Matts Lindbladh  ,
Per-Ola Hedwall,
Ida Wallin,
Annika M. Felton,
Henrik Böhlenius,
Adam Felton
,
Per-Ola Hedwall,
Ida Wallin,
Annika M. Felton,
Henrik Böhlenius,
Adam Felton
Short-rotation bioenergy stands as an alternative to spruce plantations: implications for bird biodiversity
Lindbladh M., Hedwall P.-O., Wallin I., Felton A. M., Böhlenius H., Felton A. (2014). Short-rotation bioenergy stands as an alternative to spruce plantations: implications for bird biodiversity. Silva Fennica vol. 48 no. 5 article id 1135. https://doi.org/10.14214/sf.1135
Highlights
- There is a gap in knowledge regarding the biodiversity implications of replacing production forests with bioenergy stands
- We compared the avian biodiversity of early rotation hybrid aspen stands and spruce plantations, the latter being the dominant production forest type in southern Sweden
- Our results indicate that young hybrid aspen stands can support relatively diverse and distinctive bird communities.
Abstract
Global efforts to decrease dependence on fossil fuels have increased interest in bioenergy production. One source of bioenergy is fast growing deciduous tree species, such as hybrid aspen (Populus × wettsteinii Hämet-Ahti). The majority of research on hybrid aspen which assesses biodiversity implications, has however primarily focused on agricultural lands as the reference condition. This has resulted in a substantial gap in our knowledge regarding the biodiversity implications of replacing production forest types with hybrid aspen, a form of reforestation taking place in northern Europe. In this study we address this knowledge gap by comparing the avian biodiversity of young hybrid aspen and spruce (Picea abies L.) plantations of similar age, the latter being the most prevalent forestry alternative in in southern Sweden. We found that hybrid aspen stands had higher bird species richness and abundance as well as a distinct community composition compared to the spruce stands. We suggest that the most likely driver was the greater structural and tree species complexity in the aspen stands, provided for by the fenced exclusion of ungulates from the regenerating hybrid aspen stands. Our results indicate that at least during early stages of regeneration, and in comparison to the dominating production forest type in the region, hybrid aspen stands can support relatively high levels of bird diversity, and a bird species composition more closely associated with broadleaf habitat types requiring restoration in this region.
Keywords
Norway spruce;
Picea abies;
hybrid aspen;
Sweden;
Populus × wettsteinii;
production forests
-
Lindbladh,
Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden
E-mail
matts.lindbladh@slu.se
- Hedwall, Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail per-ola.hedwall@slu.se
- Wallin, Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail ida.wallin@slu.se
- Felton, Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail annika.felton@slu.se
- Böhlenius, Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail henrik.bohlenius@slu.se
- Felton, Southern Swedish Forest Research Centre, SLU – Swedish University of Agricultural Sciences, P.O. Box 49, SE-230 53 Alnarp, Sweden E-mail adam.felton@slu.se
Received 6 March 2014 Accepted 26 September 2014 Published 30 December 2014
Views 219164
Available at https://doi.org/10.14214/sf.1135 | Download PDF

1 Introduction
Efforts are being made to decouple the global economy from carbon intensive energy sources. Sweden has a policy of increasing renewable energy sources to at least 50% of all energy usage by 2020 (www.naturvardsverket.se/en/). Such policy targets, as well as an increased recognition of the potential benefits for diversifying production landscapes in a changing climate (Felton et al. 2010), has led to a larger interest in bioenergy production. One source of wood fibres for bioenergy production is fast growing deciduous tree species. Hybrid aspen (Populus × wettsteinii Hämet-Ahti), a cross between the European Populus tremula (L.) and the American P. tremuloides (Michx.), is considered to be one of the fastest growing deciduous species in Europe and is regularly planted for bioenergy production (Tullus et al. 2007; Tullus et al. 2012). Whereas previously in Sweden the planting of hybrid aspen stands was primarily considered for use on agricultural lands, stands are now also being established on lands once occupied by wood production stands of Norway spruce (Picea abies L.), hereafter “spruce” (Rytter and Stener 2011), the dominating tree species in Sweden.
For each tree species considered in production forestry, and especially regarding introduced tree species and hybrids, it is important for the landowner and society to weigh the relative advantages and disadvantages of each choice, and thereby limit potential conflicts between environmental objectives (Lindenmayer et al. 2012). In the case of hybrid aspen there are potential adverse ecological outcomes, due to its partially non-native origin (Felton et al. 2013). However, there is also the potential for win-win benefits, primarily associated with their being a broadleaf production forest alternative in this region where there are initiatives to increase the availability of broadleaved tree species, including native aspen (Gustafsson et al. 2014). For this reason, the planting of hybrid aspen may be beneficial because of both its contribution to climate change mitigation and its potential capacity to increase stand or landscape-level biodiversity.
In this study we compared the avian biodiversity of two production forest alternatives established on forest land, and at early stages of stand development: 1) short-rotation hybrid aspen stands, and 2) the dominating forest type in the region, spruce plantations. We refer to these production-forest alternatives as ‘aspen stands’ and ‘spruce stands’ respectively. Hence, we do not compare two natural forest types, but the different biodiversity implications of two forest management alternatives. There is empirical support for expecting conifer and broadleaf stands to differ in bird community composition (Bibby et al. 1989; Peck 1989; Roberge and Angelstam 2006; Sweeney et al. 2010), but more directly relevant data is unfortunately lacking. In a recent literature review on hybrid aspen (Tullus et al. 2012), only four studies were identified which assessed biodiversity, all of which focused on understorey vegetation (Weih et al. 2003). Moreover, the majority of ecological research on fast-growing broadleaf trees has so far relied upon agricultural lands as the reference condition (Sage 1998; Berg 2002; Schulz et al. 2009; Baum et al. 2012).
We had two primary aims: first to assess differences in the bird communities of the two forest management models considered; and second, to determine the extent to which specific management decisions and interventions, such as the landscape context and green-tree retention levels, influenced bird community diversity and composition.
2. Material and methods
2.1 Study area and species
Norway spruce is the most common tree species in southern Sweden (approximately 45 % of total volume). Spruce forests are generally planted as monocultures, and then pre-commercially and commercially thinned of mostly pioneer broadleaves two to three times, with the aim of having a monoculture at the end of the 50–70 years rotation period. There are strong production incentives for planting hybrid aspen, due to its high production capacity and rotation periods of just 20–30 years (Tullus et al. 2007; Tullus et al. 2012) and the interest among forest owners in hybrid aspen has increased in recent years (Rytter and Stener 2011).
