Jyrki Hytönen  ,
Egbert Beuker,
Anneli Viherä-Aarnio
,
Egbert Beuker,
Anneli Viherä-Aarnio
Clonal variation in basic density, moisture content and heating value of wood, bark and branches in hybrid aspen
Hytönen J., Beuker E., Viherä-Aarnio A. (2018). Clonal variation in basic density, moisture content and heating value of wood, bark and branches in hybrid aspen. Silva Fennica vol. 52 no. 2 article id 9938. https://doi.org/10.14214/sf.9938
Highlights
- Hybrid aspen clones differed in their moisture content, ash content, basic density and heating value
- Stem wood had lower ash content, basic density and effective heating value than stem bark
- There was significant vertical variation in wood and bark along the stem in moisture content and basic density.
Abstract
Hybrid aspen (Populus tremula × P. tremuloides) is one of the fastest growing tree species in Finland. During the mid-1990s, a breeding programme was started with the aim of selecting clones that were superior in producing pulpwood. Hybrid aspen can also be grown as a short-rotation crop for bioenergy. To study clonal variation in wood and bark properties, seven clones were selected from a 12-year-old field trial located in southern Finland. From each clone, five trees were harvested and samples were taken from stem wood, stem bark and branches to determine basic density, effective heating value, moisture and ash content. Vertical within-tree variation in moisture content and basic density was also studied. The differences between clones were significant for almost all studied properties. For all studied properties there was a significant difference between wood and bark. Wood had lower ash content (0.5% vs. 3.9%), basic density (378 kg m–3 vs. 450 kg m–3) and effective heating value (18.26 MJ kg–1 vs. 19.24 MJ kg–1), but higher moisture content (55% vs. 49%) than bark. The values for branches were intermediate. These results suggest that the properties of hybrid aspen important for energy use could be improved by clonal selection. However, selecting clones based on fast growth only may be challenging since it may lead to a decrease in hybrid aspen wood density.
Keywords
moisture content;
wood;
bark;
hybrid aspen;
basic density;
branches;
clonal variation;
heating value;
ash content
-
Hytönen,
Natural Resources Institute Finland (Luke), Natural resources, Teknologiakatu 7, FI-67100 Kokkola, Finland
E-mail
jyrki.hytonen@luke.fi
- Beuker, Natural Resources Institute Finland (Luke), Production systems, Vipusenkuja 6, FI-57200 Savonlinna, Finland E-mail egbert.beuker@luke.fi
- Viherä-Aarnio, Natural Resources Institute Finland (Luke), Production systems, Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail anneli.vihera-aarnio@luke.fi
Received 13 December 2017 Accepted 5 April 2018 Published 10 April 2018
Views 101095
Available at https://doi.org/10.14214/sf.9938 | Download PDF

1 Introduction
Hybrid aspen (Populus × wettsteinii Hämet-Ahti), a cross between the native European aspen (P. tremula L.) and American aspen (P. tremuloides Michx.), is considered to be the fastest growing tree species in Finland (Hynynen 1999; Beuker et al. 2016). In the 1950s and 60s, hybrid aspen was cultivated in Finland with the purpose of growing raw material for the match industry, but with the fall of match industry in the early 1970s interest in aspen decreased (Beuker 2000; Kärki 2001). A new breeding programme with hybrid aspen was started in Finland during the mid-1990s with the aim of producing pulpwood (Beuker 2000). Individual hybrid aspen trees, superior in growth and quality, were selected phenotypically from stands and trials that had been planted during the 1950s and 60s. The selected trees were vegetatively propagated through micro-propagation. Because of the large variation between the trees in succeeding in the propagation process, only clones that had a high rate of propagation success were selected for further testing in field trials.
For pulpwood production, the recommendation is to plant about 1200 hybrid aspen plants per hectare and grow for 20 to 25 years without thinning. After clear cutting, the next generation can be established from root suckers, which are produced in abundance from the existing root system (McCarthy and Rytter 2015; Hytönen 2018). Dense, sprout originated hybrid aspen stands could also be grown for bioenergy applying much shorter rotations than those recommended for pulpwood (Tullus et al. 2012; Hytönen 2018). Another option would be a combination of short rotation cultivation for bioenergy or pulpwood, followed by selective thinning, and production of larger dimension trees for veneer or sawn timber (McCarthy and Rytter 2015).
Hybrid aspen grows best on fertile forest sites and former agricultural land with good aeration and water conditions (Lutter et al. 2017). At the most suitable sites in Southern Finland the yield of the first rotation stands can be up to 20 m3 ha–1 per year in rotations of around 20 years (Beuker et al. 2016). Growth of the second rotation is expected to be even better due to higher stand density and vigorous initial growth of the coppice due to the existing root system. Hybrid aspen is very susceptible to damage by mammal herbivores. Herbivore damage may not only affect the survival and growth rate of the plantations, but also has a major effect on stem and wood quality. The plantations should be fenced against moose and the individual trees protected against voles and hares. Failures of earlier hybrid aspen plantations were mainly due to unsuitable origin of the planting material, unsuitable site conditions, as well as poor management at establishment phase and seedling stage or damage by browsing mammals (Viherä-Aarnio 1999).
The properties of the biomass feedstock can be determined by various characteristics, depending on the end use. Wood density is considered to be one of the most important quality traits for the usefulness for timber, as a high density has a positive effect on most of the mechanical strength properties (Heräjärvi 2004a,b). Native European aspen is known to have several favourable properties regarding mechanical processing (Söyrilä 1992; Heräjärvi et al. 2006), but studies on the corresponding properties of hybrid aspen are limited (Heräjärvi et al. 2006; Heräjärvi 2009). Energy yield, measured as heating value, is one of the most important wood quality characteristics for energy plantations (Kenney et al. 1990). The ash content of biomass is known to vary between tree species and tree components (Hakkila and Kalaja 1983; Voipio and Laakso 1992; Hytönen and Nurmi 2015). High wood density entails a higher heating value per volume, whereas high ash content decreases the heating value of biomass (Sheng and Azevedo 2005). In addition, high amounts of ash can also affect the clogging of the ash handling mechanisms of power plants and may lead to higher cleaning and maintenance requirements of the boilers (Niu et al. 2016).
