Timo Domisch  ,
Saija Huuskonen,
Juho Matala,
Ari Nikula
,
Saija Huuskonen,
Juho Matala,
Ari Nikula
Interactive effects of moose browsing and stand composition on the development of mixed species seedling stands
Domisch T., Huuskonen S., Matala J., Nikula A. (2024). Interactive effects of moose browsing and stand composition on the development of mixed species seedling stands. Silva Fennica vol. 58 no. 4 article id 23077. https://doi.org/10.14214/sf.23077
Highlights
- We conducted meta-analyses assessing effect sizes of moose exclusion on seedling density and height, as well as regressions between stand density and deciduous seedling proportion and effect sizes
- The more deciduous trees in a seedling stand, the smaller was the fencing effect, but at some point, the conifer seedling growth was impaired by too many deciduous seedlings. At the same time, the protective fencing effect increased with stand density
- An appropriate deciduous admixture in conifer-dominated mixed seedling stands seems to improve moose damage tolerance.
Abstract
The moose (Alces alces L.), a common large herbivore in the boreal region, impairs forest regeneration by browsing on tree seedlings and saplings. Moose prefer deciduous species, but during winter more coniferous seedlings are used. We used meta-analyses, separately for deciduous and coniferous seedlings, for evaluating whether excluding moose browsing affected seedling density and height. In addition, we compared (1) deciduous seedling proportion, (2) stand density, (3) elapsed time from fencing and (4) estimated moose density with moose exclusion effect sizes. Fencing had a positive effect on coniferous seedling height. With more deciduous trees in a seedling stand, the fencing effect for both seedling height and density of coniferous seedlings decreased. On the other hand, the fencing effects increased with denser stands. At some point effect sizes turned to negative, and conifer species varied in their response to browsing. This implies that deciduous seedlings can protect conifers from browsing by moose up to some mixing ratio, but when deciduous seedling densities are too high, their negative effect increases, presumably through increased competition. Our results suggest that a moderate deciduous admixture in conifer-dominated mixed seedling stands can decrease moose damage but also underline the significance of timely silvicultural measures to minimize the negative effects of excessive deciduous seedlings and too dense stands. Due to differences in coniferous and deciduous species, as well as their compositions and amounts in studied experiments, more studies adjusted to local conditions are still needed to give exact measures for silvicultural recommendations.
Keywords
boreal forest;
tree species;
Alces alces;
ungulates;
moose damage;
mixed stands;
meta-analysis
-
Domisch,
Natural Resources Institute Finland (Luke), Natural Resources, Yliopistokatu 6B, FI-80100 Joensuu, Finland
 https://orcid.org/0000-0001-7026-1087
E-mail
timo.domisch@luke.fi
https://orcid.org/0000-0001-7026-1087
E-mail
timo.domisch@luke.fi
- Huuskonen, Natural Resources Institute Finland (Luke), Natural Resources, Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail saija.huuskonen@luke.fi
-
Matala,
Natural Resources Institute Finland (Luke), Natural Resources, Yliopistokatu 6B, FI-80100 Joensuu, Finland
https://orcid.org/0000-0002-5867-5057
E-mail
juho.matala@luke.fi
-
Nikula,
Natural Resources Institute Finland (Luke), Natural Resources, Ounasjoentie 6, FI-96200 Rovaniemi, Finland
https://orcid.org/0000-0001-8372-8440
E-mail
ari.nikula@luke.fi
Received 18 December 2023 Accepted 12 September 2024 Published 16 September 2024
Views 69572
Available at https://doi.org/10.14214/sf.23077 | Download PDF

Supplementary Files
1 Introduction
The moose (Alces alces L.) is a common large herbivore in the boreal region (Jensen et al. 2020; Niedziałkowska et al. 2022). As a game species, it has economic value (Mattson 1990), but it also significantly affects forest ecosystems and forestry by selective browsing which can affect tree species composition and causes economic damage to forest owners (Edenius et al. 2002; Nevalainen et al. 2016). The moose impact on forests is dependent on moose density (Nikula et al. 2021), but also other factors like regional forest structure (Nikula et al. 2021), competition with other cervids (Pfeffer et al. 2021; Spitzer et al. 2021) and stand characteristics (Milligan and Koricheva 2013) determine how the effects of browsing are realized.
Moose benefit from large forest regeneration areas of monocultural and even-aged Scots pine (Pinus sylvestris L.) seedling stands due to clear-cut based forest regeneration methods that have been applied in the Nordic countries since the 1950s and 1960s (Markgren 1974; Lavsund 1987). Resulting from that, abandonment of rural areas, and adopted hunting practises, moose populations increased rapidly from the early 1970s until the 1980s (Cederlund and Markgren 1987; Nygrén 1987; Østgård 1987; Lavsund et al. 2003). High moose populations have caused extensive damage to forests in Finland and other Nordic countries. In Finland, forest damage by moose and other cervids was recorded on 555 000 ha of forest land during 2014–2018 (Korhonen et al. 2021). Pine dominated stands comprised 75% of the total damage area, and the proportion of damaged stands raged between 14 to 29.8% of all pine stands (Nevalainen et al. 2016; Korhonen et al. 2021). Similar numbers were estimated for Southern Sweden, where recent assessments concluded that moose damage occurred on 12–20% of young pine stems (SFA 2018).
Moose show a seasonally changing use and forage preference of different plant species (Moen et al. 1997; Faber and Lavsund 1999), and the most obvious change in preference for different tree species can be observed between winter and summer (Spitzer et al. 2020). When the snow cover exceeds 30 cm, the diet consists almost entirely of trees and shrubs (Cederlund et al. 1980). Deciduous tree species, such as rowan (Sorbus aucuparia L.), willows (Salix spp.) and aspen (Populus tremula L.), are highly preferred (Bergström and Hjeljord 1987; Edenius 1991) whereas Scots pine is only of median species in the preference list of moose (Månsson et al. 2007). In contrast to the summer diet, which consists mostly of deciduous trees and herbs, such as birch (Betula spp.) or fireweed (Chamaenerion angustifolium (L.) Scop.), the great majority of the food during winter constitutes of Scots pine (Cederlund et al. 1980). As a consequence, economic loss to forestry occur mainly in winter (Cederlund et al. 1980; Gill 1992). However, also the summer browsing can result in a considerable economic loss (Bergqvist et al. 2012).
Milligan and Koricheva (2013), assessing effects of tree species richness on moose browsing in experimental young plantations in Western Finland, found that browsing intensity increased with tree species richness while browsing selectivity of tree species decreased, and was more evenly distributed in species-rich mixtures. In addition, also the tree species composition was an important determinant of browsing intensity. The occurrence of preferred species together (pine and birch) resulted in the greatest browsing intensity while intermediately preferred species (Siberian larch: Larix sibirica Ledeb. and black alder: Alnus glutinosa (L.) Gaertn.) experienced associational susceptibility when growing with pine and birch. In contrast, Milligan and Koricheva (2013) did not find evidence for associational resistance, indicated by the presence of a least preferred species (Norway spruce: Picea abies (L.) H. Karst.) in a mixture having no significant effect on moose browsing intensity on other tree species.
The deciduous admixture, that appears usually by natural regeneration in pine seedling stands, has been reported to have ambivalent effects on moose damage of pine seedlings. First, the deciduous admixture has been reported to increase the browsing of Scots pine and overall browsing intensity (Härkönen et al. 2008; Milligan and Koricheva 2013). Furthermore, deciduous species can hamper the development of conifer seedlings by their faster development at the initial succession phase (Bergqvist et al. 2014). On the other hand, deciduous seedlings have been reported to protect coniferous seedlings by being the more preferred species for moose (Heikkilä 1991). In addition, some studies report no effect of deciduous mixture on Scots pine browsing (Bergqvist et al. 2011).