2.2 Study sites
After the hurricane Gudrun in January 2005 in Southern Sweden (Valinger and Fridman 2011), a large number of hybrid aspen and spruce stands were planted on sites where the previous stand was lost due to wind felling. We identified 13 successfully regenerated hybrid aspen stands and 13 spruce stands in the region which met selection criteria (as follows) (Fig. 1). For simplicity stand types will hereafter be named “aspen stands” or “spruce stands”, based on the management objective, in spite the fact most stands were dominated by birch (as almost all young plantations in the region are before thinning). All aspen stands were fenced and planted soon after the previous forest stand were wind-thrown. Due to the high abundance of browsing ungulates in Sweden, fencing of hybrid aspen stands is considered a necessity to ensure their successful establishment (Ingemarsson et al. 2007; Löf et al. 2010; Rytter and Stener 2011). Fencing of hybrid stands is therefore a practice closely associated with hybrid aspen establishment in this region and needs to be considered as part of this production forest alternative. All spruce stands were unfenced, as is consistent with this production stand type as spruce stands usually require no such protective measures (Bergquist et al. 2009). All stands of both types were between four and eight years since establishment. In order for survey teams to sample two stands per morning, spruce stands were selected to be less than 3 km from a surveyed aspen stand. To minimize the potential for the same individual birds to be counted within two stands, a minimum distance of 700 m between stands was required. The stands were chosen according to shape and size to ensure that sufficient space was available to enclose the bird sampling points, see below. The average size of the aspen and spruce stands, was 6 and 11 hectares respectively.
Fig. 1. Map of Southern Sweden showing the location of the surveyed stands.
2.3 Bird survey techniques
We used point count methodology to assess the bird communities of each stand type (Bibby et al. 2000). Point counts provide an efficient and useful index of bird species composition and abundance, but like all multi-species survey methodologies must be interpreted with caution, due to associated limitations and differences in the presence, availability, and detectability of species with distinct behaviors, during the survey period (Johnson 2008). We thereby use a variety of approaches to minimize sources of detection error.
Four survey points were located within each stand, with provisos that the distance between two survey points was more than 80 m but less than 90 m, and at least 40 m from the stand edge. Points were concentrated within the center of each stand, to reduce the influence of birds using the transition zone of vegetation at the edge of the study site. This constraint also helped to ensure that survey points were not displaced over larger areas in larger stands, which could have increased bird community diversity in such stands due to an increased range of environments surveyed. Survey points were located a priori using aerial photos and the aforementioned decision rules, to avoid onsite selection bias.
We surveyed each of the study sites four times in 2013; twice in early spring and twice in late spring. The early spring surveys were conducted from 16–24 April and late spring surveys were conducted from 28 May–3 June. We chose these survey periods to coincide with annual peaks in singing activity of breeding resident and migrant passerines respectively. Notably, the majority of the tropical migrant passerines surveyed have not arrived in this region at the time of the first survey period. Daily surveys were begun at dawn, at approximately 6:00 am in April and 4:30 am in May/June, and finished at 9:00am and 7:30am respectively. This period overlapped with the daily peak in bird vocal activity. On each survey day, the same person surveyed one aspen stand and one spruce stand. The order in which the stand types were visited each day was varied systematically to ensure that no stands types were weighted towards early morning or late morning survey times. Surveys were only conducted in suitable weather for conducting bird surveys (i.e. minimal wind, no rain), to minimize environmental influences on detectability.
All point count surveys were conducted by persons experienced with both bird identification and point count surveys (AF, ML and Thomas Nyberg) (Farmer et al. 2012). Each point was surveyed for five minutes (Bonthoux and Balent 2012). Distance to individual birds from the observers was estimated, and only those birds deemed to be within 40 m were recorded for each survey point (Alldredge et al. 2007). This threshold distance was chosen to capture only those birds located within the stand, to avoid double counting birds at two survey points, and because it is substantially less than the maximum distance observers are estimated to be able to differentiate the distance to calling birds (i.e. 65 m, see Alldredge et al. 2007). Most identification was made acoustically rather than visually. In cases of uncertainty (e.g. an individual shifting among bird survey points during the survey period), the most conservative estimate of abundance was used. Due to the potential for migrating individuals to skew results, we only included birds exhibiting territorial behavior, as indicated by songs, with the exception in one case where nest and eggs were found. The inclusion of only singing rather than calling birds, should also limit the extent of detection errors. Birds observed flying overhead were not included in the survey. We assessed whether species encountered were included on the Swedish Red List of threatened taxa (Gärdenfors 2010).
2.4 Stand level vegetation structures
We conducted surveys of vegetation structure during late May and early June of 2013, i.e. the same year as the bird survey. Stand characteristics were measured using a quadrat of 127 m2 located at each of the four survey bird points within each studied stand, hence in total 508 m2 per stand. Canopy height was measured as the highest vegetation point within the quadrat, using a laser rangefinder. Woody plant individuals higher than 1 m within the quadrat were counted and identified to species. Because of the very large number of stems they were divided into four diameter classes: 0–2, 2.1–5, 5.1–10 and > 10 cm, measured at stump height (approximately 20 cm). The basal area for each species was calculated from the average diameter of each size class (for > 10 cm the average 15 cm was used), which was multiplied by the number of stems of the species in question in that size class The number of living or dead trees of DBH ≥10 cm within a radius of 40 m of the point center was counted.
2.5 Landscape-level vegetation structures
To assess the influence of landscape context on bird communities within stands we used two separate datasets, one for forest land only and one for all land use classes. The dataset for forest land is a spatially explicit description of forest land developed from satellite imagery and inventory data provided by the National Forest Survey of Sweden, using the kNN-method (k-Nearest Neighbors algorithm) (Reese et al. 2003). For our analysis the volumes from 2010 were provided for individual tree species and were combined to produce broader categories of deciduous volume and coniferous volume.
The dataset for all land classes originated from the Swedish governmental body for geographical information. This data spatially represents all land divided in to distinct categories of land-use classes updated in 2005. We combined the original land-use classifications into broader land-use classes that were more relevant for our purposes (Table 1). Land-use classes found within a 500 m circle surrounding the center of each stand (i.e. the center of the four survey points) were used to define landscape vegetation. This amounted to a total area of approximately 78 ha.
2.6 Data analysis
The abundance of each bird species in a given stand was determined by using the highest value for the number of individuals encountered from any of the four separate surveys conducted in each stand. We adopted this approach as research indicates that true avian abundance is best correlated with maximum rather than average abundance data from repeated surveys (Toms et al. 2006).
The impact of stand type (aspen or spruce stand), stand characteristics and landscape structure on species richness and bird abundance was modelled using Generalized Linear Models (GLM) in R 3.0.1 (R Development Core Team 2013). Both response variables were modelled with a Gamma distribution and log-link. The Gamma GLM relies on the assumption of homoscedasticity. The GLMs were evaluated in relation to this assumption and for other patterns in the residuals by plotting the Pearson residuals against the fitted values.