In order to know if wood quality traits can be improved through the breeding and selection of superior hybrid aspen clones, more knowledge is needed about the genetic variation as well as the within-tree variation of those traits.
The main objective of this study was to determine the clonal variation of ash and moisture content, basic density and heating value among seven selected hybrid aspen clones growing on former agricultural land in southern Finland. In addition, we examined the variation of those traits within the trees between wood, bark and branches, as well as the vertical variation for wood and bark.
2 Material and methods
2.1 Field trial and clones
The material for this study was obtained from a hybrid aspen clonal trial in Urjala, southern Finland (61°00´N, 23°29´E) that was established in spring 1998. The trial included 25 micro propagated hybrid aspen clones. The clones originated from trees that had been selected phenotypically from stands and trials that were established in southern and central Finland during the 1950s and 1960s. The selected trees were superior in growth, had good stem quality (straight with few branches) and showed no signs of diseases. The one-year-old containerized hybrid aspen plants were grown in the Haapastensyrjä nursery of the Foundation of Forest Tree Breeding. The experimental layout was a randomized complete block design with five replicates and ten trees per plot (two rows of five trees per clone per plot with a spacing of 2 m (within rows) × 4 m (between rows)). The trial was established on a former agricultural field with a sandy clay soil. Before planting, the soil was prepared by ploughing. No weed control was carried out before or after planting. The site was fenced against moose and each individual tree was protected against voles and hares by a 50 cm high Tubex tube.
For this study, seven hybrid aspen clones were selected by phenotype (Table 1). The selected clones were amongst the ten best clones for height growth in the trial. However, because in this study the total tree biomass was studied, in addition to growth, the clones were also selected to be different in branching intensity, based on exterior observation. The share of branches from total mass in the clones of hybrid aspen varied from 30 to 43%, the mean being 34%.
| Table 1. The hybrid aspen clones included in the study, the location of the stand they were selected from and the origin of their parents, as accurately as could be recovered from the old documents. | |||||||
| Clone | Location of selection | Latitude | Longitude | Mother species | Origin of the mother | Father species | Origin of the father |
| 7 | Vihti | 60°20´ | 24°26´ | Populus tremula | Tuusula, Finland | Populus tremuloides | Aleza Lake, Cariboo, BC, Canada |
| 14 | Lapinjärvi | 60°37´ | 26°11´ | Populus tremula | Toivakka, Finland | Populus tremuloides | Maple, Ontario, Canada |
| 20 | Nurmijärvi | 60°30´ | 24°42´ | Populus tremuloides | Aleza Lake, Cariboo, BC, Canada | Populus tremula | Finland |
| 21 | Vihti | 60°20´ | 24°26´ | Populus tremula | Helsinki, Finland | Populus tremuloides | Galt, Ontario, Canada |
| 24 | Vaajakoski | 62°15´ | 25°54´ | Populus tremula | Tuusula, Finland | Populus tremuloides | Maple, Ontario, Canada |
| 26 | Vaajakoski | 62°15´ | 25°54´ | Populus tremuloides | Gothenburg botanical garden, Sweden | Populus tremula | Tuusula, Finland |
| 27 | Loppi | 60°37´ | 24°27´ | Populus tremula | Punkaharju, Finland | Populus tremuloides | Canada |
2.2 Field measurements and sample collection
One tree per clone was selected at random from each block (total five trees/clone) and harvested during winter (December–January) after 12 growing seasons. However, due to mortality, two trees of clone 21 had to be taken from block 1 and two trees of clone 27 from blocks 1 and 4. After felling, height (dm) and diameter at breast height (DBH, cm) of each tree were measured. The trees were delimbed and the branches of each tree were collected, weighed and bundled. A sample consisting of 10 cm sections from top, middle and base sections of each branch bundle was taken for analysis. From the stem, 5 to 6-cm-thick sample discs were sawn at a height of 0.6 m, 1.3 m and 2 m from the stump and from there on every two metres to the top of each tree. Only two trees were tall enough to be sampled at 12 metres height. Because there were only two 12 m samples they were excluded from the data. All samples were marked (tree number, sampling height) and packed in plastic bags.
2.3 Assessment of ash and moisture content, basic density and heating value
In the laboratory, the diameter with and without bark of each disc was measured in two directions perpendicularly. Subsequently, the stem sample discs were divided horizontally into two discs (2.5–3 cm in thickness). Of these, one disc was used for determining basic density using the water replacement method (Olesen 1971). After measuring the total volume with bark of these discs, they were debarked (outer and inner bark) and re-measured to obtain the volume of wood. The volume of bark was calculated on the basis of these two measurements. Then, the bark and wood samples were dried at +105 °C to constant weight and weighed, and moisture content and basic density calculated separately for wood and bark. One basic density measurement for bark was omitted from the analysis due to a measurement error.
The other disc halves, as well as the branch sample, were used for analysing the ash content and heating value of wood, bark and branches. These sample discs were debarked carefully. Then the wood and bark samples from all disks belonging to one and the same tree were united to form one wood and one bark sample per tree, dried at +70 °C to constant weight and milled.
Calorific heating value (qp(gross)) was determined with an IKA C 500 calorimeter. In order to calculate the effective (qp(net)), lower, net calorific value on a dry basis) heating value it is necessary to known the hydrogen content of the compartments, since the heat of condensation of water vapour created during combustion has to be taken into account. The hydrogen concentrations used were obtained from Nurmi (1997) and for branches and bark they were calculated as means from figures presenting values for different sized branches and inner and outer bark (stem 6.32%, branches 6.07%, bark 6.23%). The following formula was used in the calculation (see Alakangas et al. 2016):
qp(net) = qp(gross) – 212.2 × w(H)d – 0.8 × [w(O)d + w(N)d]
qp(net) = net (effective) calorific value on a dry basis at constant pressure, kJ kg–1
qp(gross) = gross calorific value on a dry basis, kJ kg–1
w(H)d = hydrogen content in dry biofuel, %
w(O)d = oxygen content in dry fuel, %
w(N)d = nitrogen content in dry fuel, %
For w(O)d + w(N)d default value of 41% was used (Alakangas et al. 2016). Ash content of the samples was determined by ashing the samples at 550 °C for 8 hours.
