Johanna Jetsonen  ,
Annamari Laurén,
Heli Peltola,
Katariina Laurén,
Samuli Launiainen,
Marjo Palviainen
,
Annamari Laurén,
Heli Peltola,
Katariina Laurén,
Samuli Launiainen,
Marjo Palviainen
Volume growth responses of Scots pine and Norway spruce to nitrogen fertilization: quantitative synthesis of fertilization experiments in Finland
Jetsonen J., Laurén A., Peltola H., Laurén K., Launiainen S., Palviainen M. (2025). Volume growth responses of Scots pine and Norway spruce to nitrogen fertilization: quantitative synthesis of fertilization experiments in Finland. Silva Fennica vol. 59 no. 1 article id 24041. https://doi.org/10.14214/sf.24041
Highlights
- Volume growth responses increased linearly with the applied nitrogen (N) dose up to 200 kg N ha–1
- Precipitation, site fertility, and time since fertilization affected the growth responses
- Volume growth response to nitrogen fertilization was larger for Scots pine than for Norway spruce
- Results can support planning of environmentally responsible and economically viable precision fertilization.
Abstract
The effects of nitrogen (N) fertilization on tree growth have been studied widely in boreal forests in Finland, but a quantitative synthesis is still lacking. We performed a quantitative synthesis on volume growth responses to N fertilization in Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karst.) stands in experiments established on mineral soils across Finland. Our study employed findings of 9 published studies including 108 Scots pine and 57 Norway spruce observations covering a wide range of N fertilization treatments, as well as forest stand- and climatic conditions. Based on these observations, we built linear mixed models to describe the N fertilization-induced annual volume growth response of Scots pine and Norway spruce stands. Our models showed that the N dose was the best predictor for volume growth response, and the growth response increased linearly with increasing N dose for both tree species. The volume growth responses also increased along with an increase in mean annual precipitation. The annual volume growth response decreased with the time since fertilization. For Scots pine, the best model also contained site fertility; increase in site fertility increased the volume growth response. These findings emphasize the need for site-specific precision fertilization schemes to sustainably improve growth and carbon sequestration of boreal forests.
Keywords
precipitation;
growth response;
mineral soils;
site fertility
-
Jetsonen,
Department of Forest Sciences, University of Helsinki, P.O. Box 27, 00014 University of Helsinki, Finland
 https://orcid.org/0009-0006-4878-8951
E-mail
johanna.jetsonen@helsinki.fi
https://orcid.org/0009-0006-4878-8951
E-mail
johanna.jetsonen@helsinki.fi
-
Laurén,
Department of Forest Sciences, University of Helsinki, P.O. Box 27, 00014 University of Helsinki, Finland; Faculty of Science and Forestry, University of Eastern Finland, P.O. Box 111, 80101 Joensuu, Finland
https://orcid.org/0000-0002-6835-9568
E-mail
annamari.lauren@helsinki.fi
-
Peltola,
Faculty of Science and Forestry, University of Eastern Finland, P.O. Box 111, 80101 Joensuu, Finland
https://orcid.org/0000-0003-1384-9153
E-mail
heli.peltola@uef.fi
-
Laurén,
Faculty of Science and Forestry, University of Eastern Finland, P.O. Box 111, 80101 Joensuu, Finland; University of Helsinki, Faculty of Medicine, Haartmaninkatu 8, 00290 Helsinki, Finland
https://orcid.org/0009-0009-4677-9826
E-mail
katariina.lauren@helsinki.fi
-
Launiainen,
Natural Resources Institute Finland (Luke), Latokartanonkaari 9, 00790 Helsinki, Finland
https://orcid.org/0000-0001-6611-6573
E-mail
samuli.launiainen@luke.fi
-
Palviainen,
Department of Forest Sciences, University of Helsinki, P.O. Box 27, 00014 University of Helsinki, Finland
https://orcid.org/0000-0001-9963-4748
E-mail
marjo.palviainen@helsinki.fi
Received 26 June 2024 Accepted 23 January 2025 Published 29 January 2025
Views 48478
Available at https://doi.org/10.14214/sf.24041 | Download PDF

Supplementary Files
1 Introduction
Boreal forests have an essential role in climate change mitigation because they can efficiently sequester and store carbon (C) into tree biomass and soil (Mäkipää 1995; Ilvesniemi et al. 2002; Bradshaw and Warkentin 2015). However, tree growth in boreal forests is commonly limited by nitrogen (N) availability (Tamm 1991). Nitrogen fertilization increases N supply and consequently tree growth, thus offering a way to rapidly increase the rate of C sequestration (Kellomäki et al. 1982; Sikström et al. 1998; Jörgensen et al. 2021).
The effects of N fertilization on tree growth in mineral soils have been studied widely in the boreal region. The results suggest that the magnitude and duration of the growth response are affected by multiple factors (Saarsalmi and Mälkönen 2001; Hedwall et al. 2014). The duration of the growth response depends on the tree species, site fertility and the geographical location. For Norway spruce (Picea abies (L.) Karst.) N fertilization has been observed to increase growth typically for 8–10 years and for Scots pine (Pinus sylvestris L.) 6–8 years following the fertilizer application (Laakkonen et al. 1983; Pettersson 1994). The growth response is more persistent in the northern boreal zone than in the southern boreal zone (Laakkonen et al. 1983; Pettersson 1994). In the southern boreal zone, however, the growth response is larger, because the warmer climatic conditions enhance potential photosynthesis, nutrient uptake, and nutrient cycling (Pettersson 1994; Philben et al. 2016). In previous studies, N fertilization increased the annual volume growth by 1–2 m3 ha–1 for Scots pine and Norway spruce stands during the first 6–10 years after fertilization (Kukkola and Saramäki 1983; Valinger et al. 2000).
The magnitude of the growth response can be increased by choosing appropriate properties and dose of the fertilizer (Kukkola and Saramäki 1983). According to the current practical forestry guidelines in Finland, the recommended dose of N is 100–200 kg ha–1 (Äijälä et al. 2019). According to previous studies, the annual volume growth response increases until the N dose is 250 kg ha–1, after which the volume growth response starts to saturate, and extra N supply does not markedly improve the growth anymore (Kukkola and Saramäki 1983; Pukkala 2017). Furthermore, the different N in fertilizers affect growth responses (Kukkola and Saramäki 1983). Common N compounds used for forest fertilization are ammonium nitrate, ammonium sulfate, urea, and urea-formaldehyde (Lehto and Ilvesniemi 2023). The ammonium compounds and urea are water soluble, and rapidly released into the soil, whereas the urea-formaldehyde is slowly mineralized into ammonium and nitrate (Jahns et al. 2000, 2003; Smolander et al. 2022). Fast release of the water-soluble N increases the risk of N leaching (Binkley et al. 1999) and may alter ground vegetation species composition (Jetsonen et al. 2024). The adverse environmental effects of N-fertilization can be decreased, and cost-efficiency of fertilization improved when the dose and properties of the N fertilizer meet the nutrient needs of the stands.
