Noé Dumas,
Mathieu Dassot,
Jonathan Pitaud,
Jérôme Piat,
Lucie Arnaudet,
Claudine Richter,
Catherine Collet 
Four-year-performance of oak and pine seedlings following mechanical site preparation with lightweight excavators
Dumas N., Dassot M., Pitaud J., Piat J., Arnaudet L., Richter C., Collet C. (2021). Four-year-performance of oak and pine seedlings following mechanical site preparation with lightweight excavators. Silva Fennica vol. 55 no. 2 article id 10409. https://doi.org/10.14214/sf.10409
Highlights
- Mechanical site preparation (MSP) with lightweight excavators controls highly competitive plant species (Molinia caerulea, Pteridium aquilinum) much more efficiently than MSP with conventional methods
- This MSP approach improves four-year survival and growth of pine seedlings, but it is less evident for oak seedlings.
Abstract
Mechanical site preparation methods that used tools mounted on lightweight excavators and that provided localised intensive preparation were tested in eight experimental sites across France where the vegetation was dominated either by Molinia caerulea (L.) Moench or Pteridium aquilinum (L.) Kuhn. Two lightweight tools (Deep Scarifier: DS; Deep Scarifier followed by Multifunction Subsoiler: DS+MS) were tested in pine (Pinus sylvestris L., Pinus nigra var. corsicana (Loudon) Hyl. or Pinus pinaster Aiton) and oak (Quercus petraea (Matt.) Liebl. or Quercus robur L.) plantations. Regional methods commonly used locally (herbicide, disk harrow, mouldboard plow) and experimental methods (repeated herbicide application; untreated control) were used as references in the experiments. Neighbouring vegetation cover, seedling survival, height and basal diameter were assessed over three to five years after plantation. For pines growing in M. caerulea, seedling diameter after four years was 37% and 98% greater in DS and DS+MS, respectively, than in the untreated control. For pines growing in P. aquilinum, it was 62% and 107% greater in the same treatments. For oak, diameter was only 4% and 15% greater in M. caerulea, and 13% and 25% greater in P. aquilinum, in the same treatments. For pines, the survival rate after four years was 26% and 32% higher in M. caerulea and 64% and 70% higher in P. aquilinum, in the same treatments. For oak, it was 3% and 29% higher in M. caerulea and 37% and 31% higher in P. aquilinum. Herbicide, when applied for three or four years after planting, provided the best growth performances for pines growing in M. caerulea and P. aquilinum and for oaks growing in P. aquilinum. For these species and site combinations, DS+MS and DS treatments reduced the neighbouring vegetation cover for one to four years following site preparation.
Keywords
vegetation control;
reforestation;
seedling growth;
seedling survival
- Dumas, Université de Lorraine, AgroParisTech, INRAE, UMR Silva, 54000 Nancy, France E-mail noe.dumas@inrae.fr
- Dassot, EcoSustain, Environmental Engineering Office, Research and Development, 31, rue de Volmerange, 57330 Kanfen, France; Institut National de l’Information Géographique et Forestière, 1 rue des Blanches Terres, 54250 Champigneulles, France E-mail mathieu.dassot@ign.fr
- Pitaud, Office National des Forêts, Département Recherche Développement et Innovation, route d’Amance, 54280 Champenoux, France E-mail jonathan.pitaud@onf.fr
- Piat, Office National des Forêts, Département Recherche Développement et Innovation, 3 rue du petit château, 60200 Compiègne, France E-mail jerome.piat@onf.fr
- Arnaudet, Office National des Forêts, Département Recherche Développement et Innovation, 100 boulevard de la Salle, 45760 Boigny-sur-Bionne, France E-mail lucie.arnaudet@onf.fr
- Richter, Office National des Forêts, Département Recherche Développement et Innovation, Boulevard de Constance, 77300 Fontainebleau, France E-mail claudine.richter@onf.fr
-
Collet,
Université de Lorraine, AgroParisTech, INRAE, UMR Silva, 54000 Nancy, France
 https://orcid.org/0000-0003-0861-7796
E-mail
catherine.collet@inrae.fr
https://orcid.org/0000-0003-0861-7796
E-mail
catherine.collet@inrae.fr
Received 26 June 2020 Accepted 19 April 2021 Published 29 April 2021
Views 101281
Available at https://doi.org/10.14214/sf.10409 | Download PDF

Supplementary Files
1 Introduction
Rapid establishment, high seedling survival rate and fast early growth are the basis of successful forest plantation. In most plantation sites, the development of young trees may be seriously impaired by problems with the physical, chemical or biological characteristics of soils or by competition from neighbouring vegetation (Wagner et al. 2006; du Toit et al. 2010). Mechanical site preparation (MSP) is widely used to alleviate such constraints and create suitable microsites where the seedlings can establish and grow and, ultimately, enhance plantation success (Burton et al. 2000). In some countries, the use of herbicides is now strictly controlled (Ammer et al. 2011; Thiffault and Roy 2011) and MSP is receiving increasing attention since it represents an alternative to chemicals to control the development of competing vegetation.
Managing the vegetation and the soil by MSP has proven to be effective if an appropriate method is used (von der Gönna 1992; Löf et al. 2012). In the first place, MSP should aim at controlling the vegetation and reducing its negative effect until the young trees reach a height where they have outcompeted the vegetation. Depending on their morphological and biological features (root density, root depth, plant cover, plant height, seed bank, sprouting ability, clonal propagation, growth rate), plants respond differently to the disturbance caused by MSP and, in order to be effective, the choice of the MSP method must take the features of the neighbouring vegetation into account (Balandier et al. 2006).