For several tree species (e.g., Scots pine seedlings or saplings), browsing intensity has been demonstrated to correlate with foraging availability (Bergström et al. 1995; Hörnberg 2001). Månsson et al. (2007) determined moose selectivity indices for various tree species in Southern Sweden. They concluded that rowan, willow, and aspen had a much higher probability of being browsed than Scots pine and downy birch (Betula pubescens Ehrh.), with juniper (Juniperus communis L.) and silver birch (Betula pendula Roth) ranging in between. Moose always preferred silver birch over downy birch, especially in the height class from 1.5 to 3 m (Danell and Ericson 1986). Due to the high moose population in their study area in Southern Finland (3.6–10.9 per 1000 ha), de Chantal et al. (2009) concluded that the successful recruitment of mature stems of aspen, willow and rowan in the landscape was uncertain. Due to its higher density the situation for birch was much better. Danell et al. (1985) observed that after moose browsing, birches produced larger shoots, and shoots on browsed trees had a longer growing season than on unbrowsed trees. Bergqvist et al. (2014) observed that in Central Sweden, pine damage was dependent on moose density and pine cover, birch damage only on moose density, while aspen damage was dependent on birch and pine cover. Nevalainen et al. (2016), analysing data from national forest inventory data in Finland, concluded, that damage decreases in Scots pine-dominated saplings with increasing pine proportion in a stand but increases with increasing birch or spruce proportion in a stand.
Basically, there are three alternatives for reducing moose damage: (1) actively reducing moose densities, (2) increasing amount of food, or (3) decreasing food availability (Angelstam et al. 2000). Bergqvist et al. (2014) concluded that increasing stand density might be a more efficient way to reduce moose damage than reducing moose populations. De Vriendt et al. (2020) brought up the idea that large herbivores could be used as cost-effective means for controlling stand density, particularly of competing species, when combining hunting and forest management. Increasing the number of Scots pine trees in young stands has also been suggested to decrease moose damage at a given moose population density (Lyly and Saksa 1992; Ball and Dahlgren 2002). Andrén and Angelstam (1993) suggested that stand density can be managed by silvicultural measures, which is important in that moose browsing was reduced when the density of young trees exceeded about 1500 to 2500 per ha, depending on the geographical location. In comparison, Lyly and Saksa (1992) evaluated that at least 4000 pine seedlings in regeneration would be needed to increase stand tolerance to moose browsing.
As one target of future forest management is to increase the proportion of mixed forests (Felton et al. 2016; Huuskonen et al. 2021), one of the most challenging issues is how forest management and silvicultural measures can decrease the moose damage risk in mixed seedling and sapling stands. This is of particular importance in conifer dominated seedling or sapling stands with an existing admixture of deciduous tree species. One important question would be how forest management could cope with moose browsing, so that the main target trees species remain protected, and damage is directed to other than the main target tree species. Related to this, another point would concern the optimum conifer density and admixture of deciduous seedlings, which would decrease pine damage but still not affect its growth.
Our main aim was to estimate the moose exclusion effect in conifer dominated mixed seedling and sapling stands. Another aim was to find out how other factors, such as deciduous seedling density, moose density or the time elapsed from fencing, would affect the observed effect sizes. We hypothesise, (1) that forest management and appropriate silvicultural measures can decrease moose damage risk in conifer dominated mixed seedling and sapling stands, and (2) that factors increasing moose damage risk and related to spatial conditions and stand structure can be identified. Accordingly, the tree species composition, species richness and the amount of available food in plantations affect the browsing selection by moose and thus tree damage.
2 Materials and methods
First, we conducted meta-analyses (1) to estimate an overall moose exclusion effect in boreal seedling or sapling stands and to assess whether it is significantly different from zero, separately for deciduous and coniferous seedlings, and (2) to identify and characterise factors that could have an impact on the between-study heterogeneity and the overall effect estimate. Then we conducted regression analyses to find out how deciduous seedling proportion, total stand density, moose density and the time elapsed from fencing, affected the observed effect sizes.
We used Web of Science, Google Scholar, ScienceDirect and Scopus as databases for searching potential articles, and the time of searching was first November 2020 and then again June 2021. The temporal extent of publication was not restricted. The following keywords and different combinations of them were used: ’moose’, ‘herbivore’, ‘damage’, ‘shoot’, ‘boreal or hemiboreal’, ‘density’, ‘height’, ‘fenc*’, ‘excl*’, ‘browsing’, ‘conifer or deciduous or broadleaf’ and ‘pine or spruce or birch’. Additional research papers that were referenced within or cited by these papers were added to achieve a comprehensive database of moose exclusion effects on the density and height on boreal and hemiboreal tree species.
Large herbivore (here moose) exclusion, i.e., fencing, was one important criterion for selecting a study. Another one was, that the density or height of deciduous and coniferous seedlings were reported as response variables both for fenced moose exclosures and open controls. We concentrated on studies conducted under boreal or hemiboreal conditions, and where moose was the main affecting herbivore. Data were compiled from article text and tables directly or extracted from figures using WebPlotDigitizer version 4.4 (Rohatgi 2021).
From all 250 potential studies we selected only those that reported averages and standard errors or standard deviation for the treatment (herbivore exclusion = fencing) and the respective control. This yielded in a total of 14 studies (Table 1) which included 76 observations for seedling density and 46 observations for seedling height.
| Table 1. List of publications and studied tree species used for the meta-analyses, assessing effect sizes of moose exclusion on seedling density and height. Abbreviations for tree genera or species refer to Table 2. | ||||
| Author(s) | Year | Tree genus or species used in the analyses | Location | |
| 1 | Angell & Kielland | 2009 | SALIX | Alaska, USA |
| 2 | Faison et al. | 2016 | DECID | Massachusetts, USA |
| 3 | Heikkilä et al. | 2003 | BETULA, PICEA, PINSYL, SALIX, SORBUS | C Finland |
| 4 | Heikkilä & Tuominen | 2009 | BETULA, PICEA, PINSYL, POPTRE | S Finland |
| 5 | Hidding et al. | 2013 | ABIBAL, BETULA, PICEA, POPTRE, PRUPEN | Quebec, CAN |
| 6 | Jensen et al. | 2012 | QUERCUS | S Sweden |
| 7 | Kielland & Bryant | 1998 | SALIX | Alaska, USA |
| 8 | Kolstad et al. | 2018 | BETULA, PICEA, PINSYL, SORBUS | S and C Norway |
| 9 | Leonardson et al. | 2015 | CONIFER | S Sweden |
| 10 | Nes | 2015 | ABIBAL, BETULA, PICEA, PINSYL, SORBUS | Quebec, CAN, S and C Norway |
| 11 | Nosko et al. | 2020 | ABIBAL | Newfoundland, CAN |
| 12 | Siipilehto & Heikkilä | 2005 | PINSYL | C Finland |
| 13 | Speed et al. | 2013 | BETULA, PICEA, PINSYL, POPTRE, SALIX, SORBUS | S and C Norway |
| 14 | Thompson et al. | 1992 | ABIBAL, BETULA, POPTRE, PRUPEN | Newfoundland, CAN |
The average observation time after moose exclusion was 10.2 ± 3.8 SD years for height, and 8.1 ± 4.3 SD years for density assessments. From studies that reported results for several years, we chose those from the last observation year.
Meta-analyses were conducted using the ‘meta’ package in R. We considered random-effects models, due to different experimental locations and conditions and different tree species studied, and subgroups, indicated by tree species or genus (Betula, Picea, Pinus, Abies, Salix and Sorbus). We used the groups “coniferous seedlings” (CONIFER) or “deciduous seedlings (DECID) when the studies did not distinguish seedlings by species.
Effect sizes, expressed as Cohen’s d-values were calculated using Eqs. 1 and 2 (Cohen 1988):

where M2 is mean of group 2 (moose exclusion) and M1 is mean of group 1 (control), SD is standard deviation, and:

where SD1 is standard deviation of group 1 (control) and SD2 is standard deviation of group 2 (moose exclusion).
We used moose densities for the analyses when mentioned in the publications. Overall, moose density is challenging to measure, and it is commonly estimated using winter aerial surveys, but also other approaches exist, like estimations from pellets or camera trapping (Moll et al. 2022). Average winter-time moose densities in Finland range from almost 5 per 1000 ha in the South to approx. 1 per 1000 ha in the North, and the density range of densities is between 1 and 8 per 1000 ha (Nikula et al. 2021). Local moose densities can deviate from average numbers, and regionally much higher densities have been reported, especially in other Nordic countries or Northern America (Suominen et al. 1999; Speed et al. 2013; Faison et al. 2016; Nosko et al. 2020).