Before modelling of the effects of environmental variables (Table 1) on species richness and bird abundance a data exploration was performed by plotting the predictors against each other, by Pearson correlation tests and t-tests. Aspen stands had significantly higher total basal area (t = 2.9, p = 0.009), basal area of aspen (t = 7.6, p < 0.001), tree height (t = 4.6, p = p < 0.001) and more tree species (t = 4.3, p = p < 0.001) than spruce stands. To avoid problems with collinearity these variables were excluded from the models of species richness and bird abundance that included stand type. However, to assess the total explained variance of the variables that differed between aspen and spruce stands, a model including only these variables were included in the analyses. The collinearity problem in this model implies, however, that the parameters and probabilities of single variables should be interpreted with care. To avoid over-parameterization by introducing too many variables in a single model the effects of stand characteristics and landscape structure beyond the effects of stand type were modelled in separate GLMs. The same procedure was used for testing if the effect of stand type is dependent on other stand characteristics or on the landscape structure (i.e. interaction effects). Hence six models (Table 2) were generated and compared by the Akaike Information Criterion (Akaike 1973) and R2 (variance explained).
| Table 1. Description of the environmental variables used in the statistical analyses. The variables are: Total basal area per ha (Bas tot); Stems per ha (Stems); Basal area per ha for aspen (Bas asp); Stand height (Height); Total number of living or dead retained trees > 10 cm DBH within 40 m of the four sample points (Ret tree); Number of thinned respectively unthinned stands (Thin); Total number of woody species (Tree spec); Tree evenness, the Shannon’s diversity index of the tree species composition divided by the log of the number of tree species (Tree even); % developed land within a 500 m radius (Devel); % agricultural land within a 500 m radius (Agric); % wetlands within a 500 m radius (Wetl); Volume coniferous trees per ha within a 500 m radius (Conif); Volume deciduous trees per ha within a 500 m radius (Decid). | |||||||
| Spruce stands | Aspen stands | ||||||
| Mean | SD | Count | Mean | SD | Count | ||
| Bas tot (m2 ha–1) | 1.6 | 1.1 | 3.2 | 1.7 | |||
| Stems (no ha–1) | 9241 | 6181 | 10882 | 5748 | |||
| Bas asp (m2 ha–1) | 0.0 | 0.0 | 1.1 | 0.5 | |||
| Height (m) | 3.4 | 1.1 | 5.2 | 0.8 | |||
| Ret tree (no.) | 7 | 8 | 3 | 3 | |||
| Thin | Unthin | 9 | 10 | ||||
| Thin | 4 | 3 | |||||
| Tree spec (no.) | 4.6 | 1 | 6.8 | 1 | |||
| Tree even | 0.42 | 0.13 | 0.48 | 0.09 | |||
| Devel (%) | 0.3 | 0.007 | 0.5 | 0.007 | |||
| Agric (%) | 5 | 0.068 | 1 | 0.106 | |||
| Wetl (%) | 2.3 | 4.2 | 1.3 | 0.028 | |||
| Conif (m3 ha–1) | 100 | 27 | 71 | 30 | |||
| Decid (m3 ha–1) | 24 | 9 | 28 | 14 | |||
| Table 2. The Generalized Linear Models (GLM) of species richness and bird abundance and the underlying hypotheses. | |
| Model | Hypothesis |
| GLM1 | Species richness/abundance differs between stand types. |
| GLM2 | Besides stand type species richness/abundance is also dependent on stand variables that are uncorrelated with stand type (environmental variables 2, 5, 6 and 8 in Table 1). |
| GLM3 | The impact of stand type on species richness/abundance is dependent on stand factors that are uncorrelated with stand type (same variables as GLM2). |
| GLM4 | Besides stand type species richness/abundance is also dependent on landscape composition and forest composition on the landscape level (variables 9–13) |
| GLM5 | The impact of stand type on species richness/abundance is dependent on landscape composition and forest composition on the landscape level. (same as GLM4). |
| GLM6 | The variables that are correlated with stand type are together better predictors of species richness/abundance than stand type itself (variables 1, 3, 4 and 7). |
While R2 values increase with the explanatory power of a model, and with the number of variables included, the AIC penalize model complexity. A decrease in AIC indicates a model improvement and a reduction by approximately two AIC points was used here as an indication of model enhancement. Additionally, single variables were treated as significant if the individual p-value was lower than 0.05.
To analyze the effects of stand type, stand- and landscape characteristics on the bird communities a non-metric multidimensional scaling (NMS) was performed on the bird community data by applying the metaMDS function in the Vegan package (Oksanen et al. 2013) in R 3.0.1 (R Core Team 2013). The correlation between site locations in ordination space and the environmental variables were analyzed by the envfit function and projected on the final NMS solution. This shows the strength (length of arrows) and the direction of the strongest correlation. Additionally, a permutation test of homogeneity between samples (Anderson 2001) was applied to test for effects of treatments on beta-diversity, i.e. differences between aspen and spruce stands concerning the variation in species composition. The NMS, projection of environmental variables and Permdisp test were done with a Bray-Curtis dissimilarity assessments run with 999 permutations.
3 Results
The total basal area per hectare was 3.3 m2 ha–1 and 1.6 m2 ha–1 in the aspen and spruce stands respectively (Table 1). Basal area of aspen was 1.1 m2 ha–1 in the aspen stand, whereas no aspens were found in the spruce stands (Table 3). The aspen stands had higher basal area of all tree species except spruce, and the average number of woody tree species were 6.8 and 4.6 in aspen and spruce stands respectively. The average height was 5.2 and 3.4 m in the aspen and spruce stands. The volumes of coniferous trees in the surroundings were 71 and 100 m3 ha–1 and volumes of deciduous trees were 28 and 24 m3 ha–1 for the aspen and spruce stands respectively. In the aspen stands there were on average 9041, 1672, 277 and 6 stems per ha within the size classes 0–2, 2.1–5, 5.1–10 and > 10.1 cm respectively. The corresponding numbers for spruce were 8423, 775, 72 and 0.
| Table 3. Average tree species basal area (m2 ha–1) in the aspen and spruce stands. | ||
| Aspen stands | Spruce stands | |
| Birch (Betula spp.) | 1.78 | 1.20 |
| Aspen (Populus × wettsteinii) | 1.09 | 0.00 |
| Spruce (Picea abies) | 0.04 | 0.33 |
| Pine (Pinus sylvestris) | 0.02 | 0.01 |
| Willow (Salix spp.) | 0.14 | 0.05 |
| Rowan (Sorbus aucuparia) | 0.07 | 0.01 |
| Alder (Alnus glutinosa) | 0.07 | 0.00 |
| Other broadleaves | 0.06 | 0.00 |
| Total | 3.26 | 1.61 |
In total 18 bird species were recorded, 14 in aspen stands and 12 in spruce stands (Appendix 1). The most commonly encountered bird species were Phylloscopus trochilus (L.) (Willow warbler), Sylvia borin (Boddaert) (Garden Warbler) and Parus major (L.) (Great tit), occurring in 25, 18 and 10 stands respectively. The most common species in aspen stands were Phylloscopus trochilus and Sylvia borin, both occurring in all 13 aspen stands (Fig. 2). Also Prunella modularis (L.) (Dunnock)(7 stands), Emberiza cintronella (L.) (Yellowhammer) (6) and Parus major (6) were common in the aspen stands. Phylloscopus trochilus (12 stands), Emberiza cintronella (6), Sylvia borin (5) and Anthus trivialis (L.) (Tree pipit) (5) were the most common species encountered in spruce stands. Jynx torquilla (L.) (Eurasian Wryneck) was the only red-listed bird encountered (Gärdenfors 2010), with one individual in a spruce stand.