Similarly, multiple soil constraints commonly require different approaches and fine adjustments to site preparation treatments. Soil water content is a first major constraint. Poorly drained soils strongly restrict tree establishment and growth by decreasing the amount of oxygen available for roots (Drew and Lynch 1980). Trees planted on such soils usually benefit from raised planting positions that elevate the trees above the water table level and may be combined with the formation of ditches that drain excessive water out of the stand (Hallsby and Örlander 2004). In contrast, on dry sites, it is recommended to plant trees in trenches where snow and run-off may accumulate to give the young trees better access to soil water. An additional soil constraint frequently found in young plantations is soil compaction (Cambi et al. 2017). Many forest soils have compacted horizons that may be of natural origin or caused by vehicle traffic (Smith and Johnston 2001). These hardened horizons form a barrier to tree root growth by restricting tap root depth and diminishing fine root density within the volume of soil explored by the root systems (Ampoorter et al. 2011). Surface or deep tillage that fractures and decompacts soil horizons and reduces soil bulk density has been successfully applied in such compacted soils to increase rooting volume and improve root growth and activity (Lacey et al. 2001).
There is a wide range of soil and vegetation constraints that impact seedling establishment. Soil and vegetation characteristics are not independent and these constraints often occur in specific combinations. MSP methods are designed to deal with certain sets of constraints and generally will not be efficient to alleviate another type of constraint (Nilsson et al. 2010; Cole et al. 2018). In some situations commonly found in western and central European forests at this time, there is no known effective mechanical treatment to mitigate existing soil and vegetation constraints (McCarthy et al. 2011), e.g., sites with deep-rooted competing plant species such as Pteridium aquilinum (L.) Kuhn, usually found on dry sites, or Molinia caerulea (L.) Moench, often found on highly compacted clayey and loamy soils with high water tables. Both species rapidly develop after clear-cutting and are strong resource competitors to the detriment of young seedlings (Gaudio et al. 2011; Vernay et al. 2016; Fernandez et al. 2020). The site conditions where P. aquilinum and M. caerulea prevail cover several thousand hectares in France. These challenging site conditions and desires for non-chemical site preparation approaches have created strong demand for MSP tools that are designed to alleviate these specific constraints and eventually enhance seedling establishment.
Intensive soil preparation is required to provide effective control of deep-rooted neighbouring plant species, to decompact the soil to a depth sufficient for the seedlings to establish, or to create pits and mounds to elevate seedlings above the water table in sites with strong waterlogging. However, MSP methods that induce strong soil disturbances may not be acceptable at a time when there is a demand for low impact practices that maintain long-term soil fertility (Duncker et al. 2012). Consequently, there is a clear need to develop MSP methods that alleviate strong vegetation and soil constraints while controlling the overall soil disturbance induced.
New MSP tools have been designed to control deep-rooted competing plant species and to reduce soil compaction or soil waterlogging, with limited impacts on the soil. The tools are mounted on lightweight excavators that have a high manoeuvrability and allow localised spot or line preparation. The tools perform locally intensive site preparation that leads to strong soil disturbances around the seedlings (Quibel 2020; Collet et al. 2020) and that leaves large unprepared zones between the planting spots, whereas the intensity of soil disturbance evaluated at stand scale is low (Quibel 2020). In addition, the low weight of the machines leads to reduced deep soil compaction under the wheel tracks (Collet et al. 2020). These tools seem to provide an interesting compromise between the intensity of soil disturbance required to reduce the constraints to seedling establishment and the overall soil disturbance estimated at stand scale, but their actual effects on seedling performance have not yet been evaluated.
The objective of this study is to evaluate the effects of such tools on seedling survival and growth, and to test whether the tools actually enhance seedling performance. We established an experimental network with eight sites dominated either by either P. aquilinum or M. caerulea, with different soil constraints, across different regions in France. We used two tree species at each site that have contrasting early growth dynamics: different pine species (Pinus sylvestris L., Pinus nigra var. corsicana (Loudon) Hyl. or Pinus pinaster Aiton) that establish rapidly and show strong early growth, and different oak species (Quercus petraea (Matt.) Liebl. or Quercus robur L.) that establish and grow more slowly than pines. We expected that (hypothesis H1) the new lightweight tools provided more efficient vegetation control around the seedlings than conventional methods involving disk harrowing, mouldboard plowing or herbicide application, and (hypothesis H2) seedlings growing in plots prepared with lightweight MSP tools would show greater survival, height and diameter growth than seedlings growing in unprepared plots or in plots prepared with conventional methods. We also expected that (hypothesis H3) seedling responses to MSP would be more pronounced for pine than for oak seedlings.
2 Material and methods
2.1 Study sites and experimental design
Experimental sites (Table 1) were established in one of two types of soil conditions: sites on highly compacted, waterlogged, sandy-silt acidic soils with a vegetation dominated by M. caerulea, and sites on dry sandy acidic soils with a vegetation dominated by P. aquilinum. A total of eight sites were established, which were distributed among four regions. For all sites, the altitude was lower than 300 m and the slope was less than 10%. Annual average temperature ranged between 10.4 and 13.5 °C, and annual precipitation between 730 and 900 mm. Climate was oceanic with continental influence in Haguenau, and oceanic in all other sites. Previous stand type varied among sites (broadleaved, conifers, mixed) as well as the stand removal method used (progressive regeneration cut, clear-cutting, windstorm). In each site, all remaining overstory was removed and all remaining slash was crushed one year before the experiment started.