Some studies (Heikkilä et al. 2003; Nosko et al. 2020) reported overall averages for several exclosure experiments but also presented data for each. These individual areas were 100 km or more apart from each other and could thus be regarded as independent from each other. Consequently, we used the values for each area for the analyses. Density and height subgroups for the same tree genus but different species, or same species but different location were created for Betula, Picea, Salix and Sorbus (Table 2), since one-way ANOVA tests did not indicate statistically significant differences between effect sizes (density: 4 < df < 16, 0.336 < F <0.904, 0.566 < P <0.913; height: 4 < df < 9, 0.025 < F < 9.892, 0.0944 < P <0.593).
| Table 2. Abbreviations of the tree species or genera used in the meta-analyses, assessing effect sizes of moose exclusion on seedling density and height, and sources (referring to Table 1) for analyses of density or height, respectively. | ||||
| Abbreviation | Tree species | Density | Height | |
| 1 | ABIBAL | Abies balsamea (L.) Mill. | 5, 10, 14 | 5, 11, 14 |
| 2 | BETULA | Betula papyrifera Marshall | 5, 10, 14 | 5, 14 |
| 3 | BETULA | Betula pendula Roth & B. pubescens Ehrh. | 3, 8, 10, 13 | 3, 4 |
| 4 | CONIFER | coniferous seedlings | 9 | |
| 5 | DECID | deciduous seedlings | 2 | |
| 6 | PICEA | Picea abies (L.) H. Karst. | 3, 8, 10, 13 | 3, 4 |
| 7 | PICEA | Picea glauca (Moench) Voss | 5, 10 | 5 |
| 8 | PICEA | Picea mariana (Mill.) Britton, Sterns & Poggenb. | 5 | |
| 9 | PINSYL | Pinus sylvestris L. | 3, 8, 10, 13 | 3, 4, 12 |
| 10 | POPTRE | Populus tremula L. | 5, 13, 14 | 4, 14 |
| 11 | PRUPEN | Prunus pensylvanica L. f. | 5, 14 | 14 |
| 12 | QUERCUS | Quercus robur L. & Q. petraea (Matt.) Liebl. | 6 | 6 |
| 13 | SALIX | Salix alaxensis (Andersson) Coville | 7 | |
| 14 | SALIX | Salix brachycarpa Nutt. | 7 | |
| 15 | SALIX | Salix caprea L. | 3, 13 | 1, 3 |
| 16 | SALIX | Salix lasiolepis Benth. | 7 | |
| 17 | SALIX | Salix novae-angliae Andersson | 7 | |
| 18 | SORBUS | Sorbus americana Marshall | 10 | |
| 19 | SORBUS | Sorbus aucuparia L. | 3, 8, 10, 13 | 3 |
To evaluate the observed heterogeneity and to test whether asymmetry (e.g., small-study effects) existed in the data (Balduzzi et al. 2019), we plotted funnel plots for the density and height data for deciduous and coniferous seedlings separated, indicating standard errors in relation to the respective effect sizes (‘funnel’ command in the ‘meta’ package in R). In addition, we produced the Egger’s tests (Egger et al. 1997) using the ‘metabias’ command in the ‘meta’ package for assessing possible publication bias of the data.
We related the calculated effect sizes (1) to the respective proportion of deciduous seedlings or saplings and (2) to the total seedling densities of the control treatments (moose exclosures). This was done separately for deciduous and coniferous species. Similarly, we calculated regressions between effect sizes and the observation time since moose exclusion, or for the estimated moose winter population density of the area (obtained from the respective publications if available). Regressions were calculated using the ‘lm’ command in R.
All calculations were conducted with R 4.2.1 (R Core Team 2022).
3 Results
3.1 Moose exclusion effects on seedling density and height
The meta-analysis for coniferous seedling densities indicated non-existing effects of moose exlosure (Fig. 1). Balsam fir (Abies balsamea (L.) Mill.) showed a somewhat positive effect and Norway spruce a slightly negative effect, however. In contrast, the effect sizes were more variable for coniferous seedling height and were positive for balsam fir. Overall, the results showed a significantly positive effect of moose exclosure for coniferous seedling height (Fig. 1). This indicated that coniferous seedlings were taller when moose was excluded in the fenced plots compared to the non-fenced controls.
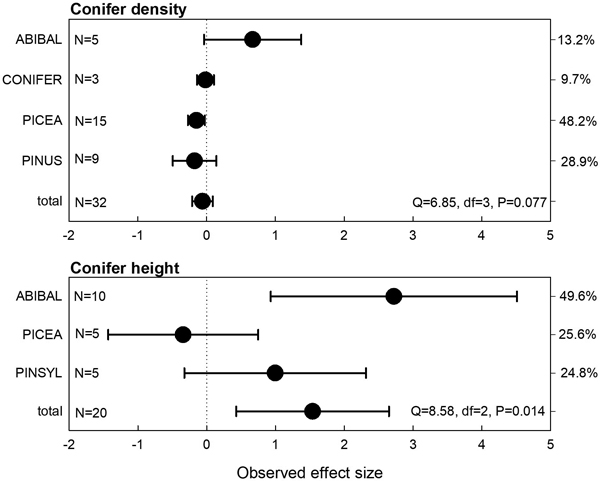
Fig. 1. Observed effect sizes for coniferous seedling or sapling density and height (treatment = moose exclosure) for individual tree species or genera (abbreviations explained in Table 2). Negative values indicate lower seedling densities and positive values higher seedling densities in the moose exclosure plots compared to the non-fenced control plots. Numbers of observations and heterogeneity test results are shown. The right y-axis depicts the weight of observational groups.
The meta-analysis indicated significant positive effects both for deciduous seedling densities and height (Fig. 2). Generally, the effect sizes were more variable than those for coniferous seedlings, indicated by significant heterogeneity test results. Moose exclusion density effects were positive for all deciduous species, but for Betula spp, Salix spp. and Sorbus spp. it was not statistically significant. Similarly, moose exclusion height effects for deciduous seedlings for all positive, but not significantly so for aspen and bird cherry (Prunus pensylvanica L. f.) (Fig. 2).
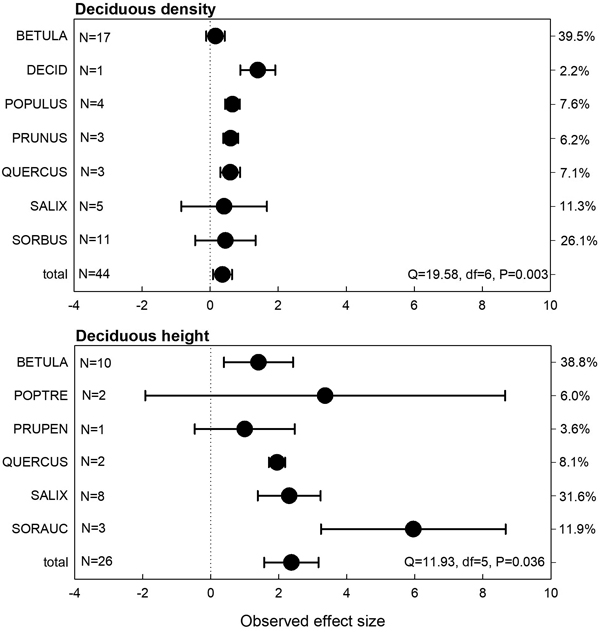
Fig. 2. Observed effect sizes for deciduous seedling or sapling density and height (treatment = moose exclosure) for individual tree species or genera (abbreviations explained in Table 2). Negative values indicate lower seedling densities and positive values higher seedling densities in the moose exclosure plots compared to the non-fenced control plots. Numbers of observations and heterogeneity test results are shown. The right y-axis depicts the weight of observational groups.
3.2 Factors affecting effect sizes of moose exclusion
The effect of the deciduous seedling proportion within the stand on the effect size of moose exclusion regarding seedling density and height differed between deciduous and coniferous seedlings. For deciduous seedling density, the moose exclusion effect sizes increased with increasing deciduous seedling proportions, whereas they decreased for coniferous seedlings (Fig. 3A). The deciduous seedling proportion did not impact the height effect of moose exclusion for deciduous seedlings, and the effect was always positive (Fig. 3B). However, it had a significant and decreasing effect on the height of coniferous seedlings, indicating that with higher deciduous seedling proportions, the difference of the moose browsing effect between moose exclusion and control plots decreased, and reached zero at a deciduous seedling proportion of approximately 90% (Fig. 3B).