The effects of N fertilization have been studied extensively in Finland since the 1960s, and the results on the growth responses have been descriptively summed in literature reviews (Kukkola and Saramäki 1983; Pettersson 1994; Saarsalmi and Mälkönen 2001; Lehto and Ilvesniemi 2023). However, quantitative synthesis of the N fertilization experiments is still lacking. The results from single fertilization experiments or from descriptive verbal syntheses do not provide sufficient support for defining precision fertilization schemes which may sustainably improve growth and carbon sequestration of boreal forests.
In this study, we performed a quantitative synthesis on observed volume growth responses to N fertilization in Scots pine and Norway spruce stands in multiple fertilization experiments established on mineral soil site types across Finland from the 1960s to the 1990s. Based on these observations, we built predictive models to describe how much the N fertilization may increase annual volume growth of Scots pine and Norway spruce stands after the fertilization. We explored the effects of N fertilizer dose, geographical location, site type, initial stand volume, and time since fertilization on the volume growth response in Scots pine and Norway spruce stands. Based on previous knowledge, we hypothesized that 1) increasing N fertilizer dose increases the volume growth response, 2) the volume growth response is larger the more fertile the sites, and 3) the volume growth response is larger in the south than in the north. The results of our quantitative synthesis are expected to provide support to planning site-specific environmentally responsible precision fertilization schemes for Scots pine and Norway spruce stands.
2 Material and methods
2.1 Systematic literature survey
We searched for published research articles for forest N fertilization studies implemented in Finland from 1960 to 2023, which were established in Scots pine or Norway spruce stands on mineral soils. Peer reviewed articles and gray literature filling these criteria were searched using the pearl growing method, where more eligible articles are found from references of other eligible articles (Schlosser et al. 2006). We used keywords “nitrogen fertilization,” “Scots pine,” and “Norway spruce” to find the first studies. The eligible articles had to be available in Google Scholar, the open repository of the Natural Resources Institute Finland (Jukuri), or Library databases of the Universities of Oulu Eastern Finland and Helsinki to be included in the study.
The eligible articles were initially included in the dataset if they contained information on the amount of fertilizer applied (kg ha–1), nutrient concentration of the fertilizer, and a growth response variable, such as stand volume, height, basal area or diameter. In total 22 articles filled the initial inclusion criteria and the information about fertilization treatment and growth response were extracted from them. Also, information about the stand characteristics before the fertilization, such as initial stand volume, site type (Cajander 1949), and the location of each study site, were extracted from the primary studies into a dataset. Long-term (1961–1990) effective annual temperature sum and long-term mean annual precipitation were also included into the dataset. In case they were not reported, we retrieved them from the Finnish Meteorological Institute open data repository (https://en.ilmatieteenlaitos.fi/open-data). If information about any other variable was missing, the data entry was left empty. If the growth responses were presented in the studies in a graph form only, PlotDigitiser-program version 3.1.5 (https://plotdigitizer.com/app) was used to extract the values.
From four studies reporting findings of repeated fertilization, we included in our analyses only the growth data measured between the first and the second fertilizer application. Additionally, three articles reporting results of repeated fertilization experiments were excluded because the effect of first fertilization could not be extracted. Also, only stands with initial volume larger than 50 m3 ha–1 at the time of fertilization were included in the analyses because seedling stands with dominant height below 7 m are not fertilized in practical forestry in Finland; this criterion excluded one article. Furthermore, three articles that did not use N in the fertilization or used it mixed with ash, were also excluded because the aim was to study the effects of N fertilizer. After these exclusions we selected the growth response variable to be the volume growth because it had the highest number of observations. From the final data we excluded four articles that did not report the initial stand volume or the volume growth. Additionally, two articles were removed because they added only one observation each to the dataset; inclusion of these articles would have hindered evaluation of the random effect covariance matrix in the mixed model analysis.