| Table 1. Characteristics of the study sites: forest name, annual mean temperature (Tmean), annual precipitation, geographic location, site name, soil and humus type, dominant vegetation species, previous stand type and previous stand removal information. Climatic data are public data from Météo France. | |||||||||
| Forest | Tmean (°C) | Precipitation (mm) | Site | Location | Soil type | Humus type | Vegetation | Previous stand type | Stand removal type and date |
| Bord-Louviers | 11 | 730 | BL | 49°18´N, 01°09´E | Sandy-loamy | Dysmoder | Pteridium aquilinum | 120-year-old Scots pine | Windstorm 1999 |
| Haguenau | 10.4 | 758 | H1 | 48°50´N, 07°55´E | Sandy | Mor | Pteridium aquilinum | >100-year-old mixed Scots pine, beech and pedunculate oak | Windstorm 1999 |
| H2 | 48°49´N, 07°55´E | Sandy-loamy | Hydromor | Molinia caerulea | >50-year-old mixed Scots pine and oak | Windstorm 1999 | |||
| H3 | 48°49´N, 07°53´E | Sandy-loamy | Hydromor | Molinia caerulea | 25 to 30-year-old Scots pine | Windstorm 1999 | |||
| Villecartier | 11 | 850 | VI | 48°28´N, 01°33´W | Loamy | Mull | Pteridium aquilinum | >140-year-old beech | Progressive regeneration cut 2010 |
| Rennes | 12 | 860 | RE | 48°13´N, 01°32´W | Loamy | Hydromor | Molinia caerulea | 180-year-old oak | Progressive regeneration cut 2010 |
| Escource | 13.5 | 900 | ES | 44°12´N, 01°05´W | Sandy | Mor | Pteridium aquilinum | 50-year-old maritime pine | Clear cut 2008 |
| Solférino | 13.5 | 900 | SO | 44°08´N, 00°53´W | Sandy | Hydromor | Molinia caerulea | 50-year-old maritime pine | Clear cut 2008 |
Two tools mounted on lightweight excavators (2.8 tons) were used in our study (Supplementary file S1):
- The deep scarifier (Scarificateur Réversible®, Kirpy, Layrac, France) consists of three 40-cm-long curved shanks on which three 60-cm-long tines and two 20-cm-long intercalary curved shanks are fixed. This tool can fracture the soil structure down to 40 cm at the most, and extracts plant root systems. It is mainly used to control competing plant species with dense and deep root systems, such as P. aquilinum. The tool extracts the vegetation (roots and shoots), shakes it thoroughly to remove soil parts stuck to the roots and lays it aside in small piles. In M. caerulea sites, the dense root system of the grass prevents the efficient separation of the soil from the roots, leading to the creation of a slight ground depression.
- The multifunction subsoiler (Sous-soleur Multifonction®, Kirpy) is made up of two parts: a rake on the top of the tool to clear away debris and remove the aboveground part of the neighbouring vegetation, and a curved tine with two lateral wings and a pointed tip to provide deep soil fracture and to form a mound. It is mainly used in compacted and poorly drained soils. It tills the soil down to 50 cm at the most and may be used to form a 20-cm-high mound where the seedling may be planted.
Five site preparation methods were compared in each site:
- DS, deep scarifier. 1 to 1.5-m-wide lines spanning the planting rows were prepared at the end of summer. The depth of soil preparation ranged from 20 to 30 cm, depending on the site.
- DS+MS, deep scarifier followed by multifunction subsoiler. Following the DS, the MS was used on the same lines to till the soil and create a mound. MS preparation was performed immediately following DS preparation. Depending on the site, the depth of soil cultivation ranged from 35 to 45 cm, and the height of the mound from 10 to 20 cm.
- HE, repeated herbicide application. Herbicides were applied each year, for a period of 1 to 4 years after planting, which varied among sites. See Table 2 for details. Herbicides were applied at the beginning of summer, and the first application took place during the summer before planting.
- RR, regional reference site preparation method. A conventional method locally used by forest managers in similar ecological conditions (herbicide, disc harrow or mouldboard plow) was used and differed among regions. See Table 2 for details.
- C, untreated control: no site preparation.
| Table 2. Description of the experimental sites: planted tree species, planting date, mechanical site preparation (MSP) treatment date, experimental design and characteristics of HE and RR treatments. Treatment names: DS+MS, deep scarifier followed by multifunction subsoiler; HE, herbicide; RR, regional reference. View in new window/tab. |
In each site, cultivation profiles, a method to analyse the impacts of cultivation practices on soil structure (Collet et al. 2020), were performed in treatment DS, DS+MF and C. An average profile for treatments DS and DS+MF is shown in Suppl. file S1.
In each site, an oak species and a pine species were compared. These oak and pine species were chosen for their contrasted life traits: pines generally show higher survival rates and more rapid growth trajectories after planting than oaks (Boutte 2015, 2020). Two or three-year-old bare root (1+1 or 1+2) oak and one-year-old containerised pine seedlings were planted during the winter season. All pine seedlings had been chemically treated against Hylobius abietis (Linnaeus, 1758) in the nursery with an insecticide (Imidacloprid). Sites were fenced in order to suppress damage by ungulates and lagomorphs.
In each site, the experimental design contained two or three blocks. Within each block, the five treatments were randomly assigned to a plot. After site preparation, each plot was divided into two equally-sized split plots, and each split plot was planted either with oak or with pine. Planted buffer zones of at least 2 m in width were established around each plot, and measurements were carried out only in the central zone of the plots. The distance between planting rows varied between 2 and 6 m, depending on site and treatment (treatment DS+MF required larger spacing between planting rows), and the distance between trees along the row was 1 m at all sites. Accordingly, split plot size varied between 1190 and 1960 m2, and tree density in split plots varied between 2200 and 3300 trees per ha. The measurement zone of each split plot included 68 to 96 trees, either pine or oak.
High mortality rates occurred during the first year in some sites and for some species (see Results). At these sites, the split plots were either fully replanted the next year or were abandoned. At the three Haguenau sites (H1, H2, H3), oak showed a high mortality rate due to strong winter frost. At H1, oak was replanted in all treatments. At H2 and H3, the losses were high only in the DS+MS treatment. At these two sites, oaks in the DS+MS treatment were replanted and an additional HE split plot was added to each block and planted the same year to take potential year effects due to differences in climate or in planting stock into account. Therefore, all oaks at H1, as well as the oaks in the DS+MS and the HE treatments at H2 and H3, were one year younger than the other seedlings at the site. These split plots were considered as a separate experimental site in all of the statistical analyses because they were planted a different year and used a different seedling stock. For all Haguenau sites (H1, H2, H3), the pines were unaffected by the winter frost event. In Escource (ES) and Solférino (SO), both oak and pine showed high mortality due to winter frost and summer drought, leading to the abandonment of the two sites. At all of the other sites (BL, RE, VI), seedling mortality was lower and the sites were not replanted.