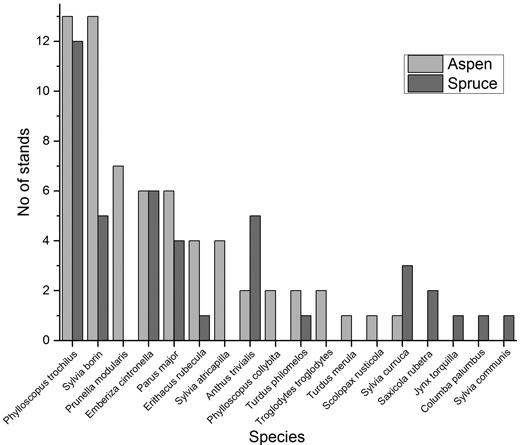
Fig. 2. Number of stands of each stand type (hybrid aspen or spruce regenerations) in which each bird species were encountered.
Aspen stands had significantly higher bird species richness and bird abundance than spruce stands, 4.9 and 8.8 compared to 2.9 and 5.0 respectively (GLM 1 in (Tables 4 and 5, Fig. 3). In order to test the impact on bird diversity of stand characteristics that were unrelated to stand type (aspen or spruce), stems per ha (Stems), retained trees (Ret tree), thinned or not (Thin) and tree evenness (Tree even) (Table 1) were added in GLM 2. This did not improve the model as it caused an increase in AIC in comparison with GLM 1. Adding the interactions between these variables and stand type did not, likewise, significantly improve the model of species richness (GLM 3, Table 4), but it did so for species abundance (GLM 3, Table 5). In addition we found that the effect of stand type on bird abundance was dependent on whether the stands were thinned, with thinning reducing the difference in bird abundance between stand types (GLM 3, Table 5). There was also a positive effect of increasing the number of retained trees in aspen stands on bird species abundance, but as the number of aspen stands with many retained trees was limited, this result should be interpreted with caution.
| Table 4. Generalized linear models (GLM) of species richness in relation to stand type, stand- and landscape variables. For each model the estimated parameters, SE of the parameter estimates as well as t- and p-statistics together with r-square and AIC are presented. See Table 1 for explanation of the variables. Note that the set of variables used in GLM 6 implies multicollinearity, why the individual parameters should be interpreted with care. The overall fit and AIC is, however, comparable with the other GLMs. | |||||||
| α/β | SE | t | p | R2 | AIC | ||
| GLM 1 | Intercept | 1.07 | 0.10 | 10.22 | <0.001 | 0.31 | 95.98 |
| Stand type | 0.52 | 0.15 | 3.51 | 0.002 | |||
| GLM 2 | 0.34 | 102.82 | |||||
| GLM 3 | 0.54 | 101.58 | |||||
| GLM 4 | 0.34 | 104.95 | |||||
| GLM 5 | Intercept | 3.14 | 0.72 | 4.374 | <0.001 | 0.69 | 95.27 |
| Stand type | –2.86 | 0.90 | –3.182 | 0.007 | |||
| Devel | 19.82 | 17.54 | 1.13 | 0.277 | |||
| Agric | –3.51 | 1.78 | –1.969 | 0.069 | |||
| Wetl | –1.90 | 2.80 | –0.68 | 0.508 | |||
| Conif | –0.01 | <0.01 | –2.971 | 0.010 | |||
| Decid | –0.02 | 0.01 | –1.733 | 0.105 | |||
| Stand type: Devel | –34.18 | 27.27 | –1.253 | 0.231 | |||
| Stand type: Agric | 2.20 | 2.21 | 0.997 | 0.336 | |||
| Stand type: Wetl | 1.73 | 5.54 | 0.313 | 0.759 | |||
| Stand type: Conif | 0.02 | 0.01 | 3.649 | 0.003 | |||
| Stand type: Decid | 0.05 | 0.02 | 2.614 | 0.020 | |||
| GLM 6 | Intercept | 0.66 | 0.33 | 1.977 | 0.061 | 0.51 | 92.89 |
| Bas tot | 0.18 | 0.09 | 2.083 | 0.050 | |||
| Bas asp | 0.04 | 0.17 | 0.25 | 0.805 | |||
| Height | –0.13 | 0.12 | –1.133 | 0.270 | |||
| Tree spec | 0.13 | 0.05 | 2.713 | 0.013 | |||
| Table 5. Generalized linear models (GLM) of bird abundance in relation to stand type, stand- and landscape variables. For each model the estimated parameters, SE of the parameter estimates as well as t- and p-statistics together with r-square and AIC. See Table 1 for explanation of the variables. Note that the set of variables used in GLM 6 implies multicollinearity, why the individual parameters should be interpreted with care. The overall fit and AIC is, however, comparable with the other GLMs. | |||||||
| α/β | SE | t | p | R2 | AIC | ||
| GLM 1 | Intercept | 1.61 | 0.13 | 12.73 | <0.001 | 0.26 | 133.74 |
| Stand type | 0.56 | 0.18 | 3.14 | 0.004 | |||
| GLM 2 | Intercept | 1.41 | 0.41 | 3.41 | 0.003 | 0.43 | 134.72 |
| Stand type | 0.41 | 0.19 | 2.14 | 0.045 | |||
| Stems | <0.01 | <0.01 | 1.97 | 0.062 | |||
| Ret tree | –0.01 | 0.02 | –0.49 | 0.628 | |||
| Thin | –0.22 | 0.22 | –0.99 | 0.332 | |||
| Tree even | 0.05 | 0.89 | 0.06 | 0.955 | |||
| GLM 3 | Intercept | 2.08 | 0.55 | 3.77 | 0.002 | 0.61 | 132.67 |
| Stand type | –0.65 | 0.88 | –0.74 | 0.471 | |||
| Stems | <0.01 | <0.01 | 1.90 | 0.076 | |||
| Ret tree | –0.06 | 0.02 | –2.47 | 0.025 | |||
| Thin | 0.63 | 0.35 | 1.78 | 0.094 | |||
| Tree even | –1.59 | 1.21 | –1.32 | 0.207 | |||
| Stand type:Stems | <0.01 | <0.01 | –0.15 | 0.881 | |||
| Stand type:Ret tree | 0.12 | 0.05 | 2.29 | 0.036 | |||
| Stand type:Thin | –1.23 | 0.45 | –2.70 | 0.016 | |||
| Stand type:Tree even | 2.05 | 1.89 | 1.08 | 0.294 | |||
| GLM 4 | 0.38 | 139.28 | |||||
| GLM 5 | 0.56 | 139.74 | |||||
| GLM 6 | Intercept | 0.88 | 0.37 | 2.42 | 0.025 | 0.59 | 123.99 |
| Bas tot | 0.22 | 0.09 | 2.33 | 0.030 | |||
| Bas asp | –0.05 | 0.19 | –0.30 | 0.769 | |||
| Height | 0.03 | 0.13 | 0.22 | 0.827 | |||
| Tree spec | 0.06 | 0.05 | 1.07 | 0.296 | |||
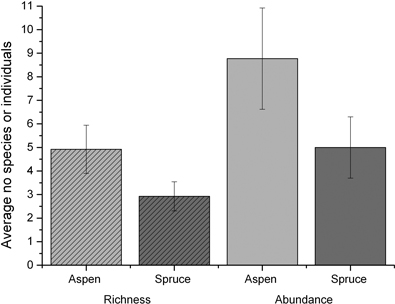
Fig. 3. Average number of bird species (richness) and individuals (abundance) found in hybrid aspen and spruce stands. Error bars show ± two standard errors.