2.4 Statistical analyses
The basic density and moisture content of stem wood, stem bark and the whole stem (wood plus bark) for each tree was calculated as a mean weighed by diameter squared at each measurement point at different heights. The clonal differences in the wood, bark and stem basic density, moisture content, heating value and ash content were tested with mixed linear models with the clone as fixed effect, replication as random effect and tree height as covariate. The clonal means were compared using Student’s LSD test at significance level p < 0.05. The effect of clone and sampling height on basic density and moisture content of wood, bark and stem was tested with repeated measures analysis of variance, where sampling height was used as a within-subject variable. Due to several trees being shorter than 10 m, the analysis was confined to 8 m of height. When in the repeated ANOVA models Mauchly’s test indicated that the assumption of sphericity had been violated, the degrees of freedom were corrected using Greenhouse-Geisser estimates of sphericity. When testing figures in percentage terms, variance stabilizing transformations of square root arcsine was used. The pairwise linear dependences between the studied traits were examined with Pearson’s linear correlation coefficients calculated on the clonal means. All analyses were done using the IBM SPSS 22.0 programme.
3 Results
3.1 Tree height and diameter
The mean height of all trees was 10.6 m and their DBH 10.8 cm (Fig. 1). There were significant differences in the height of the clones, but not in their DBH.
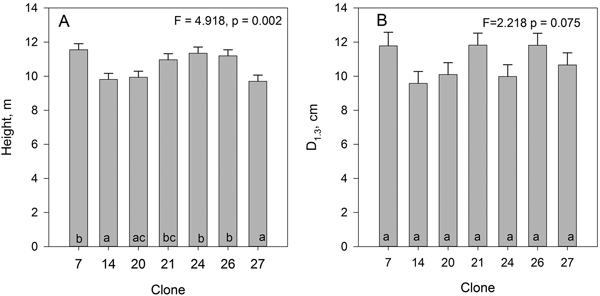
Fig. 1. Mean height (A) and DBH (B) of the studied hybrid aspen clones. Bars indicate standard error. Clones marked with the same letters do not differ from each other at the 0.05 significance level.
3.2 Proportion of wood and bark
The clones differed in wood (and bark) fraction in the stems (Fig. 2). The amount of wood in the hybrid aspen stems was on average 84.9%, with a range of 8 percentage points between clones. The wood fraction of the stem biomass remained quite constant until up to 6 m height after which it started to decrease (Fig. 2). The decrease of the wood fraction towards the top of the tree was significant. There was also a significant sampling height × clone interaction for the fraction of stem wood. For clonal means there was a strong positive correlation (r = 0.883, p = 0.008) between tree height and the proportion of wood in the stems.
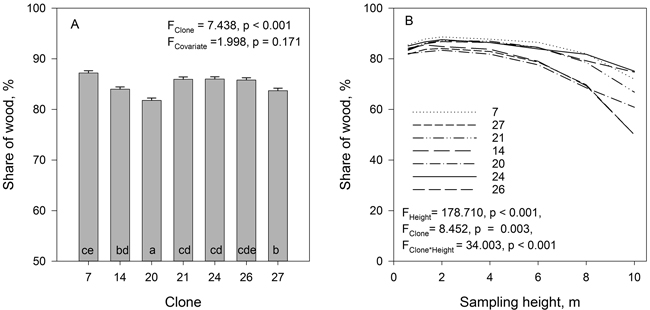
Fig. 2. The mean wood fraction of the total stem biomass (A) and the wood fractions of stem biomass at different sample heights (B) in the studied hybrid aspen clones. Tree height was used as covariate. Clones marked with the same letters do not differ from each other at the 0.05 significance level.
3.3 Ash content
The ash content was lowest in hybrid aspen wood with a mean value for all clones of 0.45%. There were clonal differences in ash content of the wood (Fig. 3). Clone 21 had the lowest ash content (0.40%) and clone 26 the highest (0.49%). Ash content of the bark was almost 10 times higher than that of the wood (mean of all clones 3.9%). Clone 20 had exceptionally low ash content in the bark compared to other clones (Fig. 3). The ash content of the branches (mean of all clones 2.4%) was intermediate to that of the wood and bark. Ash content of the wood and bark did not correlate significantly with the size of the trees (DBH, H). However, because the ash content of the bark was much higher than that of the wood, the ash content of branches correlated negatively with DBH (r = –0.475, p = 0.004) and H (r = –0.358, p = 0.035). Bigger trees had bigger branches and thus relatively less ash rich bark in the branches. When calculated on clonal means, no significant correlation was found between the ash content of wood and ash content of bark.
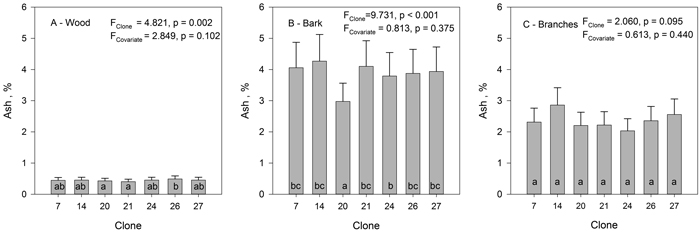
Fig. 3. The mean ash content of wood (A), bark (B) and branches (C) in hybrid aspen clones. Lines inside bars indicate standard error. Tree height was used as a covariate. Clones marked with the same letters do not differ from each other at the 0.05 significance level. View larger in new window/tab.