As explained above, the same experimental design was used for all of the experimental sites. However, pronounced differences among sites in total area, local reference site preparation treatments, plantation species, and adverse climatic conditions resulted in treatment plots that differed in (1) the way the MSP method was applied in each treatment (maximum depth reached by the tool, width of the prepared area, height of the mound); (2) the planted species and stock type; (3) the planting density; (4) the planting year; and (5) the time between site preparation and planting. Any variation in these factors was assumed to have been captured within the experimental site factor.
2.2 Measurements and data processing
Each year during winter, basal diameter and total height were measured on all seedlings in the measurement zones (68 to 96 seedlings per species, across 10 to 15 plots, at each site). The status (alive or dead) and, for dead seedlings, the most probable cause of death, were recorded.
Eight 1-m2 observation subplots were established in the measurement zone of each plot for vegetation measurements. Subplots were centred on the planting rows and located between two trees. One subplot was established on each planting row, and its position along the row was randomly chosen. Each year in September, plant species were determined and the cover (the ratio of the surface area occupied by the vertical projection of the plants on the ground, over the ground surface area described, multiplied by 100) was visually estimated for each species. Only individuals rooted inside the subplot were taken into account.
The number of species per subplot varied between 0and 11 (median value: 3.0). In total, 114 species were recorded in the study. Plant species were pooled into groups in order to avoid analysing many species with a low presence in each subplot. Seven groups were defined, adapted from Balandier (2006): (1) P. aquilinum; (2) M. caerulea; (3) other graminoids (sedges and rushes, grasses other than M. caerulea); (4) Rubus fruticosus L.; (5) Calluna vulgaris (L.) Hull and Erica cinerea L.; (6) woody plants other than Ericaceae; and (7) other species. For each record, the cover of each of the seven groups (cover_record) was computed as the sum of the cover of all individual species included in the group. Since plants within a group belonged to the same plant layer, it was possible to combine them by summing their individual cover values (Chytry et al. 2005; Fischer 2015). However, nine values (out of a total of 6517 values) were slightly higher than 100% and were equalled to 100%.
Total plant cover was estimated using the cover_record values for the seven groups. Combining plants belonging to different layers is not straightforward (Chytry et al. 2005), and Fischer (2015) proposed an index to combine the values of multiple layers into a unique value that takes the overlap between the layers into account. For each subplot and each measurement date, total plant cover was computed as:
![]()
where i, p, and y indicated plant group, subplot, and year, respectively. By construction, plant_cover ranged between 0and 100%.
In order to estimate the contribution of each group to plant_cover, the method developed by Fischer (2015) was used to compute group values that would add up to plant_cover. First, for each group, plant cover was averaged per site, treatment and year (cover_average) as the mean of eight cover_record values corresponding to the eight subplots. For each site, treatment and year combination, the seven groups were then ranked in descending order of cover_average values. Second, a cover index (cover_index) for each group in each subplot and each measurement date was then computed recursively, starting with the group with the highest cover_average value, as follows:
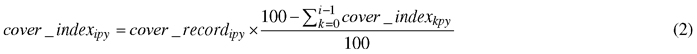
where i (from 1 to 7) indicated the ith plant group, sorted by descending order of cover_average within each site, treatment and year combination, p; and y indicated subplot and year, respectively, and where cover_index0py was set to 0.
2.3 Statistical analyses
The effects of site preparation on seedling survival one and four years after plantation were analysed separately using beta regression. Because of high short-distance spatial autocorrelation in early seedling mortality, survival was analysed using survival rates at split plot scale. Because of the low number of split plot replicates per experimental site, species were pooled into two types (pines and oaks), and experimental site was not introduced in the models. The following models were used:
![]()
where logit(Ssvt) was the logit transformation of the mean survival rate [0–1] after one year or after four years of seedlings in split plots corresponding to species s (pine or oak) in vegetation type v (M. caerulea or P. aquilinum) in treatment t; µ was the overall mean, and α, β and γ were parameters to be estimated. The second-order interaction among the three factors was not significant and was removed, first-order interactions were tested and only significant interactions were kept in the final models. Likelihood ratio tests on nested models were used to estimate factor effects. To test whether differences among treatments were significant, post-hoc pairwise comparisons were performed using treatment estimated marginal means.
To analyse the effects of site preparation on seedling growth, analyses of variance for seedling diameter and seedling height at the end of the experiment (i.e., four years after planting) were performed. Factors thought to affect the responses were incorporated into the initial models: treatment, main vegetation type and tree species as fixed factors, experimental site and plot as random factors. First-order interactions among factors were introduced into the initial models. Experimental site x treatment and species x treatment interactions were highly significant for both height and diameter, and separate models were then fitted for each species and each experimental site. Since separate models were fitted for each experimental site, main vegetation type (Molina or P. aquilinum) was removed. Plot and its interaction with treatment were introduced as random effects, but the interaction was not significant and was removed from the models. Since heteroscedastic standardised residuals were observed, a variance-structure term that permitted different levels of variance among treatments was added. This option significantly improved the models. The resulting models, established for each experimental site and each species present in the experimental site, were as follows:
![]()
where yitp was the seedling height or basal diameter after 4 years of seedling i in treatment t and plot p; µ was the overall mean; α and β were parameters to be estimated; and ε corresponded to the residuals, normally distributed within each treatment, such that ![]() Plot was a random effect nested within treatment and was assumed to be normally distributed. The models finally retained for the analyses did not compare the experimental sites or the species. To test whether the differences among treatments were significant within each experimental site and each species, post-hoc Tukey-adjusted pairwise comparisons were performed.
Plot was a random effect nested within treatment and was assumed to be normally distributed. The models finally retained for the analyses did not compare the experimental sites or the species. To test whether the differences among treatments were significant within each experimental site and each species, post-hoc Tukey-adjusted pairwise comparisons were performed.
The effects of site preparation on total plant cover were examined for each site and each year separately, using beta regression. To test whether the differences among treatments were significant within each experimental site and each year, post-hoc pairwise comparisons were performed.
