within mature Finnish Betula pendula and B. pubescens stems. Wood and Fiber Science 36(2): 216–227.
Hultén E., Fries M. (1986). Atlas of North European vascular plants: north of the Trophic of Cancer. I. Introduction. Taxonomic Index to Maps 1–996. Maps 1–996. Koeltz Scientific Books, Königstein, Germany. 498 p.
Hynynen J., Niemistö P., Viherä-Aarnio A., Brunner A., Hein S., Velling P. (2010). Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83(1): 103–119. https://doi.org/10.1093/forestry/cpp035.
Jalava M. (1945). Suomalaisen männyn, kuusen, koivun ja haavan lujuusominaisuuksista. Summary: Strength properties of Finnish pine, spruce, birch and aspen. Communicationes Instituti Forestalis Fenniae 33(3). 66 p. http://urn.fi/URN:NBN:fi-metla-201207171065.
Jonsell B. (ed.). (2000). Flora Nordica. Lycopodiaceae to Polygonaceae. Vol. 1. The Bergius Foundation, The Royal Swedish Academy of Sciences, Stockholm, Sweden. 344 p.
Kärkkäinen M. (1986). Koivutukkien ja -runkojen arvosuhteet. Abstract: Value relations of birch veneer logs and stems. Silva Fennica 20(1): 45–57. https://doi.org/10.14214/sf.a15440.
Krasutsky P.A. (2006). Birch bark research and development. Natural Product Reports 23(6): 919–942. https://doi.org/10.1039/b606816b.
Kujala V. (1946). Koivututkimuksia. Summary: Some recent data on birches. Communicationes Instituti Forestalis Fenniae 34(1). 36 p. http://urn.fi/URN:NBN:fi-metla-201207171066.
Lindquist B. (1947). On the variation in Scandinavian Betula verrucosa Ehrh. with some notes on the Betula series Verrucosae Sukacz. Svensk Botanisk Tidskrift 41: 45–80.
Luostarinen K., Verkasalo E. (2000). Birch as sawn timber and mechanical further processing in Finland. Silva Fennica Monographs 1. 40 p. https://www.silvafennica.fi/pdf/smf001.pdf.
Marttila V., Granholm H., Laanikari J., Yrjölä T., Aalto A., Heikinheimo P., Honkatukia J., Järvinen H., Liski J., Merivirta R., Paunio M. (2005). Ilmastonmuutoksen kansallinen sopeutumisstrategia. Abstract: Finland’s national strategy for adaptation to climate change. Publications of the Ministry of Agriculture and Forestry 1/2005. 276 p.
Nepveu G., Velling P. (1983). Rauduskoivun puuaineen laadun geneettinen vaihtelu. Summary: Individual genetic variability of wood quality in Betula pendula. Folia Forestalia 575. 21 p. http://urn.fi/URN:ISBN:951-40-0643-7.
Niemistö P., Hukki P., Verkasalo E. (1997). Kasvupaikan ja puuston tiheyden vaikutus rauduskoivun ulkoiseen laatuun 30-vuotiaissa istutuskoivikoissa. [Effect of site and spacing on stem quality of silver birch in 30-year-old planted birch stands]. Metsätieteen Aikakauskirja 3/1997: 349–374. https://doi.org/10.14214/ma.6476.
Olesen P.O. (1971). The water displacement method. Forest Tree Improvement 3: 3–23.
Ollinmaa P.J. (1956). Koivun vetopuun anatomisesta rakenteesta ja ominaisuuksista. Summary: On the anatomic structure and properties of tension wood in birch. Acta Forestalia Fennica 64(3). 263 p. https://doi.org/10.14214/aff.7463.
Persson A. (1994). Stem cracks in Norway spruce in southern Scandinavia: causes and consequences. Annales des Sciences Forestières 51(3): 315–327. https://doi.org/10.1051/forest:19940310.
Repola J. (2006). Models for vertical wood density of Scots pine, Norway spruce and birch stems, and their application to determine average wood density. Silva Fennica 40(4): 673–685. https://doi.org/10.14214/sf.322.