3.4 Moisture content
The moisture content of the hybrid aspen clones was highest in the wood (on average 54.9%), lowest in the bark (48.9%) and intermediate in the branches (50.1%) (Fig. 4). The average moisture content of the whole stem without branches (54.3%) was only slightly lower than that of the wood fraction due to the small share of bark of the total stem biomass. There were significant clonal differences in the moisture content for wood, bark, stem as well as branches. For wood moisture content the covariate (height) was significant indicating that bigger trees had lower moisture content. No significant correlation was found between the moisture of wood and moisture of bark, when calculated on clonal means.
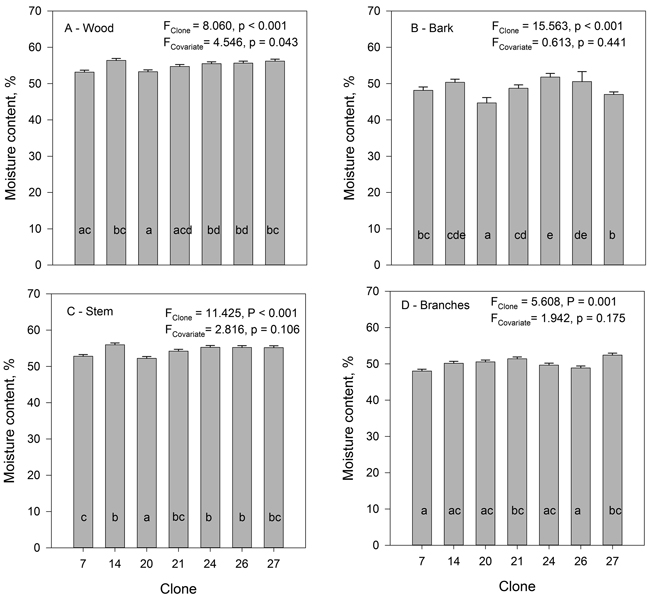
Fig.4. The mean moisture content of wood (A), bark (B), stem (wood and bark) (C) and branches (D) of the studied hybrid aspen clones. Tree height was used as covariate. Clones marked with the same letters do not differ from each other at the 0.05 significance level.
The effect of sampling height on wood, bark and stem moisture contents was significant (Fig. 5). However, wood and bark moisture contents changed differently in the vertical direction. The moisture content of wood was lowest at the base of the stem and increased in a vertical direction reaching its maximum at 2–4 m height, then gradually decreasing further upwards and in most of the clones reaching its minimum at top of the trees (Fig. 5). The bark moisture content was lowest at the base of the stem and increased consistently towards the top of the tree. The moisture of stem (wood and bark) was lowest at the stem base. However, vertical changes in stem moisture were smaller than those observed in wood and bark since the moisture of the wood was low and that of bark high at the top of the stems. There were significant clonal differences in the moisture content of the wood, bark and stem. The clone × sampling height interaction was significant for wood and stem moisture content indicating that the studied clones had different moisture content at different heights.
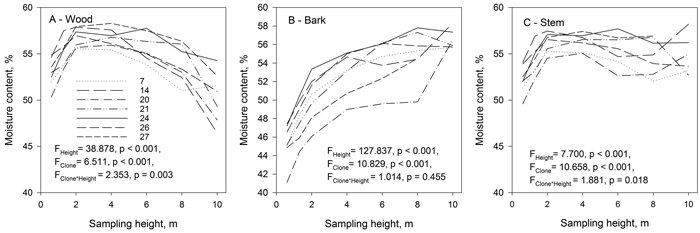
Fig. 5. The mean moisture content of wood (A), bark (B), and stem (wood and bark) (C) of the studied hybrid aspen clones at different sampling heights. View larger in new window/tab.
3.5 Basic density
The average basic density of wood for all clones was 378 kg m–3 and that of bark 450 kg m–3. There were significant clonal differences in basic density both for wood and bark (Fig. 6). The difference between the clones with the highest and lowest basic density of wood and bark was 44 kg m–3 and 91 kg m–3, respectively. Correlation between the density of wood and density of bark was not significant, when calculated on the clonal means. For clonal means there was, however, a strong negative correlation between tree height and density of wood (r = –0.796, p = 0.032), as well as between height and density of stem (r = –0.832, p = 0.020).
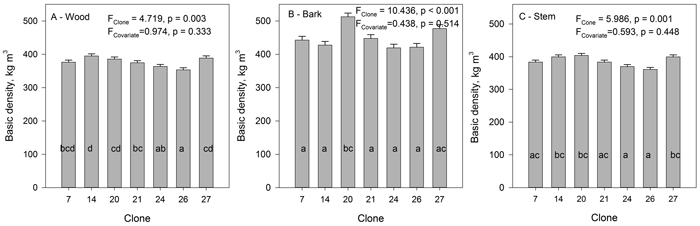
Fig. 6. The mean basic density of wood (A), bark (B) and stem (wood and bark) (C) of the studied hybrid aspen clones. Standard error of the mean. Tree height was used as covariate. Clones marked with the same letters do not differ from each other at the 0.05 significance level. View larger in new window/tab.
The effect of sampling height on wood, bark and stem basic density was significant (Fig. 7). The basic density of hybrid aspen clones decreased from the base of the stem up to 4–6 m height, but increased again further upwards toward the top of the tree (Fig. 7). The clonal variation was significant as was the clone × sampling height interaction. Clone 24 had at the lower parts of the stem exceptionally low basic density (Fig. 7). The variation in basic density of bark was higher than that of wood. Basic density of bark was highest close to the ground and generally decreased up to 4 m, slightly increasing again towards the top of the trees.
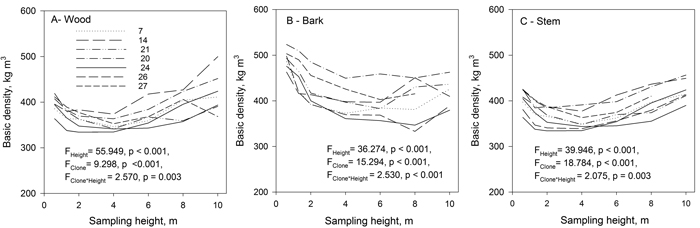
Fig. 7. Mean basic density of wood (A), bark (B), and stem (wood and bark) (C) of the studied hybrid aspen clones at different sampling heights. View larger in new window/tab.
