Annija Kārkliņa  ,
Guntis Brūmelis,
Iluta Dauškane,
Didzis Elferts,
Lāsma Freimane,
Māra Kitenberga,
Zane Lībiete,
Roberts Matisons,
Āris Jansons
,
Guntis Brūmelis,
Iluta Dauškane,
Didzis Elferts,
Lāsma Freimane,
Māra Kitenberga,
Zane Lībiete,
Roberts Matisons,
Āris Jansons
Effect of salvage-logging on post-fire tree establishment and ground cover vegetation in semi-natural hemiboreal forests
Kārkliņa A., Brūmelis G., Dauškane I., Elferts D., Freimane L., Kitenberga M., Lībiete Z., Matisons R., Jansons Ā. (2020). Effect of salvage-logging on post-fire tree establishment and ground cover vegetation in semi-natural hemiboreal forests. Silva Fennica vol. 54 no. 5 article id 10334. https://doi.org/10.14214/sf.10334
Highlights
- Effect of salvage logging on post-fire understory vegetation was assessed
- Effect of salvage logging differed depending on forest types
- In dry-poor stands, Calluna vulgaris was hindering other plant species
- In wet stands, logging had positive effect on understory vegetation diversity
- Salvage logging enhanced the effect of natural disturbance in dry-rich stands.
Abstract
Fire is a common disturbance in boreal forests causing changes in biological diversity at various spatial scales. In the past 100 years, forest management has limited fire outbreaks, but in the future, the fire-affected forest area is expected to increase in many regions due to climate change. Burned forests are typically salvage-logged, but the effect of this type of management versus natural regeneration on biological diversity is not well understood, particularly the mid-term effect to tree establishment and understory vegetation composition and diversity. Various management methods were used after a large fire in 1992 in a peatland-forest complex and neighbouring managed forests, which created an experimental setup for study of the effect of management after fire in the Sliteres National park, northwestern Latvia. Understory vegetation was described in plots using a design of four forest and three management types: natural regeneration (unmanaged) and managed sites with salvage logging followed by no further human intervention and salvage logging with planting. Post-fire management had different effect in each forest type. Species richness was higher in forest types with salvage logging than in natural regenerated sites on rich wet and rich dry forest types, but not for the poor forest types. Tree regeneration was generally greater in salvage-logged stands, but differed between forest types. Species composition was related to tree regeneration and canopy openness. In contrast to other studies, salvage logging had a positive mid-term effect to ground vegetation diversity and tree establishment in the studied stands, implying potential for concomitant management and conservation of ground cover vegetation in semi-natural stands.
Keywords
management;
forest fire;
natural disturbance;
legacies;
understory vegetation
-
Kārkliņa,
Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia
E-mail
annija.karklina@silava.lv
- Brūmelis, University of Latvia, Faculty of Biology, Jelgavas street 1, LV-1004, Riga, Latvia E-mail guntis.brumelis@lu.lv
- Dauškane, University of Latvia, Faculty of Biology, Jelgavas street 1, LV-1004, Riga, Latvia E-mail iluta.dauskane@lu.lv
- Elferts, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia; University of Latvia, Faculty of Biology, Jelgavas street 1, LV-1004, Riga, Latvia E-mail didzis.elferts@lu.lv
- Freimane, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia E-mail lasma.freimane@silava.lv
- Kitenberga, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia E-mail mara.kitenberga@silava.lv
- Lībiete, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia E-mail zane.libiete@silava.lv
- Matisons, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia E-mail robism@inbox.lv
- Jansons, Latvian State Forest Research Institute “Silava”, 111 Rigas Street, LV-2169, Salaspils, Latvia E-mail aris.jansons@silava.lv
Received 2 March 2020 Accepted 5 October 2020 Published 12 October 2020
Views 87535
Available at https://doi.org/10.14214/sf.10334 | Download PDF

Supplementary Files
1 Introduction
Fire is a disturbance of natural and human origin, which has shaped the structure and composition of boreal forest at various spatial scales (Rowe and Scotter 1973; Parviainen 1996). Changes in forest management and the end of slash-and-burn agriculture have minimized impact of fire on forest development and understory vegetation (Wallenius 2011). However, the variability in fire frequency is associated with climate and therefore is expected to be sensitive to global change (Drobyshev et al. 2014; Donis et al. 2017; Kitenberga et al. 2019).
Natural disturbances such as fire, windthrow and insect attack create new conditions for the understory vegetation. Fire causes variable tree mortality and affects understory vegetation, which play crucial roles in ecosystem processes, influencing succession, nutrient and carbon cycles and long-term productivity (Nilsson and Wardle 2005; Hart and Chen 2008). Fire also affects soil quality by nutrient release, and depending on fire severity, it can result in mineralization of part or all of the upper litter layer, damaging roots and seeds, and creating new regeneration niches for species (Parro et al. 2009). Plants have various adaptations to fire, which determine the trajectories of ecological succession of the understory after fires of variable intensity and severity (Rowe 1983). Plant strategies related to fire involve morphological and physiological traits for protecting plant tissues and improving survival during the fire (Buhk et al. 2007). For example, perennial plants are protected from complete destruction by belowground organs that are able to produce new sprouts, or they regenerate from seeds (Trabaud 1987).
Understory vegetation composition depends on past forest disturbance, soil conditions and canopy composition (Kuuluvainen 1994). After fire, forest and ground cover vegetation changes in structure are associated with soil moisture and temperature, organic matter accumulation and tree stand regeneration (Bonan and Shugart 1989; Wallenius et al. 2004). In dry forests, vegetation can be completely destroyed by fire and are typically more affected than in wet forest types (Gromtsev 2002).