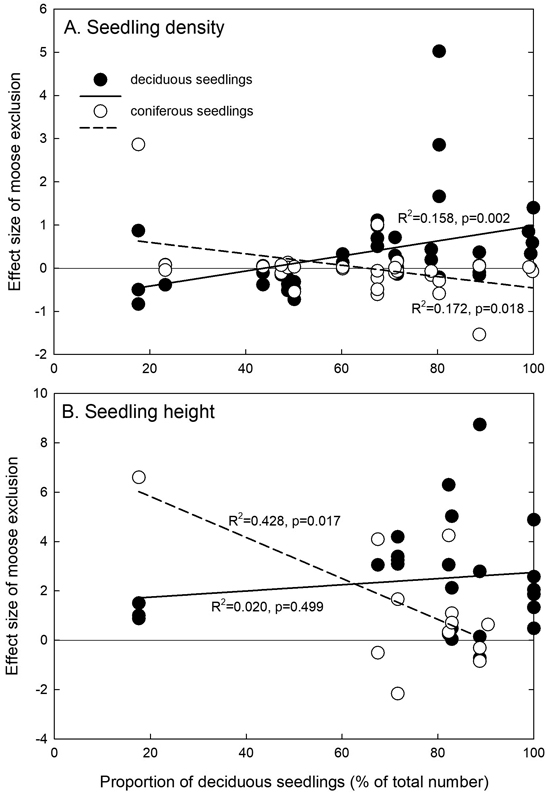
Fig. 3. Relationships between the proportion of deciduous seedlings or saplings (as percentage of the total number) and the effect size of moose exclusion for seedling or sapling (A) density and (B) height. Linear regressions are calculated separately for deciduous and coniferous seedlings. The zero line is indicated, and positive effects sizes indicate positive effects of moose exclusion.
Moose exclusion effects increased with increasing total seedling density in the plots both for deciduous and coniferous seedling density (Fig. 4A), indicating that the seedling density between fenced and control plots increased the higher the seedling density. It increased also for coniferous seedling height, and almost significantly so, but no effects were observed for deciduous seedling height (Fig. 4B).
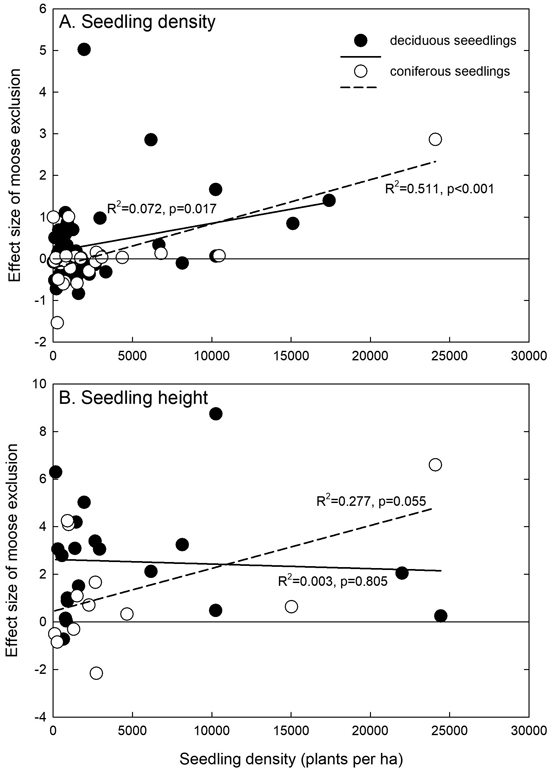
Fig. 4. Relationships between the total stand density (as plants per ha) and the effect size of moose exclusion for seedling or sapling (A) density and (B) height. Linear regressions are calculated separately for deciduous and coniferous seedlings. The zero line is indicated, and positive effect sizes indicate positive effects of moose exclusion.
Overall, we observed positive trends for the elapsed time on the effect size of moose exclusion since the start of fencing (Fig. 5), but it was significant only in the case of deciduous seedling density (Fig. 5A).
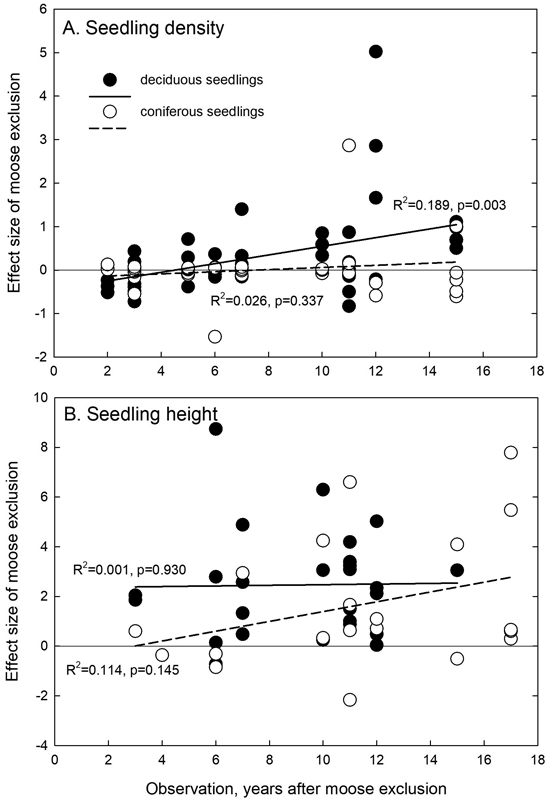
Fig. 5. Relationships between the observation time (years after moose exclusion) and the effect size of moose exclusion for seedling or sapling (A) density and (B) height. Linear regressions are calculated separately for deciduous and coniferous seedlings. The zero line is indicated, and positive effect sizes indicate positive effects of moose exclusion.
Like the effect of the deciduous seedling proportion on effect sizes, also the estimated moose density had opposite effects on deciduous and coniferous seedlings (Fig. 6). For deciduous seedlings, the effect was negative, and it was significant for seedling density (Fig. 6A). With increasing moose density, the moose exclusion effect increased for coniferous seedlings, and it was statistically significant for seedling height (Fig. 6B). The averages ± SD of the reported mean moose densities for seedling height were 15.1 ± 16.4 and for seedling density 18.9 ± 10.9 per 1000 ha, respectively.
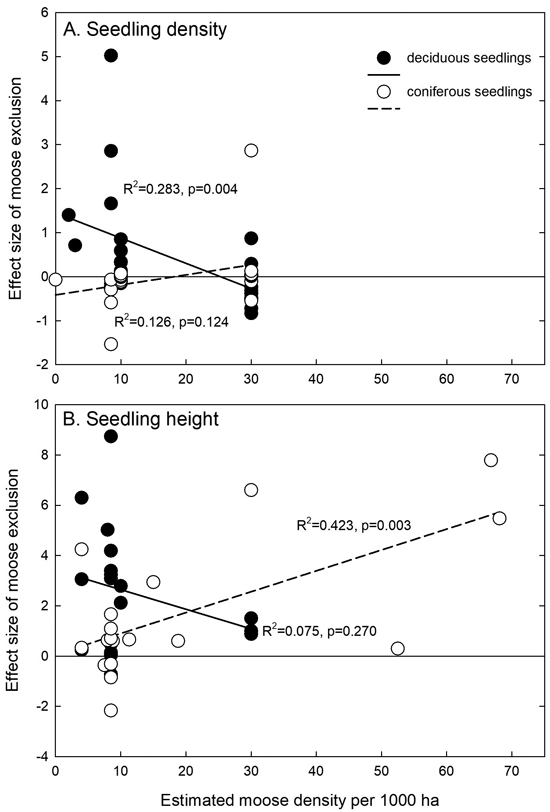
Fig. 6. Relationships between the estimated moose density and the effect size of moose exclusion for seedling or sapling (A) density and (B) height. Linear regressions are calculated separately for deciduous and coniferous seedlings. The zero line is indicated, and positive effect sizes indicate positive effects of moose exclusion.