3.6 Heating value
Analysis of covariance showed clonal differences in the effective heating value for both stem wood and stem bark of hybrid aspen clones (Fig. 8). For wood, also the covariate (tree height) was significant. However, there were no significant differences in the effective heating value for the branches of the different clones. The effective heating value of wood was lower than that of bark, and that of branches was intermediate. The average effective heating value of wood was 18.264 MJ kg–1, bark 19.239 MJ kg–1 and that of branches 18.747 MJ kg–1. The difference in heating value between the clones with the lowest and highest value of the wood was 2.4% and that of bark 4.9%. Clone 27 had the highest heating value in wood, but lowest in bark. For clonal means, no significant correlation was found between the wood density and heating value of wood, as well as between the bark density and heating value of bark. Neither did heating values of wood and bark correlate significantly.
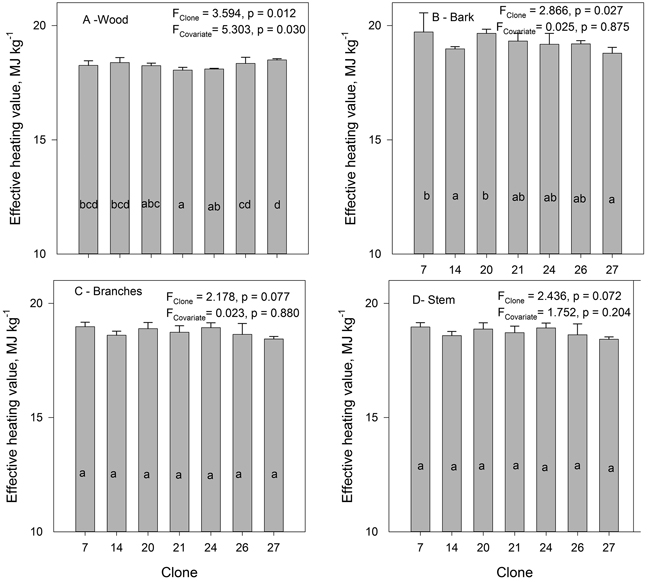
Fig. 8. The mean effective heating value of oven dry biomass of wood (A), bark (B), branches (C) and stem (wood and bark) (D) of hybrid aspen clones. Tree height was used as covariate.
4 Discussion
After 12 growing seasons, the aspens were on average 11 m in height. The trial in Urjala was one of a series of trials in southern Finland including the same clones. Compared to the other trials in this series, the growth was lowest at this site (data not shown). The poorer growth was probably mainly due to a lack of weed control and, consequently, competition from weeds especially during the first year after planting. If no weed control is conducted, competition from weeds on former agricultural lands is strong (Hytönen and Jylhä 2005, 2013) and it can have a significant effect on the survival and growth of Populus seedlings (Böhlenius and Övergaard 2015).
Stem biomass is composed of wood and bark. The proportions of these components vary with the diameter and height of the tree. Usually, the majority of the stem mass is concentrated in the wood, which makes it the single most important component to consider when ash content or heating value for the whole stem is calculated (Nurmi 1993). The amount of bark produced can be also important since the interest for secondary metabolites is growing and aspen bark could serve as a source of natural antioxidant agents under certain conditions (Diouf et al. 2009). When aspen is grown in short-rotations of 5–10 years for biomass production, the dimensions of the harvested trees remain small, and as a result of this the bark fraction can be quite high (Hytönen 2018). In this study, the bark percentage started to increase at the height of 6 m when the diameter of the stems had decreased to 4–7 cm. The average bark percentage (15%) was similar to that reported by Tullus et al. (2009) for hybrid aspens 7–8 cm at breast height. Bark percentage reached 25–50% at the top of the stems. Similar bark percentage has been reported for trees being 1–2 cm by DBH (Tullus et al. 2009).
We found clonal differences in the basic density of wood and bark in our 12-year-old hybrid aspens in accordance with the results of Rytter and Stener (2003) from 14-year-old hybrid aspens. The range between clones in the density of bark (91 kg m–3) was much greater than the range in the density of wood (44 kg m–3). The range in basic density reported in this study was in accordance with the values reported in most of the earlier studies (Illstedt and Gullberg 1993; Rytter and Stener 2003; Heräjärvi and Junkkonen 2006; Zeps et al. 2016). However, Hart et al. (2013) and Heräjärvi (2009) have measured higher basic density values of wood for hybrid aspen. No significant correlation was detected between wood density and tree diameter, and in the analysis of covariance the covariate (tree height) was not significant. However, clonal means of wood and stem density correlated negatively with height in our study, i.e. the fast growing clones had lower average wood density than the slow-growing ones. This result is in line with Heräjärvi and Junkkonen (2006) who found the fastest growing aspen trees having the lowest average wood density, although their study did not include clonal comparisons. On the other hand, Zeps et al. (2016) and Tsoumis (1991) have suggested that growth rate should have no significant influence on the wood density of diffuse porous species such as aspen. Our results suggest that selection for fast growth in hybrid aspen could lead to a decrease in wood density. However, further studies using larger sample of clones would be needed to verify this result.
The basic density of hybrid aspen bark, wood and stem decreased from the base of the stem up to 4–6 m height, but started to increase again further up toward the top of the tree. A similar trend was observed earlier by Heräjärvi and Junkkonen (2006) who also reported lowest average wood density at the height of 2–8 metres from the base in their 32-year-old trees. Accordingly, wood density has been reported to be highest at the stem base and within the living crown also in European aspen (Kärki 2001; Heräjärvi and Junkkonen 2006). Wood density of hybrid aspen varies also in the horizontal direction even though within-stem density variation is more uniform than in European aspen (Heräjärvi and Junkkonen 2006). Density in hybrid aspen is lowest 2–6 cm from the pith and increases towards cambium and wood under the bark (Heräjärvi and Junkkonen 2006; Heräjärvi et al. 2006). According to Kärki (2001) wood density of European aspen is higher directly under the bark than in the piths at tree heights up to 12 meters.