All analyses were carried out in R 3.5.2 (R Core Team 2018) and models were fitted and tested with the betareg (Cribari-Neto and Zeileis 2010), emmeans (Length 2020) and nlme (Pinheiro et al. 2019) packages. In all analyses, the accepted level of significance was α = 0.05.
3 Results
3.1 Seedling survival
Treatment effects on seedling survival after one growing season varied considerably with site, year and tree species, and no clear trend appeared. Regression models that pooled the three pine species into a pine group and pooled the two oak species into an oak group (Eq. 3) indicated no significant effects of tree species (p = 0.68) and treatment (p = 0.87). The final model including only the main vegetation type was significant (p < 0.001) but weak (pseudo-R2 = 0.17). In some experimental sites and some years (Table 3), survival rate was high for all treatments (pine: sites BL, H1, H2 and RE; oak: sites BL and RE) while in other sites, seedlings died during the first winter in all treatments (pine: site SO). In some experimental sites and years, differences among treatments appeared but were not uniform, and the outcome differed from one site to the other. In site VI, pine seedlings in the control performed the worst, with a survival rate of 64% compared to more than 85% in all other treatments. On the contrary, in the 2011 oak plantation of site H3, DS+MS showed the lowest survival rate (25%) compared to survival rates of between 80% and 91% in other treatments. Oak and pine seedlings planted the same year in the same site may show similar (site BL) or different (site SO) treatment responses. A species in a given site may show different responses in two successive years (oak in site H1 planted in 2011 and 2012). Some differences observed among treatments could be ascribed to specific events that occurred in the experimental sites during the first year: winter frost (site SO in 2011, sites H1, H2 and H3 in 2011), summer drought (site ES), and root damage by cockchafers (Melolontha spp.) (sites H1, H2 and H3 in 2011).
| Table 3. Survival after one growing season of pine and oak seedling planted in eight experimental sites in Northern France, in different site preparation treatments (HE, herbicide; RR, regional reference; DS, deep scarifier; DS+MS, deep scarifier followed by multifunction subsoiler; C, untreated control). | ||||||||
| Site | Tree species | Vegetation | Plantation year | Survival rate (%) | ||||
| HE | RR | DS | DS+MS | C | ||||
| BL | Pinus nigra | Pteridium aquilinum | 2010 | 95 | 97 | 97 | 100 | 93 |
| H1 | Pinus sylvestris | Pteridium aquilinum | 2011 | 93 | 99 | 95 | 97 | 98 |
| VI | Pinus sylvestris | Pteridium aquilinum | 2011 | 93 | 87 | 98 | 94 | 64 |
| ES | Pinus pinaster | Pteridium aquilinum | 2011 | 50 | 61 | 62 | 62 | 44 |
| H2 | Pinus sylvestris | Molinia caerulea | 2011 | 99 | 99 | 100 | 100 | 95 |
| H3 | Pinus sylvestris | Molinia caerulea | 2011 | 99 | 97 | 100 | 100 | 79 |
| RE | Pinus sylvestris | Molinia caerulea | 2011 | 96 | 95 | 93 | 97 | 93 |
| SO | Pinus pinaster | Molinia caerulea | 2011 | 0 | 0 | 0 | 0 | 0 |
| BL | Quercus petraea | Pteridium aquilinum | 2010 | 97 | 98 | 94 | 96 | 93 |
| H1 | Quercus petraea | Pteridium aquilinum | 2011 | 69 | 68 | 24 | 2 | 43 |
| H1 | Quercus petraea | Pteridium aquilinum | 2012 | 89 | 86 | 89 | 85 | 77 |
| VI | Quercus petraea | Pteridium aquilinum | 2011 | 86 | 91 | 89 | 84 | 79 |
| ES | Quercus robur | Pteridium aquilinum | 2011 | 8 | 28 | 29 | 35 | 5 |
| H2 | Quercus petraea | Molinia caerulea | 2011 | 97 | 94 | 97 | 48 | 92 |
| H2 | Quercus petraea | Molinia caerulea | 2012 | 81 | - | - | 85 | - |
| H3 | Quercus petraea | Molinia caerulea | 2011 | 87 | 80 | 85 | 25 | 91 |
| H3 | Quercus petraea | Molinia caerulea | 2012 | 75 | - | - | 92 | - |
| RE | Quercus petraea | Molinia caerulea | 2011 | 98 | 97 | 96 | 98 | 98 |
| SO | Quercus robur | Molinia caerulea | 2011 | 58 | 70 | 88 | 82 | 67 |
In contrast, treatment effects on seedling survival after four growing seasons showed stronger trends. Regression (Eq. 3) indicated significant effects of tree species type (pines vs. oak) (p < 0.001), main vegetation (p = 0.005), and treatment (p < 0.001), with an interaction between species and vegetation (p < 0.001). The final model that included these effects showed a pseudo-R2 = 0.33. Marginal means were 63% (a), 76% (b), 77% (b), 83% (bc) and 88% (c) for treatments C, HE, RR, DS and DS+MS, respectively (means followed by the same letter did not significantly differ).
Treatment effects on pine seedling survival after four growing seasons showed similar trends for all experimental sites (Table 4). In all sites, seedlings in the control performed worse, with a survival ranging from 40% to 79% (median: 60.5%). In other treatments, survival ranged from 63% to 97% (median: 90.5%). The highest survival rates were observed for MSP treatments, in general, DS+MS (ranging from 90% to 99%, median: 96%).