Salvage logging after fire is a forest management method that is often used to procure wood and to facilitate site preparation and planting (Thorn et al. 2016). Species diversity and composition of understory plant communities that develop after salvage logging can differ from those after natural succession after fire, depending on site conditions (Lindenmayer and Noss 2006). Still, controversial opinions about the effects of this management are presented, though information for hemiboreal forests is scarce. Studies about the impact of salvage logging effect differ in the period after fire studied, and also, compared with natural regeneration without tree removal, the studies show either negative impact on regeneration (Lindenmayer and Ought 2006; Lindenmayer et al. 2009), no effect, or even better regeneration (Peterson and Dodson 2016). The contrasting results may be attributed to different patterns of regeneration depending on forest type, soil, time after fire, and the size of the burnt area (Kurulok and Macdonald 2007).
In the hemiboreal zone, studies have shown that wildfires were frequent in the last 250 years (Kitenberga et al. 2019). The last major fire in Latvia covered an area of 3300 ha, mostly within the Slitere National park (Kitenberga et al. 2019). After the fire, salvage logging with and without planting was carried out in different forest types, which allows to test for differences in understory vegetation patterns depending on management type interacting with soil conditions. The aim of the study was to determine effect of management after fire (natural regeneration, salvage logging without planting, and salvage logging with planting) on tree regeneration and understory plant community composition and species richness 25 years after the disturbance. We hypothesized that the combined effect of fire and salvage logging would lead to reduced species richness and altered community composition. We also expected that the effects would be more pronounced in richer forest types as a result of more suitable conditions for establishment of trees and herbs.
2 Material and methods
2.1 Study area
The peatland-forest complex of Sliteres Nacional Park (SNP) is located in the north-west part of Latvia (57°68´–57°70´N, 22°46´–22°52´E) (Fig. 1). Latvia lies in the hemiboreal vegetation zone (Nilsson 1997), where climate conditions are influenced by air masses from the Baltic Sea and Atlantic Ocean (Kitenberga et al. 2019). According to long-term data from the Latvian Environment, Geology and Meteorology Centre, mean annual temperature in the study area is +6.9 °C (–3.9 °C in February and +16.9 °C in July). The annual sum of precipitation varies from 568 to 606 mm.
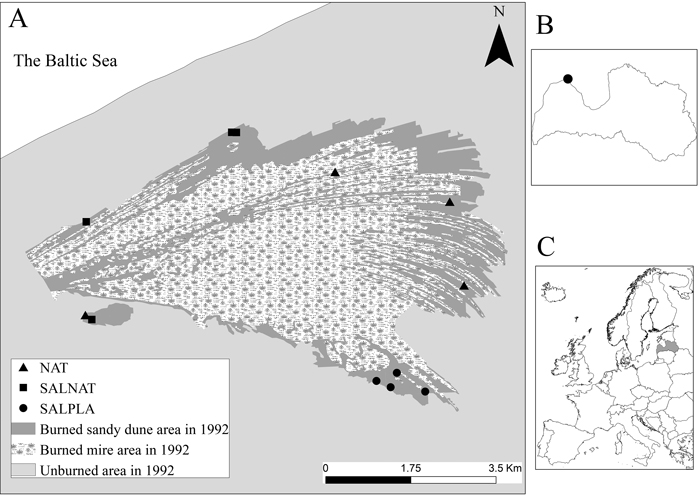
Fig. 1. A. Location of the studied forest stands in Sliteres National park. B. The study site location within Latvia. C. Location of Latvia in Europe. NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting.
The study was carried out in an area within the Slitere National park, where a large fire occurred in 1992. Of the total burnt area (3300 ha), 1022 ha was forest and the remaining area was a mire (Kitenberga et al. 2019). Three types of post-fire management were applied: 1) salvage logging on 34 ha within the national park, later left for natural regeneration (SALNAT); 2) salvage logging outside of the national park on 108 ha with planting of Pinus sylvestris L. in 1993 (SALPLA), and 3) natural regeneration, which included the largest area of the park, with no further management of the stands (NAT) (Fig. 1). The burnt area is a dune and bog complex with dry stands on sandy soils divided by dune slacks with wet forest stands.
2.2 Data sampling
Data were collected in four forest types: two dry in very poor (Cladinoso-callunosa) and slightly richer sandy soils (Vacciniosa) and two in wet-mineral soils of low fertility (Myrtilloso-sphagnosa) and peat soils of medium fertility (Sphagnosa). The study design was three management types × 4 forest types. Due to the small area that was salvage logged and not planted (within the strict reserve), only one stand for each forest type under this management type could be sampled, and thus replication of combinations of management types × forest types was not used. The size of the stands varied from 0.4 to 4.8 ha (Table 1).
| Table 1. Size of the studied forest stands by type and management. | |||
| Forest type | Size of the forest stands, ha | ||
| NAT | SALNAT | SALPLA | |
| Cladinoso-callunosa | 0.4 | 2.1 | 1.6 |
| Vacciniosa | 3.1 | 4.8 | 1.3 |
| Myrtilloso-sphagnosa | 1.5 | 2.0 | 1.5 |
| Sphagnosa | 2.0 | 1.0 | 0.8 |
| NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. | |||
Sampling of plots was conducted in summer in 2017 and 2018. At 10 locations along a transect traversing each stand, at 20–35 m regular intervals (depending on stand size) four sample plots were set up in azimuth directions; accordingly, in each forest type 40 sample plots were assessed. In total 480 sample plots (1 × 1 m) were established. In each sample plot cover (%) of all herb layer vascular plants (including tree seedlings with height <0.5 m), bryophyte and lichen species, as well as total herb and moss-lichen cover, was estimated. In addition, in a 25 m2 circular sample plot (surrounding the vegetation sampling plots), tree height and diameter at 1.3 m height (DBH) were determined for all regenerated and survived trees with height >0.5 m; diameter was given a zero value for trees with height <1.3 m. In data analysis, plots in the dry types that had been placed in small wet depressions with mire species were discarded (8 plots).