We included 9 articles to the final dataset and these 9 articles providing the data for the synthesis are summarized in Table 1. In total, the dataset consisted of 108 Scots pine observations and 57 Norway spruce observations, covering a wide range of N dose, initial stand volumes, site types, and climatic conditions. The total number of study locations was 97, and they were located between latitudes 60°N–68°N (Fig. 1). Long-term (1961–1990) mean annual temperature for the study sites varied from –1.3 to 4.9 °C, long-term mean annual precipitation from 400 to 700 mm and long-term effective temperature sum from 740 to 1280 d.d. (Table 1).
| Table 1. Description of the nitrogen (N) fertilization studies from Finland, included in the dataset. FN is the applied N dose, MAAT the mean annual air temperature, Pa the annual precipitation, and Tsum the effective temperature sum. The site type was classified according to the Cajander (1949) site classification system. The initial stand volume (Vini) range indicates the stand structure before fertilization. SF is the time since fertilization in years and n is the number of observations included into the dataset. | |||||||||||
| Article | Latitude | FN (kg ha–1) | MAAT (°C) | Pa (mm) | Tsum (d.d.) | Tree species | Vini (m3 ha–1) | Site type | SF (yrs.) | n | Reference |
| 1 | 60°26´–67°54´N | 120 | –1.2–4.4 | 470–590 | 900–1180 | Pinus sylvestris, Picea abies | 113–200 | MT, VT, CT | 10 | 12 | Laakkonen et al. (1983) |
| 2 | 61°45´–67°17´N | 120 | –1.0–3.2 | 430–640 | 790–1280 | Pinus sylvestris, Picea abies | 122–175 | MT, VT | 5 | 68 | Lipas & Levula (1980) |
| 3 | 64°48´N | 156–208 | 1.8 | 596 | 990 | Pinus sylvestris | 81–123 | MT, VT | 11 | 10 | Moilanen & Meriluoto (1984) |
| 4 | 62°40´–64°07´N | 100 | 1.7 | 630 | 1090 | Pinus sylvestris, Picea abies | 50–230 | MT, VT, CT | 4 | 12 | Salonen (1973) |
| 5 | 66°30´–68°54´N | 54–200 | –0.9–0.3 | 400–540 | 740–880 | Pinus sylvestris | 62–70 | VT | 4–10 | 6 | Lipas et al. (1983) |
| 6 | 61°24´–67°39´N | 120 | –1.1–3.2 | 520–610 | 800–1190 | Pinus sylvestris, Picea abies | 103–180 | MT, VT | 10 | 32 | Lipas (1988) |
| 7 | 61°59´N | 180 | 3.6 | 714 | 1130 | Pinus sylvestris | 112 | VT | 5 | 4 | Levula (1991) |
| 8 | 63°19´–64°53´N | 60–180 | 0.3–5.5 | 590–600 | 910–1020 | Pinus sylvestris, Picea abies | 106 | MT, VT | 5 | 18 | Gustavsen & Lipas (1975) |
| 9 | 66°30´–67°24´N | 170 | –1.3–0.2 | 500–540 | 797–840 | Pinus sylvestris | 65–74 | MT, VT | 7 | 3 | Hirvelä & Hynynen (1990) |
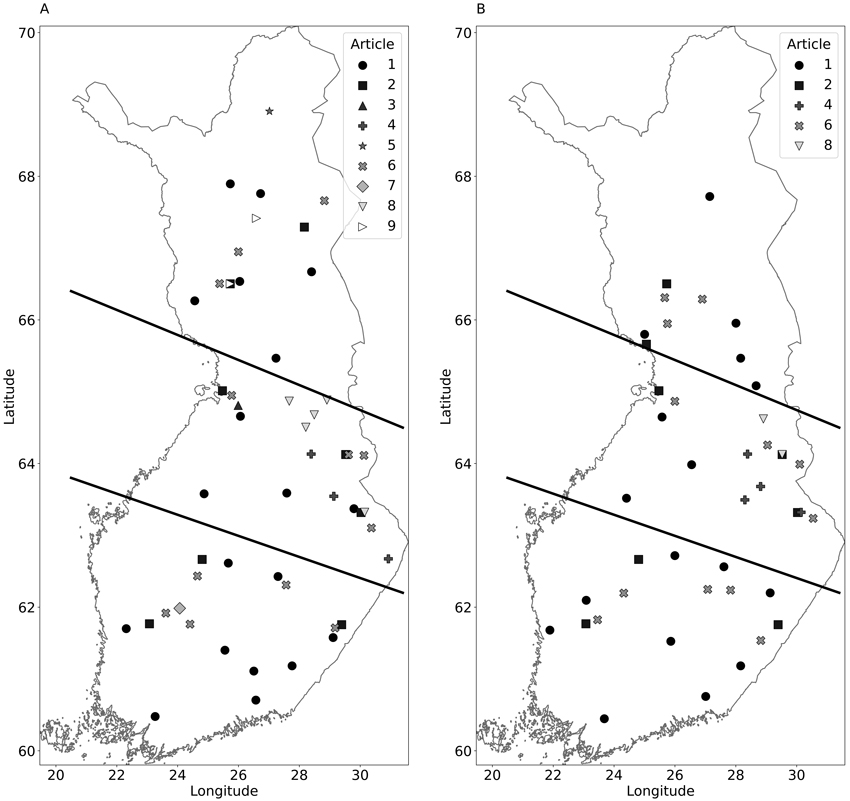
Fig. 1. Location of Scots pine sites (A) and Norway spruce sites (B) presented in the 9 reviewed articles (see Table 1 for references and article ids). The dataset contained 108 Scots pine observations and 57 Norway spruce observations from 97 different locations in Finland, covering a wide range of N dose, initial stand volumes, site types, and climatic conditions. Lines represent the grouping of observations based on location. The lines follow roughly the temperature sum, the northernmost part is temperature sum below 1000 d.d., middle is 1000–1200 d.d., and the southernmost is over 1200 d.d.
2.2 Statistical data analyses to quantify the magnitude of the volume growth response
After the volume growth was selected to be the studied growth response variable, we had to calculate the effect of fertilization and standardize it to enable comparisons between the studies. First, total volume growth response after the fertilization was calculated by subtracting the volume growth values of unfertilized control plots from the volume growth values of the fertilized plots and, where applicable, summed over the study period. Second, to enable comparison between studies with different durations, we used the mean annual growth response in our analyses. It was computed by dividing the total volume growth response with the time elapsed since the fertilization.
One-way ANOVA was used to analyze the differences in volume growth responses between geographical locations (Gsouth, Gmiddle, Gnorth, Fig. 1). The differences in volume growth responses between site types (Cajander 1949) were analyzed with nonparametric ANOVA using Kruskal–Wallis test and Wilcoxon rank sum test. All of the studies were conducted in Finland; hence they reported forest site types according to the classification of Cajander (1949), which is common forestry practice in Finland. The site type classification is based on the ground vegetation and the vegetation species composition has been shown to correlate with the site fertility (Nieppola and Carleton 1991). To obtain numerical classification of site fertility for the later statistical analyses, Cajander’s site types were combined into two groups which were coded as Boolean variable (Gfertility). The first group contained medium-fertile sites (Myrtillus site type, MT), and the other group contained all the medium-poor and poor sites (Vaccinium site type, VT and Calluna site type, CT) (Figs. 2 and 3).
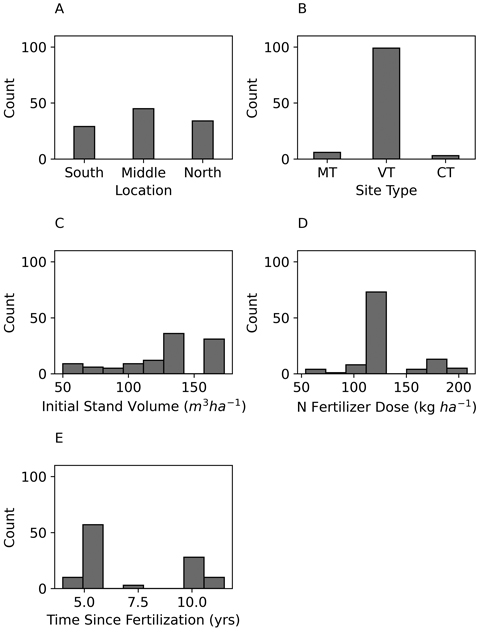
Fig. 2. Frequencies of Scots pine study site locations in Finland (A), site types according to Cajander (1949) (B), initial stand volumes (C), applied N fertilizer doses (D), and time since fertilization (E).
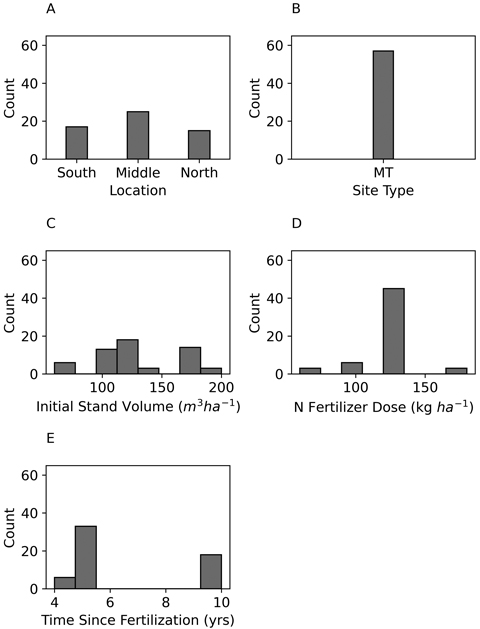
Fig. 3. Frequencies of Norway spruce study site locations in Finland (A), site types according to Cajander (1949) (B), initial stand volumes (C), applied N fertilizer doses (D), and time since fertilization (E).