| Table 4. Survival after four growing seasons of pine and oak seedling planted in eight experimental sites in Northern France, in different site preparation treatments (HE2, HE3 and HE4 herbicide applied during 2, 3 and 4 growing seasons, respectively; RR, regional reference; DS, deep scarifier; DS+MS, deep scarifier followed by multifunction subsoiler; HE, herbicide; C, untreated control). | ||||||||||
| Site | Tree species | Vegetation | Plantation year | Survival rate (%) | ||||||
| HE2 | HE3 | HE4 | RR | DS | DS+MS | C | ||||
| BL | Pinus nigra | Pteridium aquilinum | 2010 | - | 87 | - | 90 | 97 | 99 | 65 |
| H1 | Pinus sylvestris | Pteridium aquilinum | 2011 | - | - | 76 | 67 | 68 | 91 | 51 |
| VI | Pinus sylvestris | Pteridium aquilinum | 2011 | 63 | - | - | 73 | 91 | 90 | 40 |
| H2 | Pinus sylvestris | Molinia caerulea | 2011 | - | - | 90 | 97 | 97 | 97 | 84 |
| H3 | Pinus sylvestris | Molinia caerulea | 2011 | - | - | 93 | 89 | 91 | 97 | 56 |
| RE | Pinus sylvestris | Molinia caerulea | 2011 | 92 | - | - | 89 | 89 | 96 | 79 |
| BL | Quercus petraea | Pteridium aquilinum | 2010 | - | 90 | - | 97 | 90 | 90 | 82 |
| H1 | Quercus petraea | Pteridium aquilinum | 2012 | - | - | 84 | 57 | 79 | 78 | 47 |
| VI | Quercus petraea | Pteridium aquilinum | 2011 | 55 | - | - | 84 | 85 | 75 | 56 |
| H2 | Quercus petraea | Molinia caerulea | 2011 | - | - | 79 | 78 | 89 | - | 84 |
| H2 | Quercus petraea | Molinia caerulea | 2012 | - | - | 70 | - | - | 77 | - |
| H3 | Quercus petraea | Molinia caerulea | 2011 | - | - | 72 | 66 | 78 | - | 66 |
| H3 | Quercus petraea | Molinia caerulea | 2012 | - | - | 63 | - | - | 87 | - |
| RE | Quercus petraea | Molinia caerulea | 2011 | 54 | - | - | 50 | 46 | 89 | 57 |
Oak seedlings in P. aquilinum sites performed worse in the control and short-term herbicide treatments (two years), with a survival ranging between 47 and 82% in these treatments (Table 4). In other treatments, survival ranged between 57 and 97%. In M. caerulea sites, results were very heterogeneous and oak survival responded very differently to treatment depending on the experimental site.
3.2 Pine growth
After four growing seasons, pine height was comprised between 73.5 and 205.0 cm, and basal diameter between 13.5 and 57.2 mm. Models for pines (Eq. 4) led to statistically significant effects of treatments on pine seedling basal diameter in all sites (Fig. 1). In height models, treatment effect was statistically significant in sites BL, H1and H2, but not in sites VI, H3 and RE (Fig. 2).
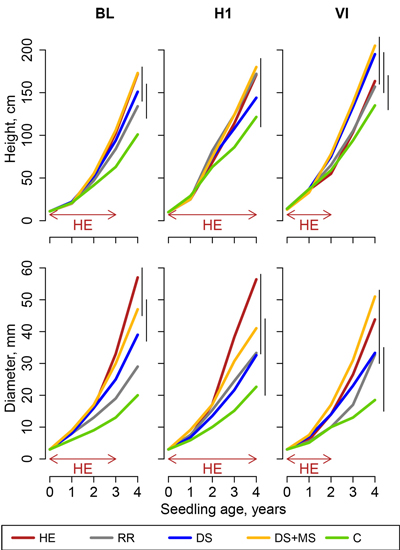
Fig. 1. Height and basal diameter of pine seedlings in Pteridium aquilinum sites (sites BL, H1 and VI, Northern France), following site preparation (HE: herbicide application; RR: regional reference; DS: deep scarifier; DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). The horizontal arrow at the bottom of the graph indicates the length of time of herbicide application in treatment HE in each experimental site (from 2 to 4 years). The probability associated with the treatment effect is indicated in a linear model performed on height at the age of 4. Vertical segments on the right-hand side of the plot indicate the results of t-tests used for post-hoc mean comparisons (treatment means joined by a segment do not significantly differ).
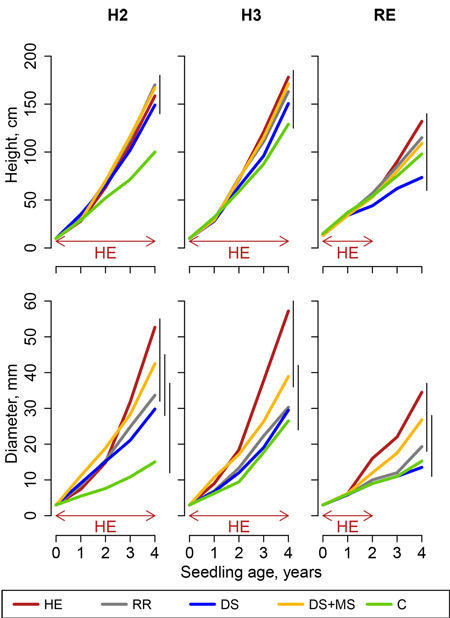
Fig. 2. Height and basal diameter of pine seedlings in Molinia caerulea sites (sites H2, H3 and RE, Northern France), following site preparation (HE: herbicide application; RR: regional reference; DS: deep scarifier; DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). The horizontal arrow at the bottom of the graph indicates the length of time of herbicide application in treatment HE in each experimental site (from 2 to 4 years). The probability associated with the treatment effect is indicated in a linear model performed on height at the age of 4. Vertical segments on the right-hand side of the plot indicate the results of t-tests used for post-hoc mean comparisons (treatment means joined by a segment do not significantly differ).
In P. aquilinum sites, pine basal diameter after four growing seasons was highest for HE and DS+MS treatments. Differences with C were statistically significant in the three experimental sites (Fig. 1). Differences between HE and DS+MS did not statistically differ in any experimental site, but the HE treatment tended to perform better in sites BL and H1 where the herbicide was applied during three or four growing seasons, contrary to site VI where the herbicide was only applied during two growing seasons. Seedlings in the control performed the worst in all experimental sites, even though differences with treatments DS and RR were not always significant. Response differences among other treatments were more difficult to interpret, even though DS treatment seemed to perform better than C but worse than DS+MS in most experimental sites.