In each sample plot centre, tree stand crown openness was determined from hemispherical images obtained using a digital camera with a wide angle, (fish-eye) photo lens and tripod, which was located 1.3 m above the ground, and the WinSCANOPY program (WinSCANOPY 2005) was used for image analysis.
2.3 Data analysis
Species richness between management types was compared with one-way ANOVA analysis for each forest type. The effect of canopy openness and survived and regenerated tree variables (diameter, height, and calculated basal area) on plant community composition constrained by management type was assessed with permutational multivariate analysis of variance (adonis2) (Oksanen et al. 2019) using R 3.6.1 (R Core Team 2019) by paired analysis of management types. Detrended correspondence analysis (DCA) using PC-Ord 7.07 was used to visualize the main gradients in plant community composition that were linked with canopy openness and regenerating tree variables in permutational multivariate analysis of variance. Species found in only one plot were removed from these analyses.
To determine species that showed affinity to a particular management type, indicator species analysis (Dufrêne and Legendre 1997) was conducted for each forest type. Calculated indicator values (IVs), which are based on both frequency of occurrence and abundance (cover), were assessed for statistical significance using Monte Carlo randomization with 9999 permutations. Each forest type was analysed separately, and therefore, permutation was restricted to occur between three management types. The indicator status of species from each forest and management type was assessed by the statistical significance (p > 0.05) and using an arbitrarily defined cut off observed IV > 30 to omit weak values. An IV value of 100 indicates fidelity to one group. Lower values show that the species occurred more than in one group.
3 Results
3.1 Effect of management type
Permutational multivariate analysis of variance showed significant effect of management, canopy openness, and regenerated saplings (>0.5 m height) and survived tree variables on species composition (Table 2 and 3). In the Cladinoso-callunosa forest type, canopy openness was not related to differences in community composition constrained by management type (Table 3), but the dimensions of survived and regenerating Scots pine (Pinus sylvestris L.) showed significant relationships with vegetation (Table 2). Pine regeneration was greater in the SALNAT management type and survived pine in the NAT type. In the other forest types, canopy openness and tree variables were significantly associated with vegetation composition constrained by management type. In the Vacciniosa forest type, openness was significantly higher in the NAT site and lower in the SALNAT and SALPLA sites. Pine had greater height and DBH in the salvage-logged sites, while density was higher in the NAT management type. Silver birch (Betula pendula Roth) regenerated best in the NAT and SALNAT management types (Table 2). In the Myrtilloso-sphagnosa forest type, vegetation composition showed significant relationship with birch, Norway spruce (Picea abies (L.) H. Karst.) and black alder (Alnus glutinosa (L.) Gaertn.) variables, but not with pine. Canopy openness was higher in the SALNAT management type in the Myrtilloso-sphagnosa forest type (Table 3), and birch, spruce and alder variables were higher in the NAT management type (Table 2). In the Sphagnosa forest type, vegetation composition was significantly associated with birch sapling and survived pine variables, which were higher in the SALNAT management type for birch and in the NAT management type for survived pine (Table 2).
| Table 2. Effect of regenerating saplings (>0.5 m height) on vegetation composition constrained by management type in permutational multivariate analysis of variance. Mean height, DBH (diameter at breast height), basal area, and density of saplings are given. The significant effects are shown in bold. The letters designate significant differences according to the pairwise Tukey’s post-hoc test. The tests were conducted separately for each of the four forest types. View in new window/tab. |
| Table 3. Measured mean canopy openness and its effect on species composition constrained by management type as determined by paired permutation multivariate analysis of variance. Significant effects are shown in bold and letters indicate significant differences according to the pairwise Tukey’s post-hoc test. The tests were conducted separately for each of the four forest types. | |||||||||
| Mean canopy openness | NAT vs SALNAT | SALNAT vs SALPLA | SALPLA vs NAT | ||||||
| NAT | SALNAT | SALPLA | p-value | F-value | p-value | F-value | p-value | F-value | |
| Cladinoso-callunosa | 45.3 | 20.3 | 47.9 | 0.8 | 0.5 | 0.02 | 2.3 | 0.07 | 1.7 |
| Vacciniosa | 32.7A | 25.2B | 24.8C | <0.001 | 4.2 | 0.008 | 2.9 | <0.001 | 4.2 |
| Myrtilloso-sphagnosa | 24.7B | 53.6A | 23.9C | 0.002 | 3.8 | <0.001 | 6.3 | 0.04 | 1.9 |
| Sphagnosa | 61.7A | 27.4B | 47.9B | 0.04 | 2.1 | <0.001 | 5.8 | 0.2 | 1.2 |
| NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. | |||||||||
3.2 DCA analysis
DCA ordination showed good separation of clusters for the NAT, SALNAT and SANPLA management types, but with overlap (Figs. 2–5). The DCA ordination of species showed significant correlation of the species gradients with canopy openness and sampling trees for all forest types. For the Vacciniosa, Myrtilloso-sphagnosa and Sphagnosa forest types the correlation of openness and sapling trees with species composition was related to separation of clusters of the management types along DCA axis 1. However, this was not the case for the Cladinoso-callunosa type, where openness was correlated to DCA axis 2, which showed variability in species composition within and not between (Table 3) the management types.
In the Cladinoso-callunosa type, mean diameter and height of pine saplings (>0.5m) was associated with DCA axis 1 in the direction of the two types of salvage logging clusters of plots. Species on the left side of the ordination included the common feathermoss species like Pleurozium schreberi and Hylocomium splendens. The cover of pine seedlings (<0.5 m height) and Vaccinium species was higher on the opposite side of the ordination (Fig. 2). The openness gradient along DCA axis 2, which was associated with Cladonia species (Fig. 2), was in the opposite direction of the vector for number of birch samplings.