None of the landscape variables had any significant influence on species richness or bird abundance independent of stand type (GLM 4, Tables 4 and 5). However, when the interactions between stand type and the landscape variables were added to the model (GLM 5, Table 4) the model improved as AIC decreased by 9.68 and the R2 doubled for species richness. The volume of coniferous forest in the surrounding landscape (Conif) had a significant impact on the effect of stand type on species richness (GLM 5, Table 4). This effect was a result of significant negative relationship between the predicted bird species richness and the volume of coniferous forest around spruce stands (r = –0.70, p = 0.008), while there was no such negative effect of the coniferous forests surrounding the aspen stands (r = 0.46, p = 0.113). Likewise, the effect of stand type on bird species richness was significantly related to the volume of surrounding deciduous forests (Decid), with a larger difference between the stand types in landscapes with much deciduous trees. In this case, however, the limited number of spruce stands in landscapes with high volume of deciduous trees may have weakened the strength of the correlation (r = 0.38, p = 0.200 for aspen and r = –0.03, p = 0.912 for spruce).
The four variables that showed significantly higher values in aspen than in spruce stands (see Tables 4 and 5): Total basal area (Bas tot), basal area for aspen (Bas asp), stand height (Height) and number of woody species (Tree spec), together explained approximately twice as much of the variation in species richness and bird abundance (GLM 6) as sole stand type (GLM 1). The AICs were also lower for GLM 6 than for GLM 1 for species richness and bird abundance.
The NMS unconstrained ordination resulted in a solution requiring only two-dimensions with a stress factor of 0.15 after four tries, indicating a stable result. The ordination diagram indicates a difference in location in ordination space for aspen and spruce stands, and thus differences in the species composition of their respective bird communities (Fig. 4). This difference was supported by a post hoc fit of environmental variables onto the ordination, according to which stand type correlated significantly with the ordination, although the squared correlation coefficient was rather low (r2 = 0.15; Table 6).
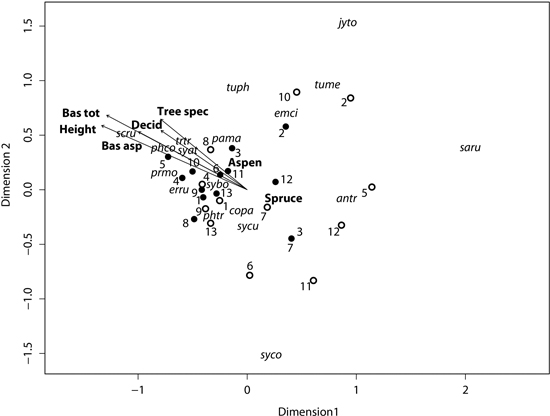
Fig. 4. Ordination diagram from the Non-metric multidimensional scaling showing the two-dimensional final solution with sites (dots) and species. The significant (p ≤ 0.05) environmental variables (names in bold) from a posthoc fit is shown as arrows (continuous) or location in ordination space (factors). Closed dots show the location of aspen sites, while open dots indicate spruce sites. The sites of a spruce/aspen pair have the same number. Phtr is Phylloscopus trochilus, sybo Sylvia borin, prmo Prunella modularis, emci Emberiza citrinella, pama Parus major, erru Erithacus rubecula, syat Sylvia atricapilla, antr Anthus trivialis, phco Phylloscopus collybita, tuph Turdus philomelos, trtr Troglodytes troglodytes, tume Turdus merula, scru Scolopax rusticola, sycu Sylvia curruca, saru Saxicola rubetra, jyto Jynx torquilla, copa Columba palumbus, syco Sylvia communis. A description of the environmental variables can be found in Table 1.
On the other hand, we found that total basal area (Bas tot), aspen basal area (Bas asp), stand height (Height), number of tree species (Tree spec) and volume deciduous trees in the landscape (Decid) separately were more important for the bird community composition than stand type. These variables showed r-squares between r2 = 0.23 and 0.53, with basal area and tree height as the most informative (Table 6). All significant variables were negatively and most strongly correlated with the first NMS dimension.
| Table 6. Posthoc fit of environmental variables on the two-dimensional solution from the Non-metric multidimensional scaling. The table shows the independent correlations (continuous) with the two dimensions separately, or location of the centroid (factors) in ordination space together with r-square and p-value for each variable in relation to both dimensions. | ||||
| NMDS1 corr/centr | NMDS2 corr/centr | r2 | P | |
| Bas tot | –0.88 | 0.47 | 0.53 | 0.001 |
| Stems | –0.95 | 0.32 | 0.21 | 0.072 |
| Bas asp | –0.88 | 0.47 | 0.32 | 0.010 |
| Height | –0.91 | 0.40 | 0.53 | 0.002 |
| Ret tree | 0.43 | –0.90 | 0.00 | 0.986 |
| Tree spec | –0.78 | 0.63 | 0.26 | 0.032 |
| Tree even | 0.00 | 1.00 | 0.03 | 0.690 |
| Devel | –0.38 | 0.92 | 0.11 | 0.260 |
| Agric | –0.72 | 0.69 | 0.06 | 0.512 |
| Wetl | –0.86 | –0.51 | 0.09 | 0.339 |
| Conif | 0.80 | –0.61 | 0.16 | 0.134 |
| Decid | –0.82 | 0.57 | 0.23 | 0.043 |
| Spruce stand | 0.21 | –0.13 | 0.15 | 0.013 |
| Aspen stand | –0.21 | 0.13 | ||
| Unthin | –0.09 | 0.00 | 0.05 | 0.315 |
| Thin | 0.24 | –0.00 | ||
The species Scolopax rusticola (L.) (Eurasian Woodcock), Phylloscopus collybita (Vieillot) (Chiffchaff), Prunella modularis, Sylvia atricapilla (L.) (Blackcap) and Troglodytes troglodytes (L.) (Eurasian Wren) were most indicative of aspen stands while Jynx torquilla, Turdus merula (L.) (Blackbird), Anthus trivialis, Saxicola rubetra (L.) (Whinchat) and Sylvia communis (L.) (Common whitethroat) are closer to spruce stands in the ordination space. The ordination indicates a larger dispersion (i.e. β-diversity) of spruce stand bird communities than that which occurred for aspen stands. Accordingly, the Permdisp test showed a significant difference in distance to group centroids (p = 0.011) of 0.45 and 0.31 for spruce and aspen stands respectively.