3.3 Evaluation of meta-analyses
When judged visually, the funnel plots suggested some asymmetry and potentially small study bias (Figures S1 and S2 in Supplementary file S1), but the Egger’s test indicated that the data still could be regarded as symmetric and unbiased in regard of small study effects (deciduous seedling density: t = 1.79, df = 42, P = 0.0806; coniferous seedling density: t = –0.51, df = 30, P = 0.6165: deciduous seedling height: t = 2.61, df = 24, P = 0.0855: coniferous seedling height: t = 1.90, df = 18, P = 0.0732).
4 Discussion
4.1 Factors affecting moose browsing
We hypothesised that by controlling species composition and abundance and thus the forage available for moose, it would be possible to decrease moose damage in stands that were regenerated with conifers. This was verified to some extent, as we found that the proportion of deciduous seedlings or total seedling density in a stand affected the effect size of moose exclusion. We observed a somewhat opposite trend of deciduous seedling proportion for deciduous and coniferous seedlings, both for seedling density and height. For deciduous seedling density, the effect sizes increased with increasing proportion of deciduous seedlings, whereas for coniferous seedlings it was the opposite. The exclosure effect was decreasing for coniferous seedling height, too, but the slope was much steeper. This seems logical, as we can expect that moose browsing would affect seedling density less than its height, if not resulting in lethal damage. Similarly, Nosko et al. (2020) observed taller seedlings inside fenced areas compared to outside, for the same diameter, implying that moose affected the height growth and seedling shape. McLaren et al. (2009) concluded that when seedling stands were fenced, one suppressing factor (moose browsing) was replaced with another suppressing factor (competing fast growing deciduous tree species). Decreasing effect sizes with increasing deciduous seedling proportions seem plausible, considering a protective effect, and at some point, the effect sizes turn to negative, indicating that a high proportion of deciduous seedlings impairs the conifer growth and that conifer seedlings outside the exclosures might benefit from moose browsing of deciduous trees. On the other hand, exclosure effects for coniferous seedlings increased with increasing total stand density, indicating that moose damage to coniferous seedlings increased with rising stand density.
The positive moose exclusion effect for deciduous seedling density with elapsing time seems to indicate natural regeneration of deciduous seedlings with time in the exclusion areas but not in the open ones. This simply shows the effective fencing impact. No effects of elapsing time on seedling height implies that seedling height is more affected by other factors.
Furthermore, we hypothesised that it would be possible to recognise factors that increase the risk of moose browsing, and to identify measures to decrease this risk. One of the most important factors is moose density, but there are also other factors that can be identified as risk factors. An admixture of deciduous seedlings has been reported to increase moose browsing of coniferous seedlings (Bergqvist et al. 2014), but our data showed that with increasing proportion of deciduous trees moose exclusion effect sizes decreased. The decreasing effect of moose exclusion with increasing deciduous seedling density implies a protective effect: the higher deciduous seedling proportion in the stand, the smaller the moose effect on coniferous seedlings. However, in our data, the decreasing effect size in relation to the proportion of deciduous trees was greatly affected by two balsam fir sites with a low proportion of deciduous trees and extremely high moose population. If only Scots pine sites would have been analysed, the decrease in effect size would have not been detected. This points out the differences between coniferous species and the need for more species-specific analysis for practical applications.
From a silvicultural point of view, a high browsing pressure is especially harmful to pine seedling and sapling stands (Wallgren et al. 2013), and an admixture of preferred deciduous tree species like rowan, aspen, and birch, can even increase this risk (Heikkilä and Härkönen 1996; Nevalainen et al. 2016). On the other hand, preferred species can be protected by concealing, as observed by Jensen et al. (2012) for oak seedlings, which were protected by shrubs against moose browsing. Bergqvist et al. (2014) concluded that one efficient way for reducing the proportions of damaged Scots pine could be increasing the absolute number of pines and taking care that the pine seedlings are not overgrown by birch. Similar results were observed by Heikkilä and Mikkonen (1992) and Ball and Dallgren (2002), where pine density, site productivity as well as birch density increased total browsing, but the relative proportion of browsed pines decreased at high pine density. If the pine density is higher than 1500–2500 trees ha–1, preferably over 4000–5000 trees ha–1, moose damage decreases because the density of browsed pines follows less than a 1:1 ratio slope (Andrén and Angelstam 1993; Bergqvist et al. 2014). On the other hand, if densities were even higher, moose damage can increase again (Lyly and Saksa 1992), most probably due to shelter for moose against predation and abundant forage in the dense stands. Our results, showing increasing exclosure effects with increasing stand density, confirm these observations.
In theory, we could determine the critical percentage of deciduous admixture, when effect sizes turn negative, and fencing had a negative effect on coniferous seedlings density and height. For coniferous seedling density this happened at 65% deciduous admixture, and for coniferous seedlings height at 90%. This would mean that until a certain degree, coniferous seedlings would be protected by a deciduous admixture from moose browsing, in terms of seedling density until a deciduous proportion of 65% and regarding seedling height until a proportion of 90%. It seems reasonable, that the protective effect decreases with increasing deciduous proportion, but as there was a large variation in several affecting factors, like seedling size, soil, climate, and tree species, it is impossible to give an exact value when the protective effect would turn into a clearly hampering effect.
An important issue when addressing pine browsing damage is the season when damage occurs. Winter browsing is almost entirely directed to pine (Cederlund et al. 1980; Bergqvist et al. 2018), whereas during summer, also other preferred tree species are available. There are observations, that if available, willow and aspen are preferred over pine (Nichols et al. 2015). Given this fact, pine stands could include especially aspen and willow, so that moose would be attracted to other species than pine. Also, considering biodiversity, an admixture of deciduous trees is of great importance. However, other problems might occur, like pine twisting rust (Melampsora pinitorqua (Braun) Rostr.), infecting pine shoots and using aspen as alternate host (Mattila et al. 2001).
One future challenge will be to increase the share of mixed forests, but maintaining silvicultural quality in a way that it is not impaired by moose browsing. In this context, large herbivores can be a threat to biodiversity by decreasing the abundance of preferred deciduous species (Edenius et al. 2002). However, the preferred deciduous species usually occur at high densities, and moose browsing would not threaten their abundance as such. Impaired tree quality, resulting from moose browsing could be of some issue during later stand development (Härkönen et al. 2009; Matala et al. 2020). In addition, moose can increase spatial heterogeneity at stand level by affecting tree species abundance and proportion in a patchwork type pattern (Edenius et al. 2002). Considering mixed forests, certain tree species are preferred and could attract moose, but on the other hand, Milligan and Koricheva (2013) did not find any evidence of associational resistance. This was indicated by the fact that the presence of a least preferred species (spruce) in a mixture had no significant effect on moose browsing on other tree species. In Norway, spruce could benefit from intensive moose browsing on rowan (Hjeljord and Grønvold 1988).
There are indications that moose damage risk is higher on fertile and organic soils and in stands with heavy soil preparation (Nikula et al. 2008; Ruuhola et al. 2016). Fertile soils regenerated to Scots pine are often found in former Norway spruce stands and one alternative would be to use a tree species with a reduced risk of browsing damage, e.g., spruce on these sites (Lodin et al. 2017). However, given the higher abiotic and biotic damage risks of spruce, together with a lower potential of wood yield production, this can lead to non-optimal result from a forestry point of view. Also, sustaining high moose populations in combination with the increased establishment of spruce is likely to increase browsing pressure on remaining pine stands (Wallgren et al. 2013; Bergqvist et al. 2014).
Silvicultural early cleaning is needed for regulating early succession stand structure and proportions between competing tree species, e.g., pine and birch. Cleaning of birches has reduced the damage to pine trees in several studies (McLaren et al. 2000; Bergqvist et al. 2001; Karlsson et al. 2002; Härkönen et al. 2008). Intensive cleaning may increase the browsing pressure and damage severity to conifer species (Heikkilä and Mikkonen 1992; Lyly and Saksa 1992; McLaren et al. 2000). The timing of early cleaning or precommercial thinning is decisive, as birch will benefit from early measures, and the positive effect will be lost, whereas later actions will not have the desired effect. The importance of timing will be emphasized in mixed forest management to maintain vital tree species mixtures in the future. Generally, silvicultural recommendations in Sweden advise to wait until the stand has reached a “moose-safe height” of approx. 4.5 m to avoid severe moose damage (Ball and Dahlgren 2002). Härkönen et al. (2008) concluded that pine stand cleaning decreases moose damage, which could be attributed to decreased competition between overtopping deciduous trees and pine, resulting in increased pine growth, or a decreased attraction of moose into cleaned stands. De Vriendt et al. (2020) suggested that moose browsing could be used as a cheaper alternative to cleaning, whereas Härkönen (1998) concluded that moose browsing does not compensate for the lack of silvicultural cleaning.