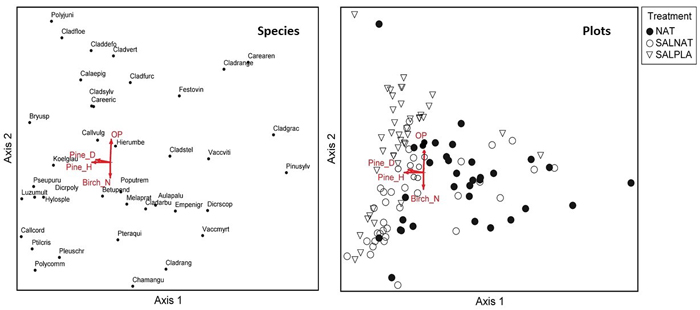
Fig. 2. Species and sample plot ordination (Detrended correspondence analysis) for Cladinoso callunosa stands. Vectors show correlations between the vegetation gradients and canopy openness (OP), mean diameter at breast height of regenerating Scots pine ( Pine_D ), mean height of regenerating Scots pine (Pine_H) and density of regenerating Silver birch (Birch_N). NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. Species abbreviations are shown in full in Supplementary file S1. View larger in new window/tab.
In the Vacciniosa forest type, openness and the NAT management type on the right side of the DCA ordination were associated with species like Vaccinium myrtillus and pine seedlings. DCA axis 2 was suggestive of a soil fertility gradient for the salvage-logged plots, with herb species like Trientalis europaea and the shrub species Sorbus aucuparia on the top and lichen species on the bottom of the ordination (Fig. 3). These plots occurred near the edges of dune slacks where black alder also occurred.
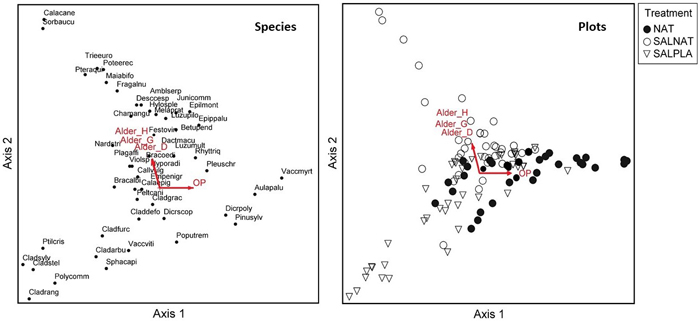
Fig. 3. Species and sample plot ordination (Detrended correspondence analysis) for Vacciniosa stands. Vectors show correlations between the vegetation gradients and canopy openness (OP), mean diameter at breast height of regenerating black alder (Alder_D), mean height of regenerating black alder (Alder_H) and basal area of regenerating black alder (Alder_G). NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. Species abbreviations are shown in full in Suppl. file S1. View larger in new window/tab.
In the ordination of the wet Myrtilloso-sphagnosa type, where openness was associated with the SALNAT management type (Fig. 4), species characteristic of the openness end of the gradient were Sphagnum girgensohnii and Calluna vulgaris, along with number of pine and spruce saplings (Fig. 4). However, pine size (diameter, basal area and height) were associated with the opposite side of the gradient in the direction of the SALPLA plot cluster, with herb and shrub species typical of rich wet forest types, like Lysimachia vulgaris, Galium rivale, Frangula alnus and Rubus saxatilis.
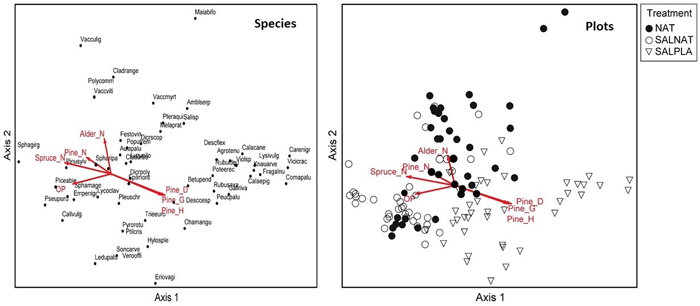
Fig. 4. Species and sample plot ordination (Detrended correspondence analysis) for Myrtilloso-sphagnosa stands. Vectors show significant correlations of vegetation gradients and canopy openness (OP), mean diameter at breast height of regenerating Scots pine (Pine_D), mean height of regenerating Scots pine (Pine_H), basal area of regenerating Scots pine (Pine_G), density of regenerating Norway spruce (Spruce_N), Scots pine (Pine_N) and black alder (Alder_N). NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. Species abbreviations are shown in full in Suppl. file S1. View larger in new window/tab.
For the Sphagnosa type, number, height, diameter and basal area of both birch and pine were correlated with DCA axis 1 in the direction of the SALNAT cluster of plots (Fig. 5). Typical poor mire species like Oxycoccus palustris and Sphagnum species occurred on the right side of the ordination and typical boreal forest low shrub (V. myrtillus and C. vulgaris) and moss species (Dicranum polysetum and Aulacomnium palustre) on the left side, which was associated with greater canopy openness (Fig. 5).