The analyses were conducted separately for Scots pine and Norway spruce data. Long-term effective temperature sum (Tsum), long-term mean annual precipitation (Pa), site fertility (Gfertility), geographical location (Gsouth, Gmiddle, Gnorth, Glat), initial stand volume (Vini), time since fertilization (SF), and the applied N dose (FN), were used to explain the mean annual volume growth response in our analyses. Two different approaches were used to describe the geographical location: Latitude in decimal degree (Glat) and a dummy variable indicating whether the site was in Northern (Gnorth), Middle (Gmiddle), or Southern (Gsouth) Finland (Fig. 1). Initial stand volume (Vini) was the measured volume of the tree stand (m3 ha–1) before fertilization. Time since fertilization was also used as an explanatory variable because volume growth response is known to decrease gradually over time (Kukkola and Saramäki 1983). Linear mixed models were used to analyze the effects of different variables on the mean annual volume growth responses (Eq. 1).
![]()
where the dependent variable y is the mean annual volume growth response obtained by fertilization (m3 ha–1 yr–1), βT is a transpose of a coefficient vector (β1, β2, β3, … , β10) (see Table 2), X is a vector of explanatory fixed variables (see Table 2), Z is the vector of article id (Table 2) as random factors, and ε is the residual. The statistical analyses were performed with R 4.2.2 (R Core Team 2022). For the linear mixed effect models lme-function from nlme-package (Pinheiro and Bates 2024) was used.
| Table 2. Variables used in modeling the volume growth response of Scots pine and Norway spruce and the range of values in the dataset compiled from fertilization studies made in Finland. | ||||
| Variables | Coefficients | Description | Range of values in Scots pine data | Range of values in Norway spruce data |
| y | Annual mean volume growth response obtained by N fertilization (m3 ha–1 yr–1) | 0.18–2.7 | 0.36–2.3 | |
| FN | 1 | Applied dose of nitrogen (kg ha–1) | 54–208 | 59.8–180 |
| Pa | 2 | Mean annual precipitation (mm) | 396–714 | 472–630 |
| Gfertility | 3 | Site fertility group: Boolean, 1 when Site type ∈ [MT], else 0 | 0, 1 | 0, 1 |
| Tsum | 4 | Effective temperature sum (d.d.) | 743–1282 | 884–1283 |
| Gsouth | 5 | Area group: Boolean, 1 when in Southern Finland, else 0 | 0, 1 | 0, 1 |
| Gmiddle | 6 | Area group: Boolean, 1 when in Middle Finland, else 0 | 0, 1 | 0, 1 |
| Gnorth | 7 | Area group: Boolean, 1 when in Northern Finland, else 0 | 0, 1 | 0, 1 |
| Glat | 8 | Latitude of the stand in decimal degree | 61.76–68.91 | 61.76–66.50 |
| Vini | 9 | Initial stand volume (m3 ha–1) | 47–173 | 60–200 |
| SF | 10 | Time since fertilization | 4–15 | 4–15 |
| Aid | 11 | Identification number of the article where the data is from | 1–9 | 1, 2, 4, 8 |
First, we analyzed which are the best single variables (Table 2) explaining the variation in the volume growth response. Second, we kept the best explanatory variable, and inserted an additional variable to the model. Third, we kept adding explanatory variables, and studied how the goodness-of-fit of the models improved. Fourth, we started from a full vector of explanatory variables, and applied backward stepwise regression using dredge from package MuMIn (Bartoń 2024) to find the best combination of explanatory variables. Article identification number (Aid) was used as a random factor in all of the models. In both Scots pine and Norway spruce data, Tsum and Pa were highly correlated, and decreased towards the North. To avoid the multicollinearity, only one variable reflecting the climate and growing conditions of the site (Glat, Tsum and Pa) was included in a model (Supplementary file S1). Multicollinearity of the models was evaluated based on variance inflation factor. All of the models created in the steps from one to four were compared based on the Akaike Information Criteria (AIC), R2, RMSE, and p-values (Suppl. files S2, S3, S4). The best model for both Scots pine and Norway spruce, was selected based on the smallest AIC-values.
3 Results
3.1 Synthesis of the data
The data of both Scots pine and Norway spruce consisted of almost equal numbers of observations from Southern, Middle, and Northern Finland, however little more observations were from the Middle of Finland (Figs. 2 and 3). Most of the observations for Scots pine represented medium-poor fertile Vaccinium site type (VT), whereas all observations for Norway spruce represented medium-fertile Myrtillus site type (MT). The initial stand volume was mainly between 100 m3 and 200 m3 ha–1. Majority of the stands were fertilized with 120 kg N ha–1, and the duration of experiments was typically from 4 to 10 years (Figs. 2 and 3, Table 1).
3.2 Volume growth responses in different geographical locations and site types
The mean annual volume growth response did not differ between Southern, Middle, and Northern Finland for Scots pine and Norway spruce (Fig. 4). In medium-fertile (MT) Scots pine stands the volume growth response was significantly higher (p < 0.05) than in less fertile ones. For medium-poor fertility (VT) Scots pine stands the volume growth response was significantly higher (p < 0.05) than in poor fertility (CT) stands (Fig. 4). The interaction between site type and geographical location could not be reliably assessed because of the limited number of observations from different site types.
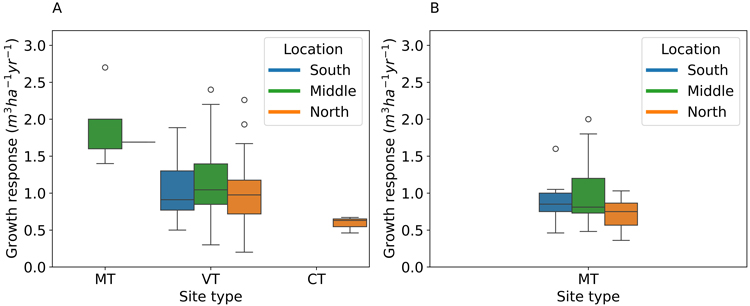
Fig. 4. The volume growth response of Scots pine (A) to N fertilization in different site types and geographical locations, and the volume growth response of Norway spruce (B) in different locations (data only available from medium-fertile (MT) sites). The site type classification is according to Cajander (1949), and all the sites are located in Finland.