4.2 Evaluation of meta-analyses
The results of the meta-analyses showed consistent results to those of the regressions, in that moose exclusion affected more seedling height than seedling density. This is not surprising, as they are based on the same data. However, conclusions covering different topics can be drawn from the conducted analyses. The regressions allow conclusions based on deciduous seedling proportions or overall seedling densities in the stand, whereas the forest plots allow more detailed conclusions concerning moose effects on different tree species. The negative effects, although non-significantly so, for spruce are consistent with the fact, that spruce is not a preferred species as a diet for moose. On the other hand, the highly significant and positive effects for willow and rowan indicate a high preference of these species. This is in accordance with previous knowledge (Bergström and Hjeljord 1987; Edenius 1991; Nichols et al. 2015). Also, the meta-analyses very clearly showed that balsam fir, although being a conifer, is a very preferred species, and thus compares more to deciduous trees. Similar observations were made by Bergerud and Manuel (1968) in Newfoundland, concluding, that with increasing fir density, also the percentage of browsed firs increased. The very high local moose density of over 30 moose per 1000 ha practically prevented birch and fir regeneration.
The number of publications that we used (14 papers) could be regarded as rather small. However, the actual data used for the meta-analyses was larger, as many publications had used several different experiments that could be regarded as independent from each other. For example, even within Central Finland, the different experiments were separated from each other at distances of over 100 km (Heikkilä et al. 2003). Even longer distances between experiments that were reported in the same publication, were in Southern and Central Norway (Speed et al. 2013) or even Canada and Norway (Nes 2015).
No asymmetry was observed in the density data, while it was seen in the height data (Suppl. file S1: Figs. S1 and S2). Publication bias is not the only reason for asymmetry, though (Egger et al. 1997; Page et al. 2020). We believe that between-study heterogeneity was the main reason for the observed asymmetry, resulting from different locations and thus climatic conditions and most probably from different tree species (being often confounded with location), and high moose densities. The data shows that effect sizes were the biggest for rowan and balsam fir, both preferred species by moose. Egger et al. (1997) state, that funnel plots are based on the fact, that precision in estimating the treatment effect will increase as the sample size increases. The assumption is, that the results from small studies scatter extensively at the bottom of the graph, and narrowing with larger studies towards the top, forming an upside-down funnel (Egger et al. 1997). The fact, that we separated different tree species from the same study, evidently added heterogeneity to the data. But as moose effects are very different for different tree species, we believe that reporting combined tree species effects for each publication was not a feasible option.
It sounds plausible that moose exclusion effect sizes would increase with higher moose densities. However, we observed this increase only for the height of coniferous seedlings. Moose densities represent estimations for the winter populations for rather large areas, though, and thus might not be entirely representative for the study plots. Small-scale moose density is dependent on many more factors than only food availability, like soil fertility or shelter. However, no data existed for deciduous seedling height at very high moose densities (Fig. 6B; Nosko et al. 2020), which might have been a result of the locally very high moose densities (Bergerud and Manuel 1968). Moose densities in those studies concentrating on silver birch or silver birch, Norway spruce or Scots pine, of which the majority were conducted in Finland, generally were lower than 10 moose per 1000 ha (observations concentrated on the left side of Fig. 6). Moose density is obviously a factor adding heterogeneity to the results, as seen in the visually observed asymmetry (Suppl. file S1: Figs. S1 and S2). It is a factor that cannot be controlled in the exclosure experiments. Funnel plot outliers to the positive side are mostly due to high moose densities in the respective research areas (usually 40–70 individuals per 1000 ha), whereas those to the negative side were due to low tree numbers of the respective species, indicating that they were not main tree species.
5 Conclusions
Our results showed that moose browsing damage increases with stand density, especially for coniferous seedlings. Seedling height was affected more by moose browsing than seedling density, and the tree species that were affected most by moose browsing were oak, rowan, and willow. Seedlings of the coniferous balsam fir were an exception, as they were affected in a similar way than deciduous tree seedlings, presumably due to extremely high moose densities. Our results also revealed a protective effect of deciduous seedlings on conifer seedlings, which was shown by decreasing moose exclusion effects with increasing proportions of deciduous seedlings in fenced plots. However, when deciduous seedling densities rose to a level too high, the negative effects were pronounced, presumably though increased competition. We conclude that generally, an appropriate deciduous admixture in conifer-dominated mixed seedling stands can have protective effects against moose damage, but more research is needed for practical recommendations for different conifer species. It is also noteworthy that timely silvicultural measures are important to decrease competitive effects of deciduous seedlings.
Declaration of openness of research materials, data, and code
Metadata can be obtained at https://doi.org/10.5281/zenodo.12188753, and codes are available upon request from the corresponding author (timo.domisch@luke.fi).
Author contributions
All authors contributed to the conceptualization and design of the work. TD did the data acquisition and analysed the data. All authors participated in the interpretation of data and results. TD wrote the manuscript draft and all authors participated in reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding of the research
This research was funded by the Natural Resources Institute Finland (research project “Can mixed forests provide a wide range of ecosystem services in the future?”) and by the ‘Catch the Carbon Programme’ of the Ministry of Agriculture and Forestry of Finland (research project “Impact of cervid species on forest health and growth from the perspective of climate change targets”).
Ethical issues
None.
References
Andrén H, Angelstam P (1993) Moose browsing on Scots pine in relation to stand size and distance to forest edge. J Appl Ecol 30: 133–142. https://doi.org/10.2307/2404277.
Angell AC, Kielland K (2009) Establishment and growth of white spruce on a boreal forest floodplain: interactions between microclimate and mammalian herbivory. For Ecol Manag 258: 2475–2480. https://doi.org/10.1016/j.foreco.2009.08.030.
Angelstam P, Wikberg PE, Danilov P, Faber WE, Nygrén K (2000) Effects of moose density in timber quality and biodiversity in Sweden, Finland and Russian Karelia. Alces 36: 133–145. https://alcesjournal.org/index.php/alces/article/view/639.
Balduzzi S, Rücker G, Schwarzer G (2019) How to perform a meta-analysis with R: a practical tutorial. Evid Based Ment Health 22: 153–160. https://doi.org/10.1136/ebmental-2019-300117.
Ball JP, Dallgren J (2002) Browsing damage on pine (Pinus sylvestris and P. contorta) by a migrating moose (Alces alces) population in winter: relation to habitat composition and road barriers. Scand J For Res 17: 427–435. https://doi.org/10.1080/028275802320435441.
Barbosa P, Hines J, Kaplan I, Martinson H, Szczepaniec A, Szendrei Z (2009) Associational resistance and associational susceptibility: having right or wrong neighbors. Annu Rev Ecol Evol Syst 40: 1–20. https://doi.org/10.1146/annurev.ecolsys.110308.120242.
Bergerud AT, Manuel F (1968) Moose damage to balsam fir-white birch forests in Central Newfoundland. J Wildl Manag 32: 729–746. https://doi.org/10.2307/3799547.
Bergqvist G, Bergström R, Edenius L (2001) Patterns of stem damage by moose (Alces alces) in young stands of Pinus sylvestris. Scand J For Res 16: 363–370. http://dx.doi.org/10.1080/02827580119307.
Bergqvist G, Bergström R, Wallgren M (2011). Browsing by large herbivores on Scots pine (Pinus sylvestris) seedlings in mixture with ash (Fraxinus excelsior) or silver birch (Betula pendula). Scand J For Res 27: 372–378. https://doi.org/10.1080/02827581.2011.635155.