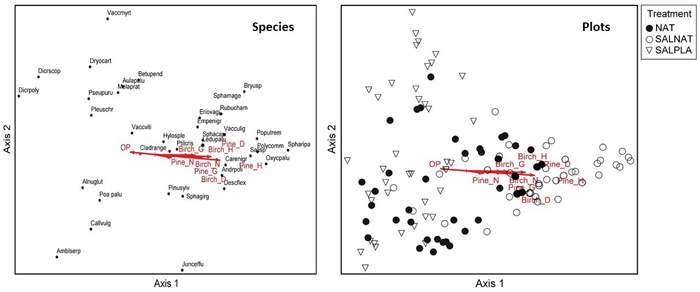
Fig. 5. Species and sample plot ordination (Detrended correspondence analysis) for Sphagnosa stands. Vectors show significant correlations of vegetation gradients and canopy openness (OP), basal area of regenerating Silver birch (Birch_G) and Scots pine (Pine_G), mean height of regenerating Silver birch (Birch_H) and Scots pine (Pine_H), mean diameter at breast height of regenerating Scots pine (Pine_D) and Silver birch (Birch_D), and density of regenerating Scots pine (Pine_N) and Silver birch (Birch_N). NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting Species abbreviations are shown in full in Suppl. file S1. View larger in new window/tab.
3.3 Indicator species analysis
Indicator species analysis did not identify specific species for a particular management type across all forest types. In the poor Cladinoso-callunosa, dry forest type, Cladonia species, Vaccinium vitis-idaea and pine seedlings were indicators of the NAT management type and other Cladonia species and Polytrichum juniperinum were indicators of the SALPLA management type. Pleurozium schreberi was an indicator for the SALNAT management type. In the Vacciniosa forest type, the NAT management type was associated with Vaccinium myrtillus, while the species Hylocomium splendens, Calluna vulgaris were indicators of the SALNAT management type and Polytrichum commune of the SALPLA management type (Table 4). In the wet Myrtilloso-sphagnosa forest type, species characteristic of the SALNAT management type were Sphagnum girgensohnii and Calluna vulgaris (Table 4), while herb and shrub species typical of rich wet types, like Lysimachia vulgaris, Galium rivale, Frangula alnus and Rubus saxatilis, along with birch seedling were indicator species for the SALPLA management type. Indicator species for the NAT management type in the wet Myrtilloso-sphagnosa forest type were Vaccinium vitis-idaea and Polytrichum commune. In the Sphagnosa forest type, Sphagnum girgensohnii was the only indicator species of the NAT management type. Typical poor mire species like Oxycoccus palustris and Ledum palustre were indicators of the SALNAT management type. Birch seedlings were common in the SALPLA management type along with typical boreal forest low shrub (V. myrtillus and C. vulgaris) and moss species (Dicranum polysetum and Aulacomnium palustre) in the Sphagnosa forest type (Table 4).
| Table 4. Indicator species of management by forest types. P-values and indicator values (IV) are given. The analysis was conducted separately for each of the four forest types. | |||||||||
| NAT | SALNAT | SALPLA | |||||||
| Species | p-value | IV | Species | p-value | IV | Species | p-value | IV | |
| Cladinoso-callunosa | Dicranum scoparium Hedw. | <0.001 | 81.3 | Melampyrum pratense L. | <0.001 | 85.0 | Cladonia deformis (L.) Hoffm. | 0.002 | 77.0 |
| Cladonia rangiferina (L.) Weber ex F.H. Wigg. | <0.001 | 51.0 | Pleurozium schreberi (Willd. ex Brid) Mitt. | 0.007 | 37.7 | Polytrichum juniperinum Hedw. | 0.002 | 38.8 | |
| Cladonia arbuscula (Wallr.) Flot. | <0.001 | 38.1 | Calluna vulgaris (L.) Hull | 0.044 | 38.5 | ||||
| Pine seedlings Pinus sylvestris L. | <0.001 | 45.0 | Cladonia furcata (Huds.) Schrad., 1794 | <0.001 | 67.9 | ||||
| Vaccinium vitis-idaea L. | 0.001 | 45.2 | |||||||
| Vacciniosa | Vaccinium myrtillus L. | <0.001 | 54.8 | Calluna vulgaris (L.) Hull | 0.01 | 41.9 | Polytrichum commune Hedw. | <0.01 | 59.6 |
| Aulacomnium palustre (Hedw.) Schwägr. | <0.001 | 33.3 | Festuca ovina L. | <0.001 | 55.6 | ||||
| Luzula pilosa (L). Willd. | <0.001 | 30.8 | |||||||
| Melampyrum pratense L. | <0.001 | 57.7 | |||||||
| Pteridium aquilinum (L.) Kuhn | <0.001 | 39.0 | |||||||
| Hylocomium splendens (Hedw.) Schimp. | <0.001 | 55.0 | |||||||
| Myrtilloso-sphagnosa | Vaccinium vitis-idaea L. | 0.002 | 46.2 | Calluna vulgaris (L.) Hull | <0.001 | 59.2 | Birch seedlings Betula pendula Roth | <0.001 | 74.6 |
| Polytrichum commune Hedw. | <0.001 | 53.2 | Sphagnum girgensohnii Russow | <0.001 | 82.5 | Eriophorum vaginatum L. | <0.001 | 39.8 | |
| Sphagnosa | Sphagnum girgensohnii Russow | 0.005 | 39.5 | Oxycoccus palustris Pers. | <0.001 | 50.2 | Birch seedlings Betula pendula Roth | <0.001 | 59.2 |
| Salix spp. | <0.001 | 42.5 | Calluna vulgaris (L.) Hull | 0.006 | 38.6 | ||||
| Polytrichum commune Hedw. | <0.001 | 88.6 | Vaccinium myrtillus L. | <0.001 | 48.2 | ||||
| Sphagnum riparium Ångstr. | <0.001 | 30.0 | Dicranum polysetum Sw. | <0.001 | 64.5 | ||||
| Ledum palustris L. | 0.023 | 34.2 | Aulacomnium palustre (Hedw.) Schwägr. | <0.001 | 54.4 | ||||
| NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. | |||||||||
3.4 Species richness
Number of species (species richness) did not significantly differ in Cladinoso-callunosa and Sphagnosa forest types between management types. Species richness was significantly higher in the Vacciniosa forest type under SALNAT management, and in the Myrtilloso-sphagnosa type under SALPLA management (Table 5).