Seedling height tended to respond similarly to diameter (higher seedlings in HE and DS+MS than in C), but statistically significant differences among treatments within each experimental site occurred less frequently.
Pine diameter responded quite similarly in M. caerulea sites (Fig. 2). HE was significantly more effective than C in the three experimental sites. It was more effective than other treatments in all experimental sites, although the differences were not always statistically significant. DS+MS was the most efficient treatment after HE, and seemed better than C and other MSP treatments. In site RE, where the waterlogging constraint was highest, seedling height and diameter were the lowest for the DS treatment.
Differences in height were much smaller, and heterogeneity among experimental sites was greater for height than for diameter. In experimental sites H2 and H3, treatment C performed worse than other treatments, and differences were significant only for site H2. In site RE, seedlings were highest in treatment HE and lowest in DS.
3.3 Oak growth
Overall, oak appeared to grow slower than pine, with a period of one, two, three or four years after planting, a period during which height increments remained below or close to zero. After four growing seasons, oak height was comprised between 34.5 and 179 cm and basal diameter between 7.9 and 25.3 mm. In most experimental sites, treatment effects were less distinct in oak than in pines. Models for oak (Eq. 4) led to statistically significant effects of treatments on seedling basal diameter in all sites except site VI and H2 (Fig. 3). In height models, treatment effect was statistically significant only in sites BL, H1 (2nd plantation) and H2 (2nd plantation), but not in other sites (Fig. 4).
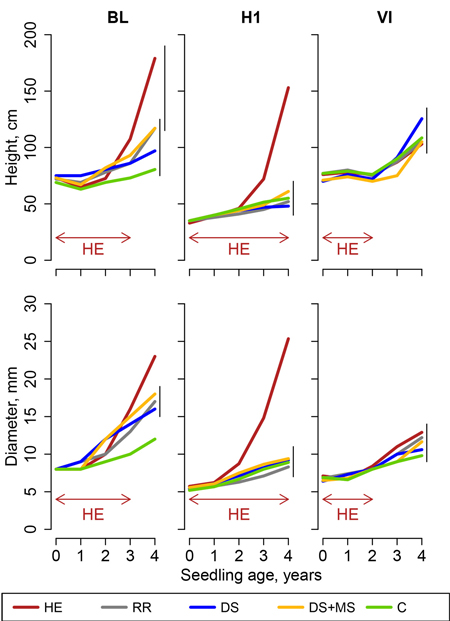
Fig. 3. Height and basal diameter of sessile oak seedlings in Pteridium aquilinum sites (sites BL, H1 and VI, Northern France), following site preparation (HE: herbicide application; RR: regional reference; DS: deep scarifier; DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). The horizontal arrow at the bottom of the graph indicates the length of time of herbicide application in treatment HE in each experimental site (from 2 to 4 years). The probability associated with the treatment effect is indicated in a linear model performed on height at the age of 4. Vertical segments on the right-hand side of the plot indicate the results of t-tests used for post-hoc mean comparisons (treatment means joined by a segment do not significantly differ).
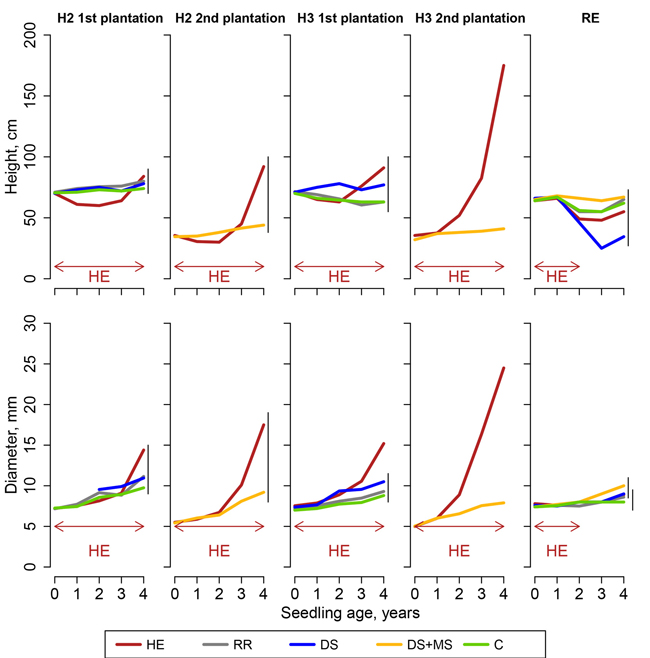
Fig. 4. Height and basal diameter of sessile oak seedlings in Molinia caerulea sites (sites H2, H3 and RE; H2 and H3 were planted twice, Northern France), following site preparation (HE: herbicide application; RR: regional reference; DS: deep scarifier; DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). The horizontal arrow at the bottom of the graph indicates the length of time of herbicide application in treatment HE in each experimental site (from 2 to 4 years). The probability associated with the treatment effect is indicated in a linear model performed on height at the age of 4. Vertical segments on the right-hand side of the plot indicate the results of t-tests used for post-hoc mean comparisons (treatment means joined by a segment do not significantly differ).
In P. aquilinum sites, oak diameter and height were greater in HE than in other treatments, except for height in site VI where herbicide was only applied during two growing seasons (Fig. 3). In sites H1 and VI, MSP treatments did not perform significantly better than the control. In site BL, diameter in treatments DS, DS+MS and RR was significantly greater than in treatment C but not significantly different from each other.
In M. caerulea sites, oak seedlings only grew very slowly in C and in all MSP treatments (Fig. 4) in all experimental sites. With the exception of site RE, where herbicide was only applied during two growing seasons, HE was the only treatment where oak reached a diameter of above 11 mm after four years. In site RE, where waterlogging was the greatest, average height slightly decreased for all treatments. In site RE, treatment DS even gave seemingly worse results than the control.
3.4 Plant cover
In all years, mean plant cover (plant_cover) in treatment C ranged from 67% to 100% (Fig. 5 and 6), with the exception of site BL during the first year where plant_cover was low (44.2%). The low value may have been caused by a floristic inventory made soon after a late frost that damaged P. aquilinum. In all control plots, the vegetation was strongly dominated either by P. aquilinum or M. caerulea.