3.3 Linear mixed models
The applied N dose (FN) was the best single variable explaining the magnitude of volume growth response both in Scots pine and Norway spruce stands (Table 3). In addition, mean annual precipitation (Pa) and time since fertilization (SF) were significant explanatory variables. For Scots pine, also the site fertility (Gfertility) explained the volume growth responses (Suppl. file S2). FN and Pa together explained 38% of the variation for Scots pine and 34% for Norway spruce (Suppl. file S3). For Scots pine, FN together with time since fertilization SF explained 38% of the variation of volume growth response and FN together with Gsouth explained 25%.
| Table 3. Comparison of alternative linear mixed models (Eq. 1) for volume growth responses to nitrogen fertilization in Scots pine and Norway spruce. The best model based on AIC-values (bold) and models with single explanatory variables (variables and their units are presented in Table 2). DF is degrees of freedom, AIC is Akaike’s Information Criteria, R2 is coefficient of determination of the fixed effects, and RMSE is residual mean square error. The models are ranked according to the AIC-value. | ||||||
| Tree species | Independent predictors included in Eq. 1 | DF | AIC | R2 | RMSE | p-value |
| Scots pine | FN + Pa + Gfertility + SF | 107 | 73.3 | 0.547 | 0.318 | <0.001 |
| FN | 107 | 105.7 | 0.314 | 0.360 | <0.001 | |
| Gfertility | 107 | 124.2 | 0.090 | 0.376 | 0.003 | |
| Pa | 107 | 127.3 | 0.050 | 0.378 | 0.016 | |
| SF | 107 | 127.6 | 0.178 | 0.368 | 0.016 | |
| Norway spruce | FN + Pa + SF | 56 | 2.7 | 0.510 | 0.204 | <0.001 |
| FN | 56 | 9.7 | 0.240 | 0.210 | <0.001 | |
| Pa | 56 | 33.1 | 0.101 | 0.272 | 0.041 | |
| SF | 56 | 34.5 | 0.157 | 0.278 | 0.169 | |
For Scots pine the best combination of explanatory variables were FN, Pa, Gfertility, and SF according to AIC-values (Suppl. file S4: Table S4.1). These fixed effect variables explained together 55% of the variation in the Scots pine volume growth response (Table 3). The best model for Norway spruce included FN, Pa, and SF, and the fixed effects explained 51% of the variation (Table 3). The article identification number as a random effect increased the goodness of fit of the Norway spruce model but did not have a significant effect on the Scots pine model.
The volume growth response increased with increasing FN and Pa (Table 4). The increase in SF decreased the annual mean volume growth response (Table 4). On medium-fertile sites, the Scots pine volume growth response was larger than on less fertile sites (Table 4, Fig. 5).
| Table 4. Parameter values of the fixed and random effects of the best linear mixed models for Scots pine and Norway spruce volume growth responses. The volume growth response is in units of m3 ha–1 yr–1 and explanatory variables, and their units are presented in Table 2. | ||||||
| Scots pine | Norway spruce | |||||
| Fixed effects | β | SE | p-value | β | SE | p-value |
| Intercept | –0.271 | 0.319 | 0.399 | –1.254 | 0.598 | 0.041 |
| FN (kg N ha–1) | 0.009 | 0.001 | <0.001 | 0.009 | 0.001 | <0.001 |
| Pa (mm) | 0.001 | 0.001 | 0.022 | 0.003 | 0.001 | 0.006 |
| Gfertility | 0.619 | 0.155 | <0.001 | |||
| SF (yrs.) | –0.071 | 0.013 | <0.001 | –0.059 | 0.023 | 0.086 |
| Random effects | Variance | SD | Variance | SD | ||
| Aid | <0.001 | <0.001 | 0.012 | 0.107 | ||
| Residual | 0.101 | 0.318 | 0.044 | 0.211 | ||

Fig. 5. Mean annual measured (dots) and modeled (lines, the best models according to Table 4) volume growth responses as a function of applied N fertilizer dose for different annual precipitation (Pa), site fertility level (Gfertility = 1 includes MT, and Gfertility = 0 includes VT and CT, Cajander (1949)). Pa values represent conditions in Northern (red and orange lines) and Southern (blue and green lines) Finland while time since fertilization (SF) is set to 10 years for both tree species. Panel A shows Scots pine and B the Norway spruce volume growth responses.
4 Discussion
The effects of N fertilization on volume growth, wood production, and its economic profitability in different growing conditions have been investigated in previous studies, and the results have been discussed in literature reviews by Saarsalmi and Mälkönen (2001) and Lehto and Ilvesniemi (2023). However, there has not yet been quantitative synthesis on growth responses of Scots pine and Norway spruce stands based on these earlier N fertilization studies in Finland. Compiling such quantitative information is needed to support the decision making in practical forestry, especially for the use of sustainable precision fertilization schemes to promote rapid carbon sequestration.
Combining data from 9 published fertilization studies for Scots pine and Norway spruce across Finland allowed us to analyze the effects of N dose, climatic conditions, and stand characteristics on the mean annual volume growth responses induced by N fertilization. Only Kukkola and Saramäki (1983) have earlier built predictive models for the volume growth responses after N fertilization based on fertilization experiments established in 1955–1965 in Southern Finland. After that, many new fertilization experiments have been established and results published. In our dataset, all of the included articles reported results from experiments that were fertilized after 1965. We also filled in the missing weather data from the historic records (Finnish Meteorological Institute 2024) to consider the effect of climatic conditions across the south-north gradient of Finland.
We found that N fertilizer dose, mean annual precipitation, site fertility, and time since N fertilization affected the mean annual volume growth responses. The growth response to N fertilization is primarily induced by the increase in needle N-concentration, which enhances leaf-level photosynthetic rate (Kellomäki and Wang 1997a, 1997b). As predicted, in our models, the volume growth response increased linearly with respect to N dose, but it should be noted that the volume growth response may saturate at high N doses. This is due to the fact that N fertilization increases the needle mass and leaf area index (Choi et al. 2005), which eventually starts to increase the shading in the canopies (Hyvönen et al. 2007). Higher needle mass increases the interception, evaporation, and transpiration, which may decrease soil water content (Linder et al. 1987; Bergh et al. 2005; Launiainen et al. 2016), and therefore may limit the volume growth response.
A N fertilizer dose of 120 kg ha–1 is overrepresented in the dataset because three of the studies included (Lipas and Levula 1980; Laakkonen et al. 1983; Lipas 1988) had a high number of extracted observations for stands fertilized with 120 kg N ha–1, and the number of the extracted observations was high (Table 1). All these three experiments were started in 1968 or 1969, and according to Laakkonen et al. (1983) the 120 kg N ha–1 was the upper limit of the recommended fertilizer dose at the time. For Scots pine this was not a problem as there were a sufficient number of other N doses in the data, but for Norway spruce this may cause some uncertainty when using the model with lower or higher N doses than 120 kg N ha–1. We performed statistical analyses to investigate which variables explained the variation in the volume growth responses in only the 120 kg N ha–1 fertilized studies, and the results remained the same as with the whole dataset.