| Table 5. Species richness according to forest type and management. Significant effects are shown in bold (according to ANOVA). The different letters indicate significant differences between management types according to the pairwise Tukey’s post-hoc test. The analysis was conducted separately for each of the four forest types. | |||
| Number of species (with mosses) | |||
| NAT | SALNAT | SALPLA | |
| Cladinoso-callunosa | 19 | 26 | 22 |
| Vacciniosa | 16A | 40B | 26A |
| Myrtilloso-sphagnosa | 29A | 23A | 49B |
| Sphagnosa | 22 | 31 | 22 |
| NAT – natural regeneration, SALNAT – salvage logged and left for natural regeneration, SALPLA – salvage logging with planting. | |||
4 Discussion
Varying effect on ground vegetation of canopy openness and management types across forest types was found. This implies that effects of salvage logging are dependent on forest types. Indicator species analysis, which was used to show affinity of species to a particular management type, did not identify any species that were not typical species of the respective forest types. Therefore, salvage logging did not appear to have created novel successional paths of plant community development with invasive or ruderal species, nor with species not typical of the respective forest types. Post-fire management can have different effect in different forest types, which may be explained by its impact on soil, post-disturbance legacies, like survived and dead trees, and regenerated trees, which in turn may have influenced understory vegetation composition (Nilsson and Wardle 2005; Jõgiste et al. 2017).
Not only understory vegetation, but also the tree layers of stands, can be destroyed by intense fire, while surviving trees affect further regeneration (Adámek et al. 2016). In our case the basal area of survived trees was very low (<10 m2 ha–1) and no survived trees were observed in the SALPLA management type. However, the basal area of survived pines was significantly associated with vegetation composition in Vacciniosa and Sphagnosa forest types, which might be explained by increased shading around the solitary and clumps of large trees. Larger mean tree height in the salvage-logged stands in the dry forest types, indicates more rapid pine regeneration, which can be explained by ground disturbance (Brūmelis et al. 2005), and of course planting of pine in the SALPLA management type. However, cover of pine seedlings was higher in the NAT management type in the poorest forest type, which can be explained by slower tree growth and low cover of feathermoss species allowing microsites for establishment. Birch, spruce and black alder regeneration was improved in the NAT and SALNAT stands in the wet rich Myrtilloso-sphagnosa forest type, and birch in the wet poor Sphagnosa forest type.
Canopy openness reflects the canopy cover due to surviving and regenerated trees that colonized the stands already at the first years after the disturbance (Donis 1995). The lack of the significant differences in species richness in the Cladinoso-callunosa forest type among management types might be explained by lack of significant differences in canopy openness or another factor like soil degradation after fire (Dale et al. 2001). The changing light conditions during the succession may lead to more diverse understory vegetation in the burned area due to an increase of shade-tolerant tree species (Hart and Chen 2008). Higher species richness in the SALNAT site in the Vacciniosa forest type and in the SALPLA site for the Myrtilloso-sphagnosa forest type suggest a positive effect of salvage logging for species richness. In the Vacciniosa forest type, species richness was lowest in the NAT management type where the canopy was more open and highest in the SALNAT management type where the canopy was more closed. Black alder in the SALNAT site in the dry Vacciniosa forest type occurred in some plots close to edges of dune slacks, which was typical for the dune complex. The significant association of this trees species with vegetation can also be explained by this edge effect.
Indicator species analysis showed that Calluna vulgaris was more common in the SALPLA and SALNAT management sites in all forest types. This suggests that salvage logging caused greater soil disturbance by removal of the upper litter layer with competitive species, creating suitable conditions for this fire-adapted species (Nilsson et al. 1996; Reich et al. 2001; Parro et al. 2009). In typical Cladinoso-callunosa forest type five years after fire there is a transition from Calluna vulgaris cover to Vaccinium vitis-idaea and Vaccinium myrtillus cover (Nilsson et al. 1996; Norberg et al. 2001; Nilsson and Wardle 2005), which were more common in the SALPLA management, shown by indicator species analysis. Salvage logging after fire creates more suitable habitat for ericaceous dwarf shrubs, but pine forest stand natural regeneration can be hindered for up to 50 years because of great cover of Calluna vulgaris (Vacchiano et al. 2013). Pine seedlings were associated with NAT management type for both dry stands, possibly because the Calluna vulgaris stage promoted by salvage logging was avoided.
The Myrtilloso-sphagnosa stands were severely damage during the fire, as canopy trees were lacking in all management types. In wet forest types light condition can significantly change microclimate, causing stands to become drier, which adversely affects regeneration (Lindenmayer et al. 2009). In both wet types, the colonization of birch, spruce, alder and pine in the understory of the salvage logged and planted management type can be explained by surface disturbance creating establishment niches and more suitable (richer or moister) forest types. In the Myrtilloso-sphagnosa type, the ordinations suggested that greater growth of regenerating pine was associated with the SALPLA management, but in NAT and SALNAT with number of regenerating alder, pine and spruce. The better natural regeneration of trees (number) in these the NAT and SALNAT can be explained by suitable microsites and sunlit conditions created by fire and salvage logging. In wet stands, trees can establish on dense Sphagnum hummocks in well-lit conditions (Brūmelis et al. 2009). Planting of pine in the SALPLA management likely increased shading, as suggested by decreased openness, which created unsuitable conditions for further establishment, but led to faster canopy development. In the Sphagnosa type, the ordination showed that SALNAT plots had greater number and size of regenerated pine and birch, indicating a positive effect of salvage logging. However, there was no evidence that planting in the SALPLA management was associated with more rapid development of a closed canopy.