Raulo J. (1978). Forestation chain for birch (Betula pendula Roth) in Finland. Silva Fennica 12(1): 17–24. https://doi.org/10.14214/sf.a14838.
Stener L.-G., Hedenberg Ö. (2003). Genetic parameters of wood, fibre, stem quality and growth traits in a clone test with Betula pendula. Scandinavian Journal of Forest Research 18(2): 103–110. https://doi.org/10.1080/02827580310003678.
Tammisola J., Varhimo A., Velling P., Viherä-Aarnio A., Kauppinen V., Sundquist J., Lapinjoki S. (1995). Variation in the wood and pulping properties of European white birch. Paperi ja Puu – Paper and Timber 77(10): 648–654.
Tsoumis G. (1991). Science and technology of wood. Structure, properties and utilization. Van Norstand Reinhold, New York. 494 p. ISBN: 978-3-941300-22-4.
Velling P. (1979a). Puuaineen tiheys kahdessa rauduskoivun jälkeläiskokeessa. Summary: Wood density in two Betula pendula Roth progeny trials. Folia Forestalia 416. 24 p. http://urn.fi/URN:ISBN:951-40-0425-6.
Velling P. (1979b). Erilaisten rauduskoivuprovenienssien alkukehityksestä taimitarhalla ja kentäkokeessa. Summary: Initial development of different Betula pendula Roth provenances in the seedling nursery and in field trials. Folia Forestalia 379. 14 p. http://urn.fi/URN:ISBN:951-40-0373-x.
Verkasalo E., Paukkonen K. (1999). Koivun ominaisuudet ja käyttömahdollisuudet sahauksessa ja jatkojalostuksessa Suomessa. Summary: Properties and potential of birch (Betula sp.) for utilisation in saw milling and further processing in Finland. Finnish Forest Research Institute, Research Paper 751. 91 p. http://urn.fi/URN:ISBN:951-40-1706-4.
Verkasalo E., Heräjärvi H., Möttönen V., Haapala A., Brännström H., Vanhanen H., Miina J. (2017). Current and future products as the basis for value chains of birch in Finland. Proceedings of the International Scientific Conference on Hardwood Processing 2017, Lahti, Finland. In press.
Viherä-Aarnio A., Heikkilä R. (2006). Effect of the latitude of seed origin on moose (Alces alces) browsing on silver birch (Betula pendula). Forest Ecology and Management 229(1–3): 325–332. https://doi.org/10.1016/j.foreco.2006.04.011.
Viherä-Aarnio A., Velling P. (2008). Seed transfers of silver birch (Betula pendula) from the Baltic to Finland – effect on growth and stem quality. Silva Fennica 42(5): 735–751. https://doi.org/10.14214/sf.226.
Viherä-Aarnio A., Häkkinen R., Partanen J., Luomajoki A., Koski V. (2005). Effects of seed origin and sowing time on timing on height growth cessation of Betula pendula seedlings. Tree Physiology 25(1): 101–108. https://doi.org/10.1093/treephys/25.1.101.
Viherä-Aarnio A., Häkkinen R., Junttila O. (2006). Critical night length and its variation in two photoperiodic ecotypes of Betula pendula. Tree Physiology 26(8): 1013–1018. https://doi.org/10.1093/treephys/26.8.1013.
Viherä-Aarnio A., Kostiainen K., Piispanen R., Saranpää P., Vapaavuori E. (2013). Effects of seed transfers on yield and stem defects of silver birch (Betula pendula Roth). Forest Ecology and Management 289: 133–142. https://doi.org/10.1016/j.foreco.2012.10.030.
Walldén P. (1934). Tutkimuksia koivupuun anatoomisen rakenteen ja teknillisten ominaisuuksien keskinäisestä riippuvaisuudesta solumittauksien perusteella. Referat: Untersuchungen über die Abhängigkeit der technischen Eigenschaften vom anatomischen Bau des Birkenholzes nach Zellenmessungen. Acta Forestalia Fennica 40(14): 329–366. https://doi.org/10.14214/aff.7291.
Total of 51 references.