We observed that climatic conditions affected the volume growth responses. The mean annual volume growth response increased with increasing mean annual precipitation. Higher precipitation and water availability have also been found to increase the growth response to N fertilization in previous studies (Lim et al. 2015). On the other hand, it should be noted that mean annual precipitation and the effective temperature sum were highest in southern Finland, and decreased towards the north in our dataset, and they correlated with latitude (Suppl. file S1). Due to this connection between mean annual precipitation and location, precipitation as an explanatory variable in our models may be a proxy for the geographical location of the site rather than reflect the effect of site water availability per se. Models containing effective temperature sum and latitude as an explanatory variable were also statistically significant (p < 0.05), but when comparing AIC values, the model with mean annual precipitation was chosen as the best model (Suppl. files S4, S5).
The site fertility also created variation in the volume growth responses as expected; the volume growth responses were higher in medium-fertile than in less fertile sites for Scots pine. This was supported by the previous study of Pettersson (1994), who found that the relative growth response for Scots pine was higher in fertile sites when N dose was below 150 kg ha–1. However, with the higher N fertilizer doses, it is possible that following the Liebig’s law of the minimum (Fabrika and Pretzsch 2013), the availability of other nutrients, for example phosphorus, boron, or potassium, may start to limit the growth response (Gustavsen and Lipas 1975; Tamm 1991). Nutrient poor sites are generally located on coarse-textured soils, and therefore water deficiency can also limit the growth response (Kellomäki 2022a). For Norway spruce, all the experiments in our dataset were conducted on medium-fertile sites, and therefore, a similar kind of comparison was not possible.
To identify the effect of the stand development phase on the growth response we used the initial stand volume, because it was most frequently reported in the synthesized studies. Higher initial volume predicted lower volume growth response for both Scots pine and higher volume growth response for Norway spruce, however the impact was not large enough for the variable to be included in the best models. Based on the previous studies, the stand development stage affects nutrient demand, and the N accumulation to the tree biomass is fastest in the early phases of stand development (Palviainen and Finér 2012). Tree size and the competitive position in the stand also affects the growth response; absolute volume growth is highest in dominant trees (Kukkola 1978). But the growth response tends to decrease with stand age (Gustavsen and Lipas 1975). The created models can only be applied in the stands where initial volume is over 50 m3 ha–1 because initial stand volumes lower than that were not included in the analyses.
There was less variation in the applied amounts of N in Norway spruce data than in Scots pine data (Figs. 2d and 3d). The volume growth responses for Norway spruce were also lower than for Scots pine. Also, previous studies have shown higher growth responses to N fertilization in Scots pine stands than in Norway spruce stands (Moilanen and Meriluoto 1984; From et al. 2016). The differences between Scots pine and Norway spruce can be explained by different nutrient requirements (Ingestad 1979; Mälkönen 1979), and initial nutrient balance of the site, as Scots pines typically grow in less fertile, N-limited sites (Kalliola 1973; Kellomäki 2022a). Norway spruce also needs more N to produce the same stand volume than Scots pine, because Norway spruce has higher needle mass and needle N concentrations (Palviainen and Finér 2012).
According to Kukkola and Saramäki (1983), the volume growth response for Norway spruce is longer-lasting than for Scots pine, and higher N fertilizer doses cause a longer lasting growth response, than lower dosages (Kellomäki 2022b). The peak of growth response comes also later and lasts longer for Norway spruce than for Scots pine. The delayed impact is related to the photosynthesizing foliage biomass, which consists of 6–12 needle age cohorts for Norway spruce, and only 3–5 needle age cohorts for Scots pine (Jalkanen et al. 1995; Niinemets and Lukjanova 2003). Improved N availability starts to increase photosynthesis gradually with the renewal of needle cohorts (Kukkola and Saramäki 1983; Kellomäki 2022b). The best models for both Scots pine and Norway spruce contained time since fertilization, and the mean annual volume growth response decreased with the time elapsed from fertilization as predicted. This aligns with the results of previous studies that have reported the growth response to decrease after six years and ceases after ten years (Kukkola and Saramäki 1983; Pettersson 1994).
There might have been some additional factors causing variation in the volume growth responses as the data were collected from various experiment designs. We used the articles as a random effect in the models to capture the variation caused by different methodologies. Articles included in the present study applied different ways to calculate the stand volume, which may affect the results. Variation in the volume growth responses might have been affected by the use of different fertilizers (Kukkola and Saramäki 1983). The growth response to urea is known to be weaker than to ammonium nitrate because urea evaporates easily (Laakkonen et al. 1983). One additional source of variation in the dataset might be due to different forest management history in the study sites. Thinning together with N fertilization induces a larger growth response than N-fertilization alone (Haapanen et al. 1979), because thinning also changes availability of other resources, such as light and water. These additional variations had more impact on the Norway spruce data due to lower number of observations in the dataset.
N fertilization increases wood production in boreal forests, and therefore it also increases the rate of carbon sequestration (Jörgensen et al. 2021). Thus, N fertilization can be used as a tool for climate change mitigation. However, N fertilization may lead to nutrient leaching to groundwater, nutrient export to water courses, and changes in vegetation species composition (Binkley et al. 1999; Hedwall 2014; Jetsonen et al. 2024). Deciding the optimum N fertilizer dose needs balancing between the anticipated volume growth response, economic feasibility, and avoiding the adverse environmental effects. The results of this study can be used to support planning environmentally responsible and economically viable precision fertilization, as they can provide quantitative information about volume growth responses to different N fertilization doses in different conditions. The results can be used to calculate the climate change mitigation potential and economic feasibility of fertilized Scots pine and Norway spruce stands, and to better understand the pathways of N fertilizer in the forest ecosystem. Our results suggest that in fertilization management the optimal N dose depends on the geographical location, initial stand fertility and tree species. More research is needed on the growth responses using higher N doses especially from Norway spruce stands and from the most fertile sites (Oxalis-maianthemum (OMaT) and Oxalis-myrtillus (OMT) site types) which were completely lacking from the literature.
Data and code availability
Data and code are openly available at the Zenodo data repository: https://doi.org/10.5281/zenodo.14733005.
Authors’ contributions
JJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. AL: Conceptualization, Investigation, Methodology, Writing – review & editing. HP: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – review & editing. KL: Data curation, Investigation, Visualization. SL: Conceptualization, Funding acquisition, Methodology, Writing – review & editing. MP: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – review & editing.
Funding
This research was funded by the PREFER-project (decision numbers 348096, 348102 and 348103) and UNITE flagship (decision number 357906), both funded by the Research Council of Finland.
References
Äijälä O, Koistinen A, Sved J, Vanhatalo K, Väisänen P (eds) (2019) Metsänhoidon suositukset. [Forest management recommendations]. Tapion julkaisuja.
Bartoń K (2024) MuMIn: multi-model inference. R package version 1.48.4. https://CRAN.R-project.org/package=MuMIn.
Bergh J, Linder S, Bergström J (2005) Potential production of Norway spruce in Sweden. Forest Ecol Manag 104: 1–10. https://doi.org/10.1016/j.foreco.2004.07.075.
Binkley D, Burnham H, Allen H (1999) Water quality impacts of forest fertilization with nitrogen and phosphorus. Forest Ecol Manag 121: 191–213. https://doi.org/10.1016/S0378-1127(98)00549-0.