Higher effective heating values for the bark compared to the wood have been reported for many tree species, e.g. Scots pine, downy birch and silver birch (Nurmi 1993, 1997, 2000). In this study, this was found to be true also for hybrid aspen, the bark having a 5.5% higher effective heating value than the wood. Tullus et al. (2009) reported hybrid aspen having higher calorific value in stembark than in stemwood. In contrast, Nurmi reported small European aspen trees having a lower effective heating value for bark than for wood (Nurmi 1993), whereas mature trees had a higher effective heating value for bark than for wood (Nurmi 1997). However, the material of Nurmi (1993) consisted of only two European aspen sample trees while our data consists of 35 trees thus reducing tree-to-tree variation. In our data, one tree had bark with a slightly lower effective heating value than the wood.
In general, the hybrid aspen stemwood and stembark effective heating values (MJ kg–1) in this study were comparable with those measured for native aspen, but lower than measured for silver birch, downy birch, grey alder and black alder (Nurmi 1993). Birch and alder bark have a considerably higher heating value than that found in hybrid aspen or European aspen (Nurmi 1993). The changes in the amounts of bark and wood proportions often have a significant effect on the effective heating value of the stem. The older and bigger trees are, the smaller their bark percentage is. Since the clonal differences in heating value were small in magnitude (0.45 MJ kg–1 for wood, 0.93 MJ kg–1 for bark), the selection of clones for biomass cultivation on the basis of the heating value does not seem feasible according to our results.
The ash content varies according to species, tree age and tree components (Hakkila and Kalaja 1983; Voipio and Laakso 1992; Hytönen and Nurmi 2015). Wood had much lower ash content (0.45%) than bark (3.9%), which is in accordance with other studies. There was also significant clonal variation in the ash content. The differences in ash content between clones having highest and lowest ash content in wood, bark and branches was 21%, 38% and 41% respectively. Ash content of hybrid aspen wood was similar to that reported earlier for 7-year-old hybrid aspens and for downy birch, silver birch and grey alder wood (0.3–0.6%) (Hakkila and Kalaja 1983; Voipio and Laakso 1992; Tullus. et al. 2014; Hytönen and Nurmi 2015). Much higher wood ash content has been measured in young short-rotation willows (0.8–1.7%) (Lyons et al. 1986; Senelwa and Sims 1999; Hytönen and Nurmi 2015). The ash content of hybrid aspen stem bark (3.9%) was higher than that of the species growing in the Nordic countries reported as having a bark ash content of between 2.5–3.5% (Hakkila and Kalaja 1983,Voipio and Laakso 1992) and much higher than that reported for young downy and silver birch trees (1.5–1.6%) (Hytönen and Nurmi 2015). The ash content of hybrid aspen bark was even higher than that reported for young short-rotation willow (2.7–3.6%) (Lyons et al. 1986; Dzurenda 2010a,b; Hytönen and Nurmi 2015). High ash content, as well as high contents of Ca, Cu, K, Mg were reported by Kuusinen (1996) in the bark of European aspen. The high variation in ash content between the studied hybrid aspen clones could be related to clonal differences in their nutrient concentrations. E.g. Rytter and Stener (2003) reported clonal variation in stem concentrations of K, P and Mg. Since hybrid aspen is considered to be nutrient demanding species (Hytönen 2018) the clonal variation in nutrient use efficiency should be studied.
Many factors affect the moisture content of trees and it varies e.g. by species, season, growing site, and within the tree. Aspen has highest moisture content in autumn and in the winter (Hakkila 1962). Hybrid aspens cut in the winter and in autumn (Tullus et al. 2009) had lower moisture content in the bark than in the wood. The seasonal variation in downy birch wood and bark is high, and wood and bark have on the average similar moisture content (Ferm 1985). The moisture content of hybrid aspen wood was lowest close to the base of the stem and reached its maximum at 2–4 m height, after which it gradually decreased further upwards and in most of the clones reached its minimum at 10 m height. The bark moisture content was lowest at stump level and increased towards the top of the tree. A similar, but much smaller increase towards the top of the tree has been observed also in downy birch (Ferm 1985). The difference in moisture content between hybrid aspen clones having lowest and highest moisture content was 12%, 8% and 9% in wood, bark and branches, respectively. Most often moisture content is the determining factor in thermal conversion affecting energy yield (Nurmi and Hillebrand 2007). Nevertheless, moisture content is not a feasible criterion for clonal selection, because after felling of trees it is dependent on post-harvest handling and storage conditions.
Significant differences among the studied 12-year-old hybrid aspen clones were detected in growth, moisture content, ash content, basic density and heating value of wood and bark, as well as in the share of wood and bark in their stems. Although based on a limited material (seven clones in one trial), our results suggest that these biomass characteristics could be improved by clonal selection, which could improve the suitability of hybrid aspen for various end uses. An advantage of clonal selection is that the total genetic variation can be used. However, when selecting for various characteristics also the correlation between characteristics should be taken into account (Bisoffi and Gullberg 1996; Yu et al. 2001). In this study the negative correlation detected between height growth and wood density indicated that selection for fast growth may have a negative effect on wood density in hybrid aspen.
Acknowledgements
Jussi Tiainen, Heikki Kinnunen, Seppo Vihanta and Eero Saari are acknowledged for the field work and Seppo Vihanta for assisting in data processing. Reetta Kolppanen and Kaisa Jaakola at Luke Kannus and Kokkola laboratories are thanked for analysis.
References
Alakangas E., Hurskainen M., Laatikainen-Luntama J., Korhonen J. (2016). Properties of indigenous fuels in Finland. VTT Technology 272. 222 p. http://www.vtt.fi/inf/pdf/technology/2016/T272.pdf.
Beuker E. (2000). Aspen breeding in Finland, new challenges. Baltic Forestry 6(2): 81–84.