Bergqvist G. Bergström R, Wallgren M (2012). Summer browsing by moose on Scots pine. Scand J For Res 28: 110–116. https://doi.org/10.1080/02827581.2012.725767.
Bergqvist G, Bergstrom R, Wallgren M (2014) Recent browsing damage by moose on Scots pine, birch and aspen in young commercial forests – effects of forage availability, moose population density and site productivity. Silva Fenn 48, article id 1077. https://doi.org/10.14214/sf.1077.
Bergström B, Hjeljord O (1987) Moose and vegetation interactions in northwestern Europe and Poland. Swedish Wildl Res 1: 213–228.
Bergström R, Jernelid H, Lavsund S, Lundberg P, Wallin K (1995) Älgtäthet – betestryck –fodertillgång – skogstillstånd – skadenivåer – skaderisker. [Moose density –browsing pressure – forage availability – forest composition – forest damage – damage risks]. Projekt Balanserad Älgstam, Final report. Swedish Hunters Organisation and Swedish University of Agricultural Sciences (SLU).
Cederlund G, Markgren G (1987) The development of the Swedish moose population, 1970–1983. Swedish Wildl Res 1: 55–62.
Cederlund G, Ljungquist H, Markgren G, Stalfelt F (1980) Foods of moose and roe deer at Grimsø in central Sweden; results of rumen content analyses. Swedish Wildl Res 11: 169–247.
Cohen J (1988) Statistical power analysis for the behavioral sciences, 2nd edition. Routledge, New York.
Danell K, Ericsson L (1986) Foraging by moose on two species of birch when these occur in different proportions. Ecography 9: 79–84. https://doi.org/10.1111/j.1600-0587.1986.tb01194.x.
Danell K, Huss-Danell K, Bergström R (1985) Interactions between browsing moose and to birch species in Sweden. Ecology 66: 1867–1878. https://doi.org/10.2307/2937382.
de Chantal M, Lindberg H, Kallonen S (2009) The condition and survival of Populus tremula and other deciduous saplings in a moose winter-foraging area in southern Finland. Ann Bot Fennici 46: 280–290. https://doi.org/10.5735/085.046.0404.
De Vriendt L, Thiffault N, Royo AA, Barrette M, Tremblay JP (2020) Moose browsing tends spruce plantations more efficiently than a single mechanical release. Forests 11, article id 1138. https://doi.org/10.3390/f11111138.
Edenius L (1991) The effect of resource depletion on the feeding behavious of a browser: winter foraging by moose on Scots pine. J Appl Ecol 28: 318–328. https://doi.org/10.2307/2404132.
Edenius L, Bergman M, Ericsson G, Danell K (2002) The role of moose as a disturbance factor in managed boreal forests. Silva Fenn 36: 57–67. https://doi.org/10.14214/sf.550.
Egger M, Smith GD, Schneider M, Minder C (1997) Bias in meta-analysis detected by a simple, graphical test. BMJ 315: 629–634. https://doi.org/10.1136/bmj.315.7109.629.
Faber WE, Lavsund S (1999) Summer foraging on Scots pine Pinus sylvestris by moose Alces alces in Sweden – patterns and mechanisms. Wildl Biol 5: 93–106. https://doi.org/10.2981/wlb.1999.013.
Faison EK, DeStefano S, Foster DR, Rapp JM, Compton JA (2016) Multiple browsers structure tree recruitment in logged temperate forests. PLoS ONE 11, article id e0166783. https://doi.org/10.1371/journal.pone.0166783.
Felton A, Nilsson U, Sonesson J, Felton AM, Roberge J-M, Ranius T, Ahlström M, Bergh J, Björkman C, Boberg J, Drössler L, Fahlvik N, Gong P, Holmström E, Keskitalo ECH, Klapwijk MJ, Laudon H, Lundmark T, Niklasson M, Nordin A, Pettersson M, Stenlid J, Sténs A, Wallertz K (2016) Replacing monocultures with mixed-species stands: ecosystem service implications of two production forest alternatives in Sweden. Ambio 45: 124–139. https://doi.org/10.1007/s13280-015-0749-2.
Härkönen S (1998) Effects of silvicultural cleaning in mixed pine‐deciduous stands on moose damage to Scots pine (Pinus sylvestris). Scand J For Res 13: 429–436. https://doi.org/10.1080/02827589809383003.
Härkönen S, Miina J, Saksa T (2008) Effect of cleaning methods in mixed pine–deciduous stands on moose damage to Scots pines in southern Finland. Scand J For Res 23: 491–500. http://dx.doi.org/10.1080/02827580802491371.
Härkönen S, Pulkkinen A, Heräjärvi H (2009) Wood quality of birch (Betula spp.) trees damaged by moose. Alces 45: 67–72.
Heikkilä R (1991) Moose browsing in a Scots Pine plantation mixed with deciduous tree species. Acta For Fenn 224. https://doi.org/10.14214/aff.7670.
Heikkilä R, Mikkonen T (1992) Effects of density of young Scots pine (Pinus sylvestris) stand on moose (Alces alces) browsing. Acta For Fenn 231. https://doi.org/10.14214/aff.7677.
Heikkilä R, Härkönen S (1996) Moose browsing in young Scots pine stands in relation to forest management. For Ecol Manag 88: 179–186. https://doi.org/10.1016/S0378-1127(96)03823-6.
Heikkilä R, Tuominen M (2009) The influence of moose on tree species composition in Liesjärvi National Park in Southern Finland. Alces 45: 49–58. https://alcesjournal.org/index.php/alces/article/view/13.
Heikkilä R, Hokkanen P, Kooiman M, Ayguney N, Bassoulet (2003) The impact of moose browsing on tree species composition in Finland. Alces 39: 203–213.
Hidding B, Tremblay JP, Coté SD (2013) A large herbivore triggers alternative successional trajectories in the boreal forest. Ecology 94: 2852–260. https://doi.org/10.1890/12-2015.1.
Hjeljord O, Grønvold S (1988) Glyphosate application in forest – ecological aspects. VI. Browsing by moose (Alces alces) in relation to chemical and mechanical brush control. Scan J For Res 3: 115–121. https://doi.org/10.1080/02827588809382501.
Hörnberg S (2001) Changes in population density of moose (Alces alces) and damage to forests in Sweden. For Ecol Manag 149: 141–151. https://doi.org/10.1016/S0378-1127(00)00551-X.
Huuskonen S, Domisch T, Finér L, Hantula J, Hynynen J, Matala J, Miina J, Neuvonen S, Nevalainen S, Niinistö P, Nikula A, Piri T, Siitonen J, Smolander A, Tonteri T, Uotila K, Viiri H (2021) What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For Ecol Manag 479, article id 118558. https://doi.org/10.1016/j.foreco.2020.118558.
Jensen AM, Götmark F, Löf M (2012) Shrubs protect oak seedlings against ungulate browsing in temperate broadleaved forests of conservation interest: a field experiment. For Ecol Manag 266: 187–193. https://doi.org/10.1016/j.foreco.2011.11.022.
Jensen WF, Rea RV, Penner CE, Smith JR, Bragina EV, Razenkova E, Balciauskas L, Bao H, Bystiansky S, Csányi S, Chovanova Z, Done G, Hackländer K, Heurich M, Jiang G, Kazarez A, Pusenius J, Solberg EJ, Veeroja R, Widemo F (2020) A review of circumpolar moose populations with emphasis on Eurasian moose distributions and densities. Alces 56: 63–78. https://alcesjournal.org/index.php/alces/article/view/265.
Kielland K, Bryant JP (1998) Moose herbivory in taiga: effects on biogeochemistry and vegetation dynamics in primary succession. Oikos 82: 377–383. https://doi.org/10.2307/3546979.
Kolstad AL, Austrheim G, Solberg EJ, De Vriendt L, Speed JDM (2018) Pervasive moose browsing in boreal forests alters successional trajectories by severely suppressing keystone species. Ecosphere 9, article id e02458. https://doi.org/10.1002/ecs2.2458.
Korhonen KT, Ahola A, Heikkinen J, Henttonen HM, Hotanen J-P, Ihalainen A, Melin M, Pitkänen J, Räty M, Sirviö M, Strandström M (2021) Forests of Finland 2014–2018 and their development 1921–2018. Silva Fenn 55, article id 10662. https://doi.org/10.14214/sf.10662.