4 Discussion
4.1 Bird species richness and abundance
As in Swedish production forests in general (Felton et al. 2011; Johansson et al. 2013), the bird diversity in both stand types is primarily composed of relatively common species. Nevertheless, the hybrid aspen stands had higher bird species richness and abundance as well as a distinct community composition compared to the spruce stands. These results appear to be influenced by factors which are coupled with decisions regarding stand development as a hybrid aspen or spruce stand. We suggest this because the GLMs explained significantly more variation in bird species richness and abundance when four stand variables were added to the models. The four variables – total basal area, aspen basal area, stand height and number of woody species – were highly correlated and found to be significantly higher in the aspen stands. Hence, the higher levels of richness and abundance of birds were associated with stands that can be described as being more structurally complex, at least with respect to tree species composition, tree species richness, and stand height (which allows for greater vertical stratification); factors shown to be important determinants of bird diversity (Hinsley et al. 2009; Hewson et al. 2011). One reason is probably that the diverse tree species composition and high tree species richness has a positive influence on the diversity and amount of insects (Kennedy and Southwood 1984), and relatedly, several of the bird species found to be more common in the aspen stands are predominantly insectivores during breeding season, e.g. Sylvia atricapilla, Sylvia borin and Prunella modularis (BWPi 2007). The importance of structural complexity as an underlying driver of observed differences in bird communities in our study is also supported by the impact that thinning had on reducing the difference in bird abundance between the stand types.
Previous studies have indicated that bird communities (and mammals, and some invertebrate groups) are often less species rich in short rotation forestry stands than in natural or semi-natural forests (Hanowski et al. 1997; Christian et al. 1998; Dhondt and Wrege 2003; Schulz et al. 2009). A meta-analysis recently conducted in the United States found that Populus based short rotation forestry stands generally harbored lower levels of avian diversity than reference woodlands, but none of the studies directly compared short rotations stands with forest plantations (Riffell et al. 2011). Likewise, comparisons of the biodiversity in short rotation production forests with broadleaves and more conventional conifer dominated production forests are largely lacking in Northern Europe. Our study thereby helps to address this gap in the ecological literature.
As the study involves two different stand types, it was possible that resultant differences in species richness and abundance arose because of the influence of associated differences in vegetation densities on the frequency of bird detections during surveys. However, as in this case the stands with the highest bird species richness and abundance were also the stands with the densest and highest vegetation, this potential concern can only serve to support the validity our findings, rather than be a challenge to them.
There was also an apparent positive effect of increasing the number of retained trees in aspen stands on bird species abundance. As the study was not explicitly designed to tease out their potential benefit to bird communities, the statistical results were inconclusive and should be interpreted with caution. Nevertheless, we made repeated observations of individuals using retained trees in both spruce and aspen stands, which is indicative of at least some minimal level of contribution as a resource. This perspective is further justified by other studies of green tree retention in Northern Europe which have shown their importance for the bird fauna (Rosenvald and Löhmus 2007; Söderström 2009)
4.2 Landscape effects
Species richness was affected by an interaction between the total volume of coniferous forest in the landscape and stand type, with a negative correlation between coniferous forests and species richness in spruce stands. This was counterintuitive, as young spruce stands in landscapes with high volumes of coniferous trees could have been predicted to harbour more birds due to a “landscape complementation” effect (Dunning et al. 1992; Tubelis et al. 2004), i.e. that birds using the older stands could have found complementary resources in the nearby young stands. Likewise, there could have been a “spillover effect” from older spruce stands in the surrounding landscape, increasing the occupancy of the younger stands (Hewson et al. 2011). Production conifer stands dominate in our study area, and low volumes of coniferous forests is indicative of low forest volume in general. Because of the Gudrun hurricane in 2005 the area today has relatively much young forest with low forest cover. The higher richness in stands located in areas with less coniferous forests, may therefore have arisen because young spruce stands attracted a number of bird species associated with open habitats, in addition to common forest species. This interpretation is supported by the occurrence of Saxicola rubetra, Jynx torquilla and Sylvia communis in spruce stands only (Fig. 2), i.e. species commonly associated with open habitats (Heath et al. 2000; Ottosson et al. 2012).
4.3 Species composition and habitat requirements
The aspen stands are more tightly aggregated in ordination space relative to the spruce stands, indicating that aspen stands are more consistent in terms of the bird species compositions they support. This may be related to the greater variability in tree height and tree density in the spruce stands compared with aspen. In addition, these outcomes may be related to the vegetation structure in the surroundings, because in the outskirts of the ordination space, and associated with the spruce stands, are the species representative of open landscapes (e.g. Jynx torquilla, Saxicola rubetra and Sylvia communis) (BWPi 2007). Correspondingly, several bird species that were encountered exclusively or significantly more often in the aspen stands also seem to have habitat requirements that are consistent with their observed occurrence in the two stand types assessed. Sylvia atricapilla was recorded exclusively in aspen stands (Fig. 2). This species is highly arboreal and is primarily associated with deciduous dominated woodlands possessing a tall undergrowth (BWPi 2007). These habitat requirements were better associated with the aspen stands, than with the spruce stands assessed. Prunella modularis and Sylvia borin, which often are associated with coniferous woods and shrubs (BWPi 2007), were also found almost exclusively in aspen stands. The absence of such an association in our study may result from the low and open nature of the very young spruce regeneration stands assessed, versus the relatively dense and “shrubby” vegetation found in many of the aspen stands. Prunella modularis is a species with a decreasing population trend in recent decades (Lindström and Green 2012), and perhaps an increased number of aspen stands in the landscape could mitigate these losses.
4.4 Management and policy implications
Both aspen and spruce were only the second most common tree in their respective stand type. Most stands were dominated by birch, which is the case for all young plantations in the region before thinning regardless of focal tree species. However, presumably due to associated differences in treatment (e.g. fencing versus no-fencing), more birch was encountered in the aspen stands. In southern Sweden large herbivores, such as moose (Alces alces (L.)) and roe deer (Capreolus capreolus (L.)) currently occur at high densities, with substantial levels of browsing damages to forests that lack fencing (Ingemarsson et al. 2007). Multiple studies have demonstrated that the exclusion of browsers can significantly alter the vegetation and the associated biodiversity in Sweden (Mathisen et al. 2010; Edenius et al. 2011; Vesterlund et al. 2012). Hence, whether stands are fenced or not, may contribute to the observed differences in bird diversity and species composition between the stand types.
The young stands (4–8 years in our study) represent a large part of overall rotation length in hybrid aspen stands (1/3 in some cases), and is thus of interest to conservation efforts. Bird species turnover is often high during stand development (Wells et al. 2011), and the bird community will probably change over coming years as stands are thinned and steered towards relatively pure aspen or spruce vegetation. Because of this it will be important to determine the extent to which the longer rotation period of spruce stands may facilitate a greater temporal variation in bird community composition and diversity than can be achieved in aspen stands. While the benefits of a longer rotation time in spruce stands is worthy of consideration, the bird diversity supported throughout is perhaps unlikely to override the landscape benefits of planting deciduous based stands (Felton et al. 2011), like hybrid aspen, in regions disproportionately dominated by production conifers.