Beuker E., Viherä-Aarnio A., Hynynen J. (2016). Growth potential of first generation hybrid aspen plantations in southern Finland. In: Poplars and other fast-growing trees – Renewable resources for future green economies, International Poplar Commission, 25th session Berlin, Germany, 13–16 September 2016: Abstracts of submitted papers and posters. Working Paper IPC 14. 118 p.
Bisoffi S., Gullberg U. (1996). Poplar breeding and selection strategies. In: Stettler R.F., Bradshaw Jr. H.D., Heilman P.E., Hinckley T.M. (eds.). Biology of Populus and its implications for management and conservation. Part I, Chapter 6. NRC Research Press, National Research Council of Canada, Ottawa, ON. p. 139–158.
Böhlenius H., Övergaard R. (2015). Growth response of hybrid poplar to different types and levels of vegetation control. Scandinavian Journal of Forest Research 30(6): 516–525. https://doi.org/10.1080/02827581.2015.1034768.
Diouf P.N., Stevanovic T., Cloutier A. (2009). Antioxidant properties and polyphenol contents of trembling aspen bark extracts. Wood Science Technology 43(5–6): 457–470. https://doi.org/10.1007/s00226-009-0240-y.
Dzurenda L., Geffertová J., Hecl V. (2010a). Energy characteristics of wood-chips produced from Salix viminalis – clone ULV. Drvna Industrija 61(1): 27–31. http://hrcak.srce.hr/49204.
Dzurenda L., Geffertová J., Zliak M. (2010b). Energetické vlastnosti štiepky plantažnicky pestovanej dreviny Salix viminalis klon – RAPP. Abstract: Energy characteristics of the wood-chip produced from Salix viminalis – clone RAP. Acta Facultatis Xylologiae Zvolen 52(1): 85–91. https://stary.tuzvo.sk/files/DF/fakulta_df/AFX/archive/2010/2010-1/10-1-10-dzurenda-geffertova-zoliak.pdf.
Ferm A. (1985). Pienikokoisen hieskoivun kosteuden ja tiheyden vaihtelu turvemaalla. Abstract: Variation in the water content and basic density of small-sized pubescent birch (Betula pubescens) stems on peatland. Metsäntutkimuslaitoksen tiedonantoja 206: 19–39. http://urn.fi/URN:ISBN:951-40-0902-9.
Hakkila P. (1962). Kesäaikana valmistetun haapapaperipuun kuivumisesta ja lahoamisesta. Summary: On the seasoning and decay of aspen pulpwood prepared in summertime. Communicationes Instituti Forestalis Fenniae 54(6). 34 p.
Hakkila P., Kalaja H. (1983). Puu- ja kuorituhkan palauttamisen tekniikka. Summary: The technique of recycling wood and bark ash. Folia Forestalia 552. 37 p. http://urn.fi/URN:ISBN:951-40-0612-7.
Hart J.F., de Araujo F., Thomas B.R., Mansfield S.D. (2013). Wood quality and growth characterization across intra- and inter-specific hybrid aspen clones. Forests 4(4): 786–807. https://doi.org/10.3390/f4040786.
Heräjärvi H. (2004a). Static bending properties of Finnish birch wood. Wood Science and Technology 37: 523–530. https://doi.org/10.1007/s00226-003-0209-1.
Heräjärvi H. (2004b). Variation of basic density and Brinell hardness within mature Finnish Betula pendula and B. pubescens stems. Wood and Fiber Science 36(2): 216–227.
Heräjärvi H. (2009). Effect of drying technology on aspen wood properties. Silva Fennica 43(3): 433–445. https://doi.org/10.14214/sf.198.
Heräjärvi H., Junkkonen R. (2006). Wood density and growth rate of European and hybrid aspen in Southern Finland. Baltic Forestry 12(1): 2–8. https://www.balticforestry.mi.lt/bf/PDF_Articles/2006-12[1]/2_8%20Herajarvi%20&%20Junkonen.pdf.
Heräjärvi H., Junkkonen R., Koivunen H., Metros J., Piira T., Verkasalo E. (2006). Metsä- ja hybridihaapa sahatavaran ja jatkojalosteiden raaka-aineena. [European and hybrid aspen as the raw material of sawn timber and upgraded products]. Working papers of the Finnish Forest Research Institute 31. 102 p. http://urn.fi/URN:ISBN:978-951-40-2008-7.
Hynynen J. (1999). Haavan ja hybridihaavan kasvu ja tuotos. [Growth and yield of aspen and hybrid aspen]. In: Hynynen J., Viherä-Aarnio A. (eds.). Haapa ‒ monimuotoisuutta metsään ja metsätalouteen. Vantaan tutkimuskeskuksen tutkimuspäivä Tammisaaressa 12.11.1998. Metsäntutkimuslaitoksen tiedonantoja 725: 25–37. http://urn.fi/URN:ISBN:951-40-1671-8.
Hytönen J. (2018). Biomass, nutrient content and energy yield of short-rotation hybrid aspen (P. tremula × P. tremuloides) coppice. Forest Ecology and Management 413: 21–31. https://doi.org/10.1016/j.foreco.2018.01.056.
Hytönen J., Jylhä P. (2005). Effects of competing vegetation and post-planting weed control on the mortality, growth and vole damages to Betula pendula planted on former agricultural land. Silva Fennica 39(3): 365–380. https://doi.org/10.14214/sf.374.
Hytönen J., Jylhä P. (2013). Long-term effect of weed control on survival and growth of silver birch planted on arable land. Baltic Forestry 19(2): 170–178. https://www.balticforestry.mi.lt/bf/PDF_Articles/2013-19[2]/Long-term%20Effect%20of%20Weed%20Control%20on%20Survival%20and%20Growth%20os%20Silver%20Birch.pdf.
Hytönen J., Nurmi J. (2015). Heating value and ash content of intensively managed stands. Wood Research 60(1): 71–82. http://www.centrumdp.sk/wr/201501/07.pdf.
Illstedt B., Gullberg U. (1993). Genetic variation in a 26-year old hybrid aspen trial in southern Sweden. Scandinavian Journal of Forest Research 8(1–4): 185–192. https://doi.org/10.1080/02827589309382768.