The naturally regenerating NAT stands contained fallen deadwood, which likely affected regeneration. If stands after fire have a high amount of fallen deadwood with rapid decomposition, a large amount of nutrients can be released improving stand regeneration, but slowly decomposing deadwood might hinder understory vegetation establishment (Parro et al. 2015). In naturally regenerating (NAT) stands, this might partly explain the more open canopy in the Vacciniosa and Sphagnosa types, the generally low species richness on all forest types, and lower number of indicator species (only one indicator species) in the Sphagnosa forest type, and patches of Cladonia species in the Cladinoso-callunosa type. Also, as the study was carried out 25 years after the initial disturbance, we lack information on the effect of initial conditions on vegetation composition and tree regeneration, and can report only the present situation. The differences between management types may have been more pronounced initially, and the vegetation over time has become more similar to typical vegetation of the forest types across the management types.
5 Conclusion
In this study, we found that post-fire management, canopy openness and regenerating trees had significant effect on species composition and richness in four different forest types. In all studied forest types, although vegetation composition significantly differed among management types, all species were typical of the habitats with no ruderal or invasive species. Our hypothesis of this study was rejected, as in the dry and wet forest stands, the combined effect of fire and management was associated with higher species richness and more rapid development of tree saplings. Thus, considering ground vegetation cover and tree regeneration, salvage logging was shown to generally facilitate stand recovery after disturbance, but the effect differed between forest types. However, this study included only ground vegetation, hence, we did not consider the species associated with coarse woody debris that was retained without salvage logging.
Acknowledgments
The study was funded by the joint stock company “Latvijas valsts meži” project “Impact of forest management on forest and related ecosystem services”.
References
Adámek M., Hadincová V., Wild J. (2016). Long-term effect of wildfires on temperate Pinus sylvestris forests: Vegetation dynamics and ecosystem resilience. Forest Ecology and Management 380: 285–295. https://doi.org/10.1016/j.foreco.2016.08.051.
Bonan G.B., Shugart H.H. (1989). Environmental factors and ecological processes in boreal forests. Annual Review of Ecology, Evolution, and Systematics 20: 1–28. https://doi.org/10.1146/annurev.es.20.110189.000245.
Brūmelis G., Elferts D., Liepiņa L., Luce I., Tabors G., Tjarve D. (2005). Age and spatial structure of natural Pinus sylvestris stands in Latvia. Scandinavian Journal of Forest Research 20(6): 471–480. https://doi.org/10.1080/02827580500339526.
Brūmelis G., Strazds M., Eglava Z. (2009). Stand structure and spatial pattern of regeneration of Pinus sylvestris in a natural treed mire in Latvia. Silva Fennica 43(5): 76–781. https://doi.org/10.14214/sf.172.
Buhk C., Meyn A., Jentsch A. (2007). The challenge of plant regeneration after fire in the Mediterranean Basin: scientific gaps in ourknowledge on plant strategies and evolution of traits. Plant Ecology 192: 1–19. https://doi.org/10.1007/s11258-006-9224-2.
Dale V.H., Joyce L.A., McNulty S., Neilson R.P., Ayres M.P., Flannigan M.D., Hanson P.J., Irland L.C., Lugo A.E., Peterson C.J., Simberloff D., Swanson F.J., Stock B.J., Wotton M. (2001). Climate change and forest disturbances: climate change can affect forests by altering the frequency, intensity, duration, and timing of fire, drought, introduced species, insect and pathogen outbreaks, hurricanes, windstorms, ice storms, or landslides. BioScience 51(9): 723–734. https://doi.org/10.1641/0006-3568(2001)051[0723:CCAFD]2.0.CO;2.
Donis J. (1995). Forest regeneration following 1992 fire in Slītere National Park. Mežzinātne: Meža nozares augstākās izglītības 75. Gadu jubilejai veltītās zinātniski praktiskās konferences materiāli. LLU, Jelgava. p 80–88. [In Latvian].
Donis J., Kitenberga M., Snepsts G., Matisons R., Zarins J., Jansons A. (2017). The forest fire regime in Latvia during 1922–2014. Silva Fennica 51(5) article 7746. https://doi.org/10.14214/sf.7746.
Drobyshev I., Granström A., Linderholm H.W., Hellberg E., Bergeron Y., Niklasson M. (2014). Multi‐century reconstruction of fire activity in Northern European boreal forest suggests differences in regional fire regimes and their sensitivity to climate. Journal of Ecology 102(3): 738–748. https://doi.org/10.1111/1365-2745.12235.
Dufrêne M., Legendre P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67(3): 345–366. https://doi.org/10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2.
Gromtsev A. (2002). Natural disturbance dynamics in the boreal forests of European Russia: a review. Silva Fennica 36(1): 41–55. https://doi.org/10.14214/sf.549.
Hart S.A., Chen H.Y.H. (2008). Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forests. Ecological Monographs 78(1): 123–140. https://doi.org/10.1890/06-2140.1.
Jõgiste K., Korjus H., Stanturf J.A., Frelich L., Baders E., Donis J., Jansons A., Kangur A., Koster K., Laarmann D., Maaten T., Marozas V., Metslaid M., Nigul K., Polyachenko O., Randveer T., Vodde F. (2017). Hemiboreal forest: natural disturbances and the importance of ecosystem legacies to management. Ecosphere 8(2) article e01706. https://doi.org/10.1002/ecs2.1706.