Bradshaw CJ, Warkentin IG (2015) Global estimates of boreal forest carbon stocks and flux. Glob Planet Chang 128: 24–30. https://doi.org/10.1016/j.gloplacha.2015.02.004.
Cajander AK (1949) Forest types and their significance. Acta For Fenn 56, article id 7396. https://doi.org/10.14214/aff.7396.
Choi W-J, Chang SX, Allen HL, Kelting DL, Ro H-M (2005) Irrigation and fertilization effects on foliar and soil carbon and nitrogen isotope ratios in a loblolly pine stand. Forest Ecol Manag 213: 90–101. https://doi.org/10.1016/j.foreco.2005.03.016.
Fabrika M, Pretzsch H (2013) Forest ecosystem analysis and modelling. Technical University in Zvolen.
Finnish Meteorological Institute (2024) The Finnish Meteorological Institute’s open data. https://en.ilmatieteenlaitos.fi/open-data. Accessed 6 February 2024.
From F, Lundmark T, Mörling T, Pommerening A, Nordin A (2016) Effects of simulated long-term N deposition on Picea abies and Pinus sylvestris growth in boreal forest. Can J Forest Res 46: 1396–1403. https://doi.org/10.1139/cjfr-2016-0201.
Gustavsen H, Lipas E (1975) Lannoituksella saatavan kasvunlisäyksen riippuvuus annetusta typpimäärästä. [Effect of nitrogen dosage on fertilizer response]. Folia For 246. http://urn.fi/URN:ISBN:951-40-0188-5.
Haapanen T, Hari P, Kellomäki S (1979) Effect of fertilization and thinning on radial growth of Scots pine. Silva Fenn 13: 184–189. https://doi.org/10.14214/sf.a14890.
Hedwall PO, Gong P, Ingerslev M, Bergh J (2014) Fertilization in northern forests – biological, economic and environmental constraints and possibilities. Scand J Forest Res 29: 301–311. https://doi.org/10.1080/02827581.2014.926096.
Hirvelä H, Hynynen J (1990) Lannoituksen vaikutus männikön kasvuun, latvavaurioihin ja tuulituhoalttiuteen Lapissa. [Effect of fertilization on the growth, top damage and susceptibility to windthrow of Scots pine stands in Lapland]. Folia For 764. http://urn.fi/URN:ISBN:951-40-1133-3.
Hyvönen R, Ågren GI, Linder S, Persson T, Cotrufo MF, Ekblad A, Freeman M, Grelle A, Janssens IA, Jarvis PG, Kellomäki S, Lindroth L, Loustau D, Lundmark T, Norby RJ, Oren R, Pilegaard K, Ryan MG, Sigurdsson BD, Strömgren M, van Oijen M, Wallin G (2007) The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: a literature review. New Phytol 173: 463–480. https://doi.org/10.1111/j.1469-8137.2007.01967.x.
Ilvesniemi H, Forsius M, Finér L, Holmberg M, Kareinen T, Lepistö A, Piirainen S, Pumpanen J, Rankinen K, Starr M, Tamminen P, Ukonmaanaho L, Vanhala P (2002) Carbon and nitrogen storages and fluxes in Finnish forest ecosystems. In: Käyhkö J, Talve L (eds) Understanding the global system. The Finnish perspective. Finnish Global Change Programme FIGARE, pp 69–82. https://www.researchgate.net/publication/259287067_Understanding_the_global_system_-_the_Finnish_perspective_FIGARE_Scientific_Report.
Ingestad T (1979) Mineral nutrient requirements of Pinus silvestris and Picea abies seedlings. Physiol Plantarum 45: 373–380. https://doi.org/10.1111/j.1399-3054.1979.tb02599.x.
Jahns T, Kaltwasser H (2000) Mechanism of microbial degradation of slow-release fertilizers. J Polym Environ 8: 11–16. https://doi.org/10.1023/A:1010116027139.
Jahns T, Ewen H, Kaltwasser H (2003) Biodegradability of urea-aldehyde condensation products. J Polym Environ 11: 155–159. https://doi.org/10.1023/A:1026052314695.
Jalkanen R, Aalto T, Kurkela T (1995) Development of needle retention in Scots pine (Pinus sylvestris) in 1957–1991 in northern and southern Finland. Trees 10: 125–133. https://doi.org/10.1007/BF00192193.
Jetsonen J, Laurén A, Peltola H, Muhonen O, Nevalainen J, Ikonen V-P, Kilpeläinen A, Tuittila E-S, Männistö E, Kokkonen N, Palviainen M (2024) Effects of nitrogen fertilization on the ground vegetation cover and soil chemical properties in Scots pine and Norway spruce stands. Silva Fenn 58, article id 23058. https://doi.org/10.14214/sf.23058.
Jörgensen K, Granath G, Lindahl BD, Strengbom J (2021) Forest management to increase carbon sequestration in boreal Pinus sylvestris forests. Plant Soil 466: 165–178. https://doi.org/10.1007/s11104-021-05038-0.
Kalliola R (1973) Suomen kasvimaantiede. [Finnish plant geography]. WSOY, Porvoo. ISBN 951-0-05731-2.
Kellomäki S (2022a) Environmental conditions, site types, and climate change. In: Kellomäki S (ed) Management of boreal forests: theories and applications for ecosystem services. Springer, pp 13–62. https://doi.org/10.1007/978-3-030-88024-8_2.
Kellomäki S (2022b) Fertilization in management of site fertility. In: Kellomäki S (ed) Management of boreal forests: theories and applications for ecosystem services. Springer, pp 437–464. https://doi.org/10.1007/978-3-030-88024-8_14.
Kellomäki S, Wang K-Y (1997a) Effects of long-term CO2 and temperature elevation on crown nitrogen distribution and daily photosynthetic performance of Scots pine. For Ecol Manag 99: 309–326. https://doi.org/10.1016/S0378-1127(97)00059-5.
Kellomäki S, Wang K-Y (1997b) Photosynthetic responses of Scots pine to elevated CO2 and nitrogen supply: results of a branch-in-bag experiment. Tree Physiol 17: 231–240. https://doi.org/10.1093/treephys/17.4.231.
Kellomäki S, Puttonen P, Tamminen H, Westman CJ (1982) Effect of nitrogen fertilization on photosynthesis and growth in young scots pine. Preliminary results. Silva Fenn 16: 363–371. https://doi.org/10.14214/sf.a15084.
Kukkola M (1978) Lannoituksen vaikutus eri latvuskerrosten puiden kasvuun mustikkatyypin kuusikossa. [Effect of fertilization on the growth of different tree classes in a Norway spruce stand on Myrtillus-site]. Folia For 362. http://urn.fi/URN:ISBN:951-40-0353-5.