Kärki T. (2001). Variation of wood density and shrinkage in European aspen (Populus tremula). Holz als Roh-und Werkstoff 59(1–2): 79–84. https://doi.org/10.1007/s001070050479.
Kenney W.A., Sennerby-Forsse L., Layton P. (1990). A review of biomass quality research relevant to the use of poplar and willow for energy conversion. Biomass 21(3): 163–188. https://doi.org/10.1016/0144-4565(90)90063-P.
Kuusinen M. (1996). Epiphytic lichen flora and diversity in old growth boreal forests of Finland. Academic dissertation. University of Helsinki, Department of Ecology and Systematics. 29 p.
Lutter R., Tullus A., Kanal A., Tullus T., Tullus H. (2017). Above-ground growth and temporal plant–soil relations in midterm hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantations on former arable lands in hemiboreal Estonia. Scandinavian Journal of Forest Research 32(8): 688–699. https://doi.org/10.1080/02827581.2017.1278784.
Lyons G.J., Pollock H.P., Hegarty A. (1986). Fuel properties of short-rotation hardwood coppice sprouts. Journal of the Institute of Energy 59: 138–141.
McCarthy R., Rytter L. (2015). Productivity and thinning effects in hybrid aspen root sucker stands. Forest Ecology and Management 354: 215–223. https://doi.org/10.1016/j.foreco.2015.06.015.
Niu Y., Tan H., Hui S. (2016). Ash-related issues during biomass combustion: alkali-induced slagging, silicate melt-induced slagging (ash fusion), agglomeration, corrosion, ash utilization, and related countermeasures. Progress in Energy and Combustion Science 52: 1–61. https://doi.org/10.1016/j.pecs.2015.09.003.
Nurmi J. (1993). Heating values of the above ground biomass of small-sized trees. Acta Forestalia Fennica 236. 30 p. https://doi.org/10.14214/aff.7682.
Nurmi J. (1997). Heating value of mature trees. Acta Forestalia Fennica 256. 28 p. https://doi.org/10.14214/aff.7517.
Nurmi J. (2000). Characteristics and storage of whole-tree biomass. Metsäntutkimuslaitoksen tiedonantoja – The Finnish Forest Research Institute, Research Papers 758. 42 p. http://urn.fi/URN:ISBN:951-40-1715-3.
Nurmi J., Hillebrand K. (2007). The characteristics of whole-tree fuel stocks from silvicultural cleanings and thinnings. Biomass and Bioenergy 31(6): 381–392. https://doi.org/10.1016/j.biombioe.2007.01.010.
Olesen P.O. (1971). The water displacement method. Forest Tree Improvement 3: 3–23.
Rytter L., Stener L.-G. (2003). Clonal variation in nutrient content of woody biomass of hybrid aspen (Populus tremula L. × P. tremuloides Michx.). Silva Fennica 37(3): 313–324. https://doi.org/10.14214/sf.491.
Sheng C., Azevedo J.L.T. (2005). Estimating the higher heating value of biomass fuels from basic analysis data. Biomass and Bioenergy 28(5): 499–507. https://doi.org/10.1016/j.biombioe.2004.11.008.
Senelwa K., Sims R.E.H. (1999). Fuel characteristics of short rotation forest biomass. Biomass and Bioenergy 17(2): 127–140. https://doi.org/10.1016/s0961-9534(99)00035-5.
Syörilä P. (1992). Haapa viilun ja vanerin raaka-aineena. [Aspen as the raw material of veneer and plywood]. Paperi ja Puu – Paper and Timber 74(8): 621–627.
Tsoumis G. (1991). Science and technology of wood. Structure, properties and utilization. Van Norstand Reinhold, New York. 494 p.
Tullus A., Sellin A., Kupper P., Lutter R., Pärn L., Jasinska A.K., Alber M., Kukk M., Tullus T., Tullus H., Lõhmus K., Sõber A. (2014). Increasing air humidity – a climate trend predicted for northern latitudes – alters the chemical composition of stemwood in silver birch and hybrid aspen. Silva Fennica 48(4) article 1107. https://doi.org/10.14214/sf.1107.
Tullus A., Rytter L., Tullus T., Weih M., Tullus H. (2012). Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scandinavian Journal of Forest Research 27(1): 10–29. https://doi.org/10.1080/02827581.2011.628949.
Tullus A., Tullus H., Soo T., Pärn L. (2009). Above-ground biomass characteristics of young hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantations on former agricultural land in Estonia. Biomass and Bioenergy 33(11): 1617–1625. https://doi.org/10.1016/j.biombioe.2009.08.001.
Viherä-Aarnio A. (1999). Hybridihaapa – 40 vuoden takaa uudeksi viljelypuuksi. [Hybrid aspen ‒ a new crop tree after 40 years]. In: Hynynen J., Viherä-Aarnio A. (eds.). Haapa ‒ monimuotoisuutta metsään ja metsätalouteen. Vantaan tutkimuskeskuksen tutkimuspäivä Tammisaaressa 12.11.1998. Metsäntutkimuslaitoksen tiedonantoja 725: 13–23. http://urn.fi/URN:ISBN:951-40-1671-8.
Voipio R., Laakso T. (1992). Pienikokoisten puiden maanpäällisen biomassan kemiallinen koostumus. Summary: Chemical composition of the above ground biomass of small-sized trees. Folia Forestalia 789. 22 p. http://urn.fi/URN:ISBN:951-40-1202-X.
Yu Q., Pulkkinen P., Rautio M., Haapanen M., Alén R., Stener L.G., Beuker E., Tigerstedt P.M.A. (2001). Genetic control of wood physicochemical properties, growth, and phenology in hybrid aspen clones. Canadian Journal of Forest Research 31(8): 1348-1356. https://doi.org/10.1139/cjfr-31-8-1348.
Zeps M., Gailis A., Smilga J., Miezite O., Sisenis L., Zarija I. (2016). Hybrid aspen clone wood mechanical properties. Agronomy Research 14(S1): 1147–1152.
Total of 47 references.