Kitenberga M., Drobyshev I., Elferts D., Matisons M., Adamovics A., Katrevics J., Niklasson M., Jansons A. (2019). A mixture of human and climatic effects shapes the 250-year long fire history of a semi-natural pine dominated landscape of Northern Latvia. Forest Ecology and Management 441: 192–201. https://doi.org/10.1016/j.foreco.2019.03.020.
Kurulok S.E., Macdonald E.S. (2007). Impacts on postfire salvage logging on understory plant communities of the boreal mixedwood forests 2 and 34 years after disturbance. Canadian Journal of Forest Research 37(12): 2637–2651. https://doi.org/10.1139/X07-107.
Kuuluvainen T. (1994). Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: a review. Annales Zoologici Fennici 31: 35–51.
Lindenmayer D.B., Noss R.F. (2006). Salvage logging, ecosystem processes, and biodiversity conservation. Conservation Biology 20(4): 949–958. https://doi.org/10.1111/j.1523-1739.2006.00497.x.
Lindenmayer D.B., Ought K. (2006). Salvage logging in the montane ash eucalypt forests of the Central Highlands of Victoria and its potential impacts on biodiversity. Conservation Biology 20(4): 1005–1015. https://doi.org/10.1111/j.1523-1739.2006.00501.x.
Lindenmayer D.B., Hunter M.L., Burton P.J., Gibbons P. (2009). Effects of logging on fire regimes in moist forests. Conservation Lettters 2(6): 271–277. https://doi.org/10.1111/j.1755-263X.2009.00080.x.
Nilsson M.-C., Wardle D.A. (2005). Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment 3(8): 127–138. https://doi.org/10.1890/1540-9295(2005)003[0421:UVAAFE]2.0.CO;2.
Nilsson M.-C., Steijlen I., Zackrisson O. (1996). Time-restricted seed regeneration of Scots pine in sites dominated by feather moss after clear-cutting. Canadian Journal of Forest Research 26(6): 945–953. https://doi.org/10.1139/x26-104.
Nilsson S.G. (1997). Forests in the temperate–boreal transition: natural and man-made features. Ecological Bulletins 46: 61–71. https://doi.org/10.1007/978-1-4615-3524-9_10.
Norberg G., Dolling A., Jäderlund A., Nilsson M.C., Zackrisson O. (2001). Control of heather (Calluna vulgaris (L.) Hull) by steam treatment: effects on establishment and early growth of Scots pine. New Forests 21: 187–198. https://doi.org/10.1023/a:1011874222978.
Parro K., Köster K., Jõgiste K., Vodde F. (2009). Vegetation dynamics in a fire damaged forest areas: the response of major ground vegetation species. Baltic Forestry 15: 206–215.
Parro K., Metslaid M., Renel G., Sims A., Stanturt J.A., Jõgiste K., Köster K. (2015). Impact of post fire management on forest regeneration in a managed hemiboreal forest, Estonia. Canadian Journal of Forest Research 45(9): 1192–1197. https://doi.org/10.1139/cjfr-2014-0514.
Parviainen J. (1996). The impact of fire on Finnish forests in the past and today. Silva Fennica 30(2–3): 353–359. https://doi.org/10.14214/sf.a9246.
Peterson D.W., Dodson E.K. (2016). Post-fire logging produces minimal persistent impacts on understory vegetation in northeastern Oregon, USA. Forest Ecology and Management 370: 56–64. https://doi.org/10.1016/j.foreco.2016.04.004.
Reich P.B., Bakken P., Carlson D., Frelich L.E., Friedman S.K., Grigal D.F. (2001). Influence of logging, fire, and forest types on biodiversity and productivity in southern boreal forests. Ecology 82(10): 2731–2748. https://doi.org/10.1890/0012-9658(2001)082[2731:IOLFAF]2.0.CO;2.
Rowe J.S. (1983). Concepts of fire effects on plant individual and species. In: Wein R.W., MacLean D.A. (eds.). The role of fire in northern circumpolar ecosystems. John Wiley & Sons Ltd. p 135–154.
Rowe J.S., Scotter G. (1973). Fire in the boreal forest. Quaternary Research 3(3): 444–464. https://doi.org/10.1016/0033-5894(73)90008-2.
Oksanen J., Blanchet G.F., Friendly M., Kindt R., Legendre P., McGlinn D., Minchin P.R., O’Hara R.B., Simpson G.L., Solymos P., Stevens H.H.M., Szoecs E., Wagner H. (2019). Community ecology package. https://github.com/vegandevs/vegan.
Thorn S., Bässler C., Svoboda M., Müller J. (2016). Effects of natural disturbances and salvage logging on biodiversity – lessons from the Bohemian Forest. Ecology and Management 388: 113–119. https://doi.org/10.1016/j.foreco.2016.06.006.
Trabaud L. (1987). Fire and survival traits of plants. In: Trabaud L. (ed). The role of fire in ecological systems. SPB Publishing, The Hague. p. 65–89.
Vacchiano G., Lonati M., Berretti R., Motta R. (2013). Drivers of Pinus sylvestris L. regeneration following small, high-severity fire in a dry, inner-alpine valley. Plant Biosystems 149(2): 354–363. https://doi.org/10.1080/11263504.2013.819821.
Wallenius T.H., Kuuluvainen T., Vanha-Majamaa I. (2004). Fire history in relation to site type and vegetation in Vienansalo wilderness in eastern Fennoscandia, Russia. Canadian Journal of Forest Research 34(7): 1400–1409. https://doi.org/10.1139/x04-023.
Wallenius T. (2011). Major decline in fires in coniferous forests – reconstructing the phenomenon and seeking for the cause. Silva Fennica 45(1): 139–155. https://doi.org/10.14214/sf.36.
WinSCANOPY (2005). WinSCANOPY 2005 for hemispherical image analysis. Regent Instruments Inc., Canada.
Total of 37 references.