5 Conclusions
Our results indicate that, during early stages of regeneration, fenced aspen stands can support relatively high levels of bird diversity in comparison to the dominating forest type in the region. These results need to be evaluated in light of hybrid aspen’s contribution to climate change mitigation, and ecological concerns (Felton et al. 2013) when deciding upon its suitability as an alternative forest land-use.
Acknowledgements
The study was funded by the research programme “Broadleaves for the future” at SLU. AF was funded in part by Future Forests, a multi-disciplinary research programme supported by the Foundation for Strategic Environmental Research (MISTRA). We are grateful to Emma Holmström for assistance with the landscape variable analysis.
The article is based on a presentation given at the Workshop on “Climate Change and Forestry in Northern Europe” held in Uppsala, Sweden, 11–12 November, 2013. The workshop was organized by Future Forests at the Swedish University of Agricultural Sciences (SLU), EFI-Nord, and the Finnish Forest Research Institute (Metla).
References
Akaike H. (1973). Information theory as an extension of the maximum likelihood principle. In: Petrov B.N., Csaki F. (eds.). Proc. Second International Symposium on Information Theory. Akademiai Kaido, Budapest. p. 267–281.
Alldredge M.W., Simons T.R., Pollock K.H. (2007). A field evaluation of distance measurement error in auditory avian point count surveys. Journal of Wildlife Management 71: 2759–2766. http://dx.doi.org/10.2193/2006-161.
Anderson M.J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecology 26: 32–46. http://dx.doi.org/10.1111/j.1442-9993.2001.01070.pp.x.
Baum S., Bolte A., Weih M. (2012). High value of short rotation coppice plantations for phytodiversity in rural landscapes. Global Change Biology Bioenergy 4: 728–738. http://dx.doi.org/10.1111/j.1757-1707.2012.01162.x.
Berg A. (2002). Breeding birds in short-rotation coppices on farmland in central Sweden – the importance of Salix height and adjacent habitats. Agriculture Ecosystems & Environment 90: 265–276. http://dx.doi.org/10.1016/S0167-8809(01)00212-2.
Bergquist J., Löf M., Örlander G. (2009). Effects of roe deer browsing and site preparation on performance of planted broadleaved and conifer seedlings when using temporary fences. Scandinavian Journal of Forest Research 24: 308–317. http://dx.doi.org/10.1080/02827580903117420.
Bibby C.J., Aston N., Bellamy P.E. (1989). Effects of broadleaved trees on birds of upland conifer plantations in North Wales. Biological Conservation 49: 17–29. http://dx.doi.org//0006-3207(89)90110-9.
Bibby C.J., Burgess N.D., Hill D.A. (2000). Bird census techniques. London, Academic Pres.
Bonthoux S., Balent G. (2012). Point count duration: five minutes are usually sufficient to model the distribution of bird species and to study the structure of communities for a French landscape. Journal of Ornithology 153: 491–504. http://dx.doi.org/10.1007/S10336-011-0766-2.
BWPi (2007). Birds of the Western Palearctic interactive. Bird guides. Oxford University Press.
Christian D.P., Hoffman W., Hanowski J.M., Niemi G.J., Beyea J. (1998). Bird and mammal diversity on woody biomass plantations in North America. Biomass & Bioenergy 14: 395–402. http://dx.doi.org/10.1016/S0961-534(97)10076-9.
Dhondt A.A., Wrege P.H. (2003). Avian biodiversity studies in short-rotation woody crops. Cornell University Laboratory of Ornithology, Ithaca.
Dunning J.B., Danielson B.J., Pulliam H.R. (1992). Ecological processes that affect populations in complex landscapes. Oikos 65: 169–175. http://dx.doi.org/10.2307/3544901.
Edenius L., Ericsson G., Kempe G., Bergstrom R., Danell K. (2011). The effects of changing land use and browsing on aspen abundance and regeneration: a 50-year perspective from Sweden. Journal of Applied Ecology 48: 301–309. http://dx.doi.org/10.1111/J.1365-2664.2010.01923.X.
Farmer R.G., Leonard M.L., Horn A.G. (2012). Observer effects and avian-call-count survey quality: rare-species biases and overconfidence. Auk 129: 76–86. http://dx.doi.org/10.1525/Auk.2012.11129.
Felton A., Ellingson L., Andersson E., Drossler L., Blennow K. (2010). Adapting production forests in southern Sweden to climate change constraints and opportunities for risk spreading. International Journal of Climate Change Strategies and Management 2: 84–97. http://dx.doi.org/10.1108/17568691011020274.
Felton A., Andersson E., Ventorp D., Lindbladh M. (2011). A comparison of avian diversity in spruce monocultures and spruce-birch polycultures in southern Sweden. Silva Fennica 45(5): 1143–1150. http://dx.doi.org/10.14214/sf.92.
Felton A., Boberg J., Björkman C., Widenfalk O. (2013). Identifying and managing the ecological risks of using introduced tree species in Sweden’s production forestry. Forest Ecology and Management 307: 165–177. http://dx.doi.org/10.1016/j.foreco.2013.06.059.
Gärdenfors U., Ed. (2010). The 2010 Red List of Swedish species. SLU, ArtDatabanken, Uppsala.
Gustafsson L., Felton A., Felton A.M., Brunet J., Caruso A., Hjältén J., Lindbladh M., Ranius T., Roberge J.-M., Weslien J. (2014). Natural versus national boundaries: the importance of considering biogeographical patterns in forest conservation policy. Conservation Letters. http://dx.doi.org/10.1111/conl.12087.
Hanowski J.M., Niemi G.J., Christian D.C. (1997). Influence of within-plantation heterogeneity and surrounding landscape composition on avian communities in hybrid poplar plantations. Conservation Biology 11: 936–944. http://dx.doi.org/10.1046/j.1523-1739.1997.96173.x.
Heath M.F., Borggreve C., Peet N., Hagemeijer W.J.M. (2000). European bird populations: estimates and trends. Cambridge U.K., BirdLife International.
Hewson C.M., Austin G.E., Gough S.J., Fuller R.J. (2011). Species-specific responses of woodland birds to stand-level habitat characteristics: the dual importance of forest structure and floristics. Forest Ecology and Management 261: 1224–1240. http://dx.doi.org/10.1016/j.foreco.2011.01.001.
Hinsley S.A., Hill R.A., Fuller R.J., Bellamy P.E., Rothery P. (2009). Bird species distributions across woodland canopy structure gradients. Community Ecology 10: 99–110. http://dx.doi.org/10.1556/ComEc.10.2009.1.12.
Ingemarsson F., Claesson S., Thuresson T. (2007). Älg- och rådjursstammarnas kostnader och värden. [Costs and values of the moose and roe deer populations]. Skogsstyrelsen, Rapport 3. [In Swedish].