Kukkola M, Saramäki J (1983) Growth response in repeatedly fertilized pine and spruce stands on mineral soils. Commun Inst For Fenn 114. http://urn.fi/URN:ISBN:951-40-0622-4.
Laakkonen O, Keipi K, Lipas E (1983) Typpilannoituksen kannattavuus varttuneissa kangasmetsissä. [Profitability of nitrogen fertilization in mature forests on mineral soils]. Folia For 577. http://urn.fi/URN:ISBN:951-40-0645-3.
Launiainen S, Katul GG, Kolari P, Lindroth A, Lohila A, Aurela M, Varlagin A, Grelle A, Vesala T (2016) Do the energy fluxes and surface conductance of boreal coniferous forests in Europe scale with leaf area? Global Change Biol 22: 4096–4113. https://doi.org/10.1111/gcb.13497.
Lehto T, Ilvesniemi H (2023) Metsänlannoitus nyt ja tulevaisuudessa: synteesiraportti. [Forest fertilization now and in the future: synthesis]. Luonnonvara- ja biotalouden tutkimus 56/2023. http://urn.fi/URN:ISBN:978-952-380-714-3.
Levula T (1991) Tuhkalannoitus kangasmailla. [Ash fertilization on the mineral soils]. Finnish Forest Research Institute Research Papers 394: 49–59. http://urn.fi/URN:ISBN:951-40-1180-5.
Lim H, Oren R, Palmroth S, Tor-ngern P, Mörling T, Näsholm T, Lundmark T, Helmisaari H-S, Leppälammi-Kujansuu J, Linder S (2015) Inter-annual variability of precipitation constrains the production response of boreal Pinus sylvestris to nitrogen fertilization. Forest Ecol Manag 348: 31–45. https://doi.org/10.1016/j.foreco.2015.03.029.
Linder S, Benson ML, Myers BJ, Raison RJ (1987) Canopy dynamics and growth of Pinus radiata.: I. Effects of irrigation and fertilization during a drought. Can J Forest Res 17: 1157–1165. https://doi.org/10.1139/x87-179.
Lipas E (1988) Typpilannoituksen ajankohta kangasmetsissä. [Timing of nitrogen fertilization on the mineral soils]. Folia For 709. http://urn.fi/URN:ISBN:951-40-0805-7.
Lipas E, Levula T (1980) Urealannoitus eri vuodenaikoina. [Urea fertilization at different times of the year]. Folia For 421. http://urn.fi/URN:ISBN:951-40-0432-9.
Lipas E, Levula T, Välikangas P (1983) Eräitä metsänlannoitustuloksia Lapista. [Some forest fertilization results from Lapland]. Finnish Forest Research Institute Research Papers 114. http://urn.fi/URN:ISBN:951-40-0997-5.
Mäkipää R (1995) Effect of nitrogen input on carbon accumulation of boreal forest soils and ground vegetation. Forest Ecol Manag 79: 217–226. https://doi.org/10.1016/0378-1127(95)03601-6.
Mälkönen E (1979) Kangasmaiden lannoitustutkimus. [Research on forest fertilization on mineral soils]. Folia For 400: 20–29. http://urn.fi/URN:ISBN:951-40-0403-5.
Moilanen M, Meriluoto M (1984) Muhos-operaatio – lannoitustutkimusta yksityismetsissä. [Muhos-operation – fertilization research in privately owned forests]. Metsäntutkimuslaitos. http://urn.fi/URN:NBN:fi-metla-201211127146.
Nieppola JJ, Carleton T J (1991) Relations between understorey vegetation, site productivity, and environmental factors in Pinus sylvestris L. stands in southern Finland. Vegetatio 93: 57–72. https://doi.org/10.1007/BF00044924.
Niinemets Ü, Lukjanova A (2003) Total foliar area and average leaf age may be more strongly associated with branching frequency than with leaf longevity in temperate conifers. New Phytol 158: 75–89. https://doi.org/10.1046/j.1469-8137.2003.00712.x.
Palviainen M, Finér L (2012) Estimation of nutrient removals in stem-only and whole-tree harvesting of Scots pine, Norway spruce, and birch stands with generalized nutrient equations. Eur J Forest Res 131: 945–964. https://doi.org/10.1007/s10342-011-0567-4.
Pettersson F (1994) Predictive functions for impact of nitrogen fertilization on growth over five years. Report No 3, The Forestry Research Institute of Sweden, Uppsala.
Philben M, Ziegler SE, Edwards KA, Kahler III R, Benner R (2016) Soil organic nitrogen cycling increases with temperature and precipitation along a boreal forest latitudinal transect. Biogeochemistry 127: 397–410. https://doi.org/10.1007/s10533-016-0187-7.
Pinheiro J, Bates D, R Core Team (2024). nlme: linear and nonlinear mixed effects models. R package version 3.1-166. https://CRAN.R-project.org/package=nlme.
PlotDigitizer (2023) Version 3.1.5. https://plotdigitizer.com/app.
Pukkala T (2017) Optimal nitrogen fertilization of boreal conifer forest. For Ecosyst 4, article id 3. https://doi.org/10.1186/s40663-017-0090-2.
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Saarsalmi A, Mälkönen E (2001) Forest fertilization research in Finland: a literature review. Scand J Forest Res 16: 514–535. https://doi.org/10.1080/02827580152699358.
Salonen K (1973) Eri vuodenaikoina annetun typpilannoituksen vaikutuksesta kangasmetsissä. [On the response of mineral-soil forests to nitrogen application during different seasons of the year]. Suo – Mires and peat 24: 99–105. http://suo.fi/article/9417.
Schlosser RW, Wendt O, Bhavnani S, Nail-Chiwetalu B (2006) Use of information-seeking strategies for developing systematic reviews and engaging in evidence-based practice: the application of traditional and comprehensive Pearl Growing. A review. Int J Lang Comm Dis 41: 567–582. https://doi.org/10.1080/13682820600742190.
Sikström U, Nohrstedt HÖ, Pettersson F, Jacobson S (1998) Stem-growth response of Pinus sylvestris and Picea abies to nitrogen fertilization as related to needle nitrogen concentration. Trees 12: 208–214. https://doi.org/10.1007/PL00009712.
Smolander A, Martikainen P, Henttonen H (2022) Half-a-century effects of a slow-release nitrogen fertilizer, ureaformaldehyde, on stand growth and soil processes in a Scots pine stand. Forest Ecol Manag 519, article id 120320. https://doi.org/10.1016/j.foreco.2022.120320.
Tamm CO (1991) Nitrogen in terrestrial ecosystems. Questions of productivity, vegetational changes, and ecosystem stability. Springer, Berlin. ISBN 9783540518075.
Valinger E, Elfving B, Mörling T (2000) Twelve-year growth response of Scots pine to thinning and nitrogen fertilisation. Forest Ecol Manag 134: 45–53. https://doi.org/10.1016/S0378-1127(99)00244-3.
Total of 58 references.