Lavsund S (1987) Moose relationship to forestry in Finland, Norway and Sweden. Swedish Wildl Res 1:229–244.
Lavsund S, Nygrén T, Solberg EJ (2003) Status of moose populations and challenges to moose management in Fennoscandia. Alces 39: 109–130. https://alcesjournal.org/index.php/alces/article/view/469.
Leonardson J, Löf M, Götmark F (2015) Exclosures can favour natural regeneration of oak after conservation-oriented thinning in mixed forests in Sweden: a 10-year study. For Ecol Manag 354: 1–9. https://doi.org/10.1016/j.foreco.2015.07.004.
Lodin I, Brukas V, Wallin I (2017) Spruce or not? Contextual and attitudinal drivers behind the choice of tree species in southern Sweden. Forest Policy Econ 83: 191–198. https://doi.org/10.1016/j.forpol.2016.11.010.
Lyly O, Saksa T (1992) The effect of stand density on moose damage in young Pinus sylvestris stands. Scand J For Res 7: 393–403. https://doi.org/10.1080/02827589209382732.
Månsson J, Kalén C, Kjellander P, Andrén H, Smith H (2007) Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scand J For Res 22: 407–414. https://doi.org/10.1080/02827580701515023.
Markgren G (1974) The moose in Fennoscandia. Le Naturaliste Canadien 101: 185–194.
Matala J, Kilpeläinen H, Heräjärvi H, Wall T, Verkasalo V (2020) Sawlog quality and tree dimensions of Scots pine 34 years after artificial moose browsing damage. Silva Fenn 54, article id 10389. https://doi.org/10.14214/sf.10389.
Mattila U, Jalkanen R, Nikula A (2000) The effects of forest structure and site characteristics on probability of pine twisting rust damage in young Scots pine stands. For Ecol Manag 142: 89–97. https://doi.org/10.1016/S0378-1127(00)00342-X.
Mattson L (1990) Moose management and the economic value of hunting: Towards bioeconomic analysis. Scand J For Res 5: 575–581. https://doi.org/10.1080/02827589009382640.
McLaren BE, Mahoney S, Porter TS, Oosenbrug SM (2000) Spatial and temporal patterns of use by moose of pre-commercially thinned, naturally-regenerating stands of balsam fir in central Newfoundland. For Ecol Manag 133: 179–196. https://doi.org/10.1016/S0378-1127(99)00223-6.
McLaren B, Hermanutz L, Gosse J, Collet B, Kasimos C (2009) Broadleaf competition interferes with balsam fir regeneration following experimental removal of moose. For Ecol Manag 257: 1395–1404. https://doi.org/10.1016/j.foreco.2008.12.009.
Milligan HT, Koricheva J (2013) Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: an experimental study. J Anim Ecol 82: 739–748. https://doi.org/10.1111/1365-2656.12049.
Moen R, Pastor J, Cohen Y (1997) A spatially explicit model of moose foraging and energetics. Ecology 78: 505–521. https://doi.org/10.2307/2266026.
Moll RJ, Poisson MKP, Heit DR, Jones H, Pekins PJ, Kantar L (2022) A review of methods to estimate and monitor moose density and abundance. Alces 58: 31–49. https://alcesjournal.org/index.php/alces/article/view/1881.
Nes T (2015) The effects of moose (Alces alces) browsing on boreal tree species in Norway and Quebec. MSc thesis, Department of Biology, Norwegian University of Science and Technology, Trondheim. http://hdl.handle.net/11250/2351525.
Nevalainen S, Matala J, Korhonen KT, Ihalainen A, Nikula A (2016) Moose damage in National Forest Inventories (1986–2008) in Finland. Silva Fenn 50, article id 1410. https://doi.org/10.14214/sf.1410.
Nichols RV, Cromsigt JPGM, Spong G (2015) DNA left on browsed twigs uncovers bite-scale resource use patterns in European ungulates. Oecologia 178: 275–284. https://doi.org/10.1007/s00442-014-3196-z.
Niedziałkowska M, Neumann W, Borowik T, Kołodziej-Sobocińska M, Malmsten J, Arnemo JM, Ericsson G (2022) In: Corlatti L, Zachos FE (eds) Terrestrial cetartiodactyla. Handbook of the mammals of Europe. Springer, Cham. https://doi.org/10.1007/978-3-030-24475-0_23.
Nikula A, Hallikainen V, Jalkanen R, Hyppönen M, Mäkitalo K (2008) Modelling the factors predisposing Scots pine to moose damage in artificially regenerated sapling stands in Finnish Lapland. Silva Fennica 42: 587–603. https://doi.org/10.14214/sf.235.
Nikula A, Matala J, Hallikainen V, Pusenius J, Ihalainen A, Kukko T, Korhonen KT (2021) Modelling the effect of moose Alces alces population density and regional forest structure on the amount of damage in forest seedling stands. Pest Manag Sci 77: 620–627. https://doi.org/10.1002/ps.6081.
Nosko P, Roberts K, Knight T, Marcellus A (2020) Growth and chemical responses of balsam fir saplings released from intense browsing pressure in the boreal forests of western Newfoundland, Canada. For Ecol Manag 460, article id 117839. https://doi.org/10.1016/j.foreco.2019.117839.
Nygrén T (1987) The history of moose in Finland. Swedish Wildl Res Suppl 1: 49–54.
Østgård J (1987) Status of moose in Norway in the 1970’s and early 1980’s. Swedish Wildl Res Suppl 1: 63–68.
Page MJ, Sterne JAC, Higgins JPT, Egger M (2020) Investigating and dealing with publication bias and other reporting biases in meta-analyses of health research: a review. Res Synth Methods 12: 248–259. https://doi.org/10.1002/jrsm.1468.
Pfeffer SE, Singh NJ, Cromsigt JPGM, Kalén, C, Widemo F (2021) Predictors of browsing damage on commercial forests – a study linking nationwide management data. For Ecol Manag 479, article id 118597. https://doi.org/10.1016/j.foreco.2020.118597.
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rohatgi A (2021) WebPlotDigitizer. https://automeris.io/WebPlotDigitizer.
Ruuhola T, Nikula A, Nivala V, Nevalainen S, Matala J (2016) Effects of bedrock and surficial deposit composition on moose damage in young forest stands in Finnish Lapland. Silva Fenn 50, article id 1565. https://doi.org/10.14214/sf.1565.
SFA (2018) Äbin – älgbetesinventering/. [Moose damage inventory]. Swedish Forest Agency. https://www.skogsstyrelsen.se/abin.
Siipilehto J, Heikkilä R (2005) The effect of moose browsing on the height structure of Scots pine saplings in a mixed stand. For Ecol Manag 205: 117–126. https://doi.org/10.1016/j.foreco.2004.10.051.
Speed JDM, Austrheim G, Hester AJ, Solberg EJ, Tremblay JP (2013) Regional-scale alteration of clear-cut forest regeneration caused by moose browsing. For Ecol Manag 289: 289–299. https://doi.org/10.1016/j.foreco.2012.10.051.
Spitzer R, Felton A, Landman M, Singh NJ, Widemo F, Cromsigt JPGM (2020) Fifty years of European ungulate dietary studies: a synthesis. Oikos 129: 1668–1680. https://doi.org/10.1111/oik.07435.
Spitzer R, Coissac E, Felton A, Fohringer C, Juvany L, Landman M, Singh NJ, Taberlet P, Widemo F, Cromsigt JPGM (2021) Small shrubs with large importance? Smaller deer may increase the moose-forestry conflict through feeding competition over Vaccinium shrubs in the field layer. For Ecol Manag 480, article id 118768. https://doi.org/10.1016/j.foreco.2020.118768.
Suominen O, Danell K, Bergström R (1999) Moose, trees, and ground-living invertebrates: indirect interactions in Swedish pine forests. Oikos 84: 215–226. https://doi.org/10.2307/3546716.
Wallgren MR, Bergström G, Bergqvist B, Olsson M (2013) Spatial distribution of browsing and tree damage by moose in young pine forests, with implications for the forest industry. For Ecol Manag 305: 229–238. https://doi.org/10.1016/j.foreco.2013.05.057.
Total of 79 references.