Johansson T., Hjältén J., de Jong J., von Stedingk H. (2013). Environmental considerations from legislation and certification in managed forest stands: a review of their importance for biodiversity. Forest Ecology and Management 303: 98–112. http://dx.doi.org/10.1016/J.Foreco.2013.04.012.
Johnson D.H. (2008). In defense of indices: the case of bird surveys. Journal of Wildlife Management 72: 857–868. http://dx.doi.org/10.2193/2007-294.
Kennedy C.E.J., Southwood T.R.E. (1984). The number of species of insects associated with British trees: a re-analysis. Journal of Animal Ecology 53: 455–478. http://dx.doi.org/10.2307/4528.
Lindenmayer D.B., Hulvey K.B., Hobbs R.J., Colyvan M., Felton A., Possingham H., Steffen W., Wilson K., Youngentob K., Gibbons P. (2012). Avoiding bio-perversity from carbon sequestration solutions. Conservation Letters 5: 28–36. http://dx.doi.org/10.1111/j.1755-263X.2011.00213.x.
Lindström Å., Green M. (2012). Övervakning av fåglarnas populationsutveckling. Årsrapport för 2012. Biologiska institutionen, Lunds Universitet. [In Swedish].
Löf M., Bergquist J., Brunet J., Karlsson M., Welander T. (2010). Conversion of Norway spruce stands to broadleaved woodland – regeneration systems, fencing and performance of planted seedlings. Ecological Bulletins 53: 165–174.
Mathisen K.M., Buhtz F., Danell K., Bergstrom R., Skarpe C., Suominen O., Persson I.L. (2010). Moose density and habitat productivity affects reproduction, growth and species composition in field layer vegetation. Journal of Vegetation Science 21: 705–716. http://dx.doi.org/10.1111/j.1654-1103.2010.01180.x.
Oksanen J., Blanchet F.G., Kindt R., Legendre P., Minchin P.R., O’Hara R.B., Simpson G.L., Solymos P. M.H.H. S., Wagner H. (2013). Vegan: community ecology package. R package version 2.0-8. http://vegan.r-forge.r-project.org/. [Cited 25 August 2014].
Ottosson U., Ottvall R., Elmberg J., Green M., Gustafsson R., Haas F., Holmqvist N., Lindström Å., Nilsson L., Svensson M., Svensson S., Tjernberg M. (2012). Fåglarna i Sverige – antal och förekomst. Halmstad, SOF. [In Swedish].
Peck K.M. (1989). Tree species preferences shown by foraging birds in forest plantation in northern England. Biological Conservation 48: 41–57. http://dx.doi.org/10.1016/0006-3207(89)90058-x.
R Core Team (2013). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. [Cited 25 August 2014].
Reese H., Nilsson M., Pahlen T.G., Hagner O., Joyce S., Tingelof U., Egberth M., Olsson H. (2003). Countrywide estimates of forest variables using satellite data and field data from the national forest inventory. Ambio 32: 542–548. http://dx.doi.org/10.1579/0044-7447-32.8.542.
Riffell S.A.M., Verschuyl J., Miller D., Wigley T.B. (2011). A meta-analysis of bird and mammal response to short-rotation woody crops. GCB Bioenergy 3: 313–321. http://dx.doi.org/10.1111/j.1757-1707.2010.01089.x.
Roberge J.M., Angelstam P. (2006). Indicator species among resident forest birds – a cross-regional evaluation in northern Europe. Biological Conservation 130: 134–147. http://dx.doi.org/10.1016/J.Biocon.2005.12.008.
Rosenvald R., Löhmus A. (2007). Breeding birds in hemiboreal clear-cuts: tree-retention effects in relation to site type. Forestry 80: 502–515. http://dx.doi.org/10.1093/forestry/cpm027.
Rytter L., Stener L.G. (2011). Odling av hybridasp och poppel: en handledning från Skogforsk. Skogforsk, Uppsala. [In Swedish].
Sage R.B. (1998). Short rotation coppice for energy: towards ecological guidelines. Biomass & Bioenergy 15: 39–47. http://dx.doi.org/10.1016/S0961-9534(97)10055-1.
Schulz U., Brauner O., Gruss H. (2009). Animal diversity on short-rotation coppices – a review. Landbauforschung Volkenrode 59: 171–181.
Söderström B. (2009). Effects of different levels of green- and dead-tree retention on hemi-boreal forest bird communities in Sweden. Forest Ecology and Management 257: 215–222. http://dx.doi.org/10.1016/j.foreco.2008.08.030.
Sweeney O.F.M.C.D., Wilson M.W., Irwin S., Kelly T.C., O’Halloran J. (2010). The influence of a native tree species mix component on bird communities in non-native coniferous plantations in Ireland. Bird Study 57: 483–494. http://dx.doi.org/10.1080/00063657.2010.489937.
Toms J.D., Schmiegelow F.K.A., Hannon S.J., Villard M.A. (2006). Are point counts of boreal songbirds reliable proxies for more intensive abundance estimators? Auk 123: 438–454. http://dx.doi.org/10.1642/0004-8038(2006)123%5B438:APCOBS%5D2.0.CO;2.
Tubelis D.P., Lindenmayer D.B., Cowling A. (2004). Novel patch-matrix interactions: patch width influences matrix use by birds. Oikos 107: 634–644. http://dx.doi.org/10.1111/j.0030-1299.2004.13420.x.
Tullus A., Rytter L., Tullus T., Weih M., Tullus H. (2012). Short-rotation forestry with hybrid aspen (Populus tremula L. * P. tremuloides Michx.) in Northern Europe. Scandinavian Journal of Forest Research 27: 10–29. http://dx.doi.org/10.1080/02827581.2011.628949.
Tullus A., Tullus H., Vares A., Kanal A. (2007). Early growth of hybrid aspen (Populus × wettsteinii Hämet-Ahti) plantations on former agricultural lands in Estonia. Forest Ecology and Management 245: 118–129. http://dx.doi.org/10.1016/j.foreco.2007.04.006.
Valinger E., Fridman J. (2011). Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. Forest Ecology and Management 262: 398–403. http://dx.doi.org/10.1016/j.foreco.2011.04.004.
Vesterlund S.R., Suominen O., Bergstrom R., Danell K., Persson I.L. (2012). The impact of simulated moose densities on conifer aphids along a productivity gradient. Ecography 35: 105–112. http://dx.doi.org/10.1111/j.1600-0587.2011.06534.x.
Weih M., Karacic A., Munkert H., Verwijst T., Diekmann M. (2003). Influence of young poplar stands on floristic diversity in agricultural landscapes (Sweden). Basic and Applied Ecology 4: 149–156. http://dx.doi.org/10.1078/1439-1791-00157.
Wells K., Bohm S.M., Boch S., Fischer M., Kalko E.K.V. (2011). Local and landscape-scale forest attributes differ in their impact on bird assemblages across years in forest production landscapes. Basic and Applied Ecology 12: 97–106. http://dx.doi.org/10.1016/j.baae.2011.01.002.
Total of 53 references