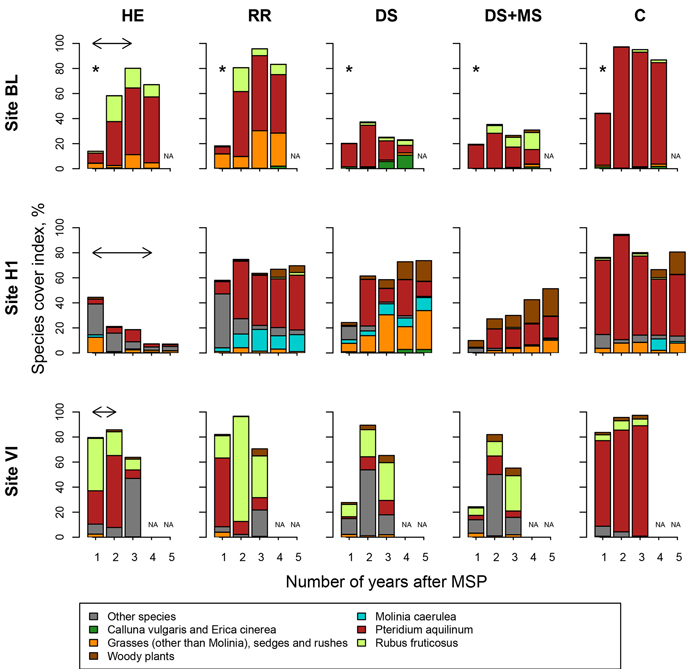
Fig. 5. Plant cover in Pteridium aquilinum sites (sites BL, H1 and VI, Northern France) following site preparation (HE: herbicide application; RR: regional reference site preparation method; DS: deep scarifier; DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). For each species, plant cover was estimated using Eq. 2. In treatment HE, the horizontal arrow at the top of the graph indicates the length of time of herbicide application in each experimental site (from 2 to 4 years). Plant species were pooled into seven groups. The asterisks above the first year in site BL correspond to a floristic inventory carried out soon after late frosts in May that reduced P. aquilinum cover. NA indicates missing values for the corresponding year.
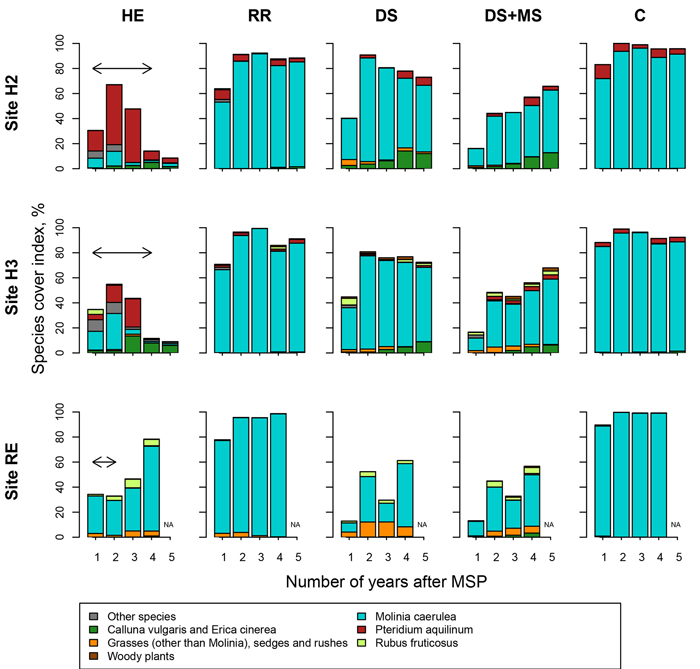
Fig. 6. Plant cover in Molinia caerulea sites (sites H2, H3, RE, Northern France) following site preparation (HE: herbicide application; RR: regional reference site preparation method; DS: deep scarifier, DS+MS: deep scarifier followed by multifunction subsoiler; C: untreated control). For each species, plant cover was estimated using Eq. 2. In treatment HE, the horizontal arrow at the top of the graph indicates the length of time of herbicide application in each experimental site (from 2 to 4 years). Plant species were pooled into seven groups. NA indicates missing values for the corresponding year.
In all sites, herbicide treatments significantly reduced plant_cover values, and the duration of the effect was related to the duration of the herbicide application: two years in VI and RE, three years in BL, five years in H1, H2 and H3 (Suppl. file S2). Herbicide strongly increased plant diversity in sites H2 and H3, whereas the vegetation remained dominated by M. caerulea in site RE.
Regional reference treatments that involved MSP (treatment RR in sites H1, H2, H3, RE and VI) only slightly decreased plant_cover values compared to C (Suppl. file S2), and always for a short length of time (no more than one year in most sites). In M. caerulea sites (H2, H3, RE), vegetation composition was not affected, whereas in P. aquilinum sites (BL, H1, VI) plant diversity seemed greater than in the control.
Lightweight MSP had a strong effect on plant_cover values: it ranged between 13% and 45% and between 10% and 24% in the first year in treatments DS and DS+MS, respectively, and the effect lasted between one and four years, depending on the site (Suppl. file S2). In site BL where the effect was the most pronounced, plant_cover values after four years were 23% and 31% in treatments DS and DS+MS, respectively. Both treatments changed species composition, especially in P. aquilinum sites. In most sites, the effects on plant_cover values and on species composition were stronger in DS+MS than in DS.
MSP treatments (DS, DS+MS, RR, in sites H1, H2, H3, RE and VI) decreased plant_cover values compared to C for time periods comprised between one and five years depending on site. Impact on vegetation composition was also highly variable: in site H1, P. aquilinum cover was much smaller in DS and DS+MS treatments than in C after five growing seasons, whereas in site H2, M. caerulea was still the dominant species after the same length of time.
4 Discussion
4.1 Impacts of MSP on competing plant species and soils
Some plant species such as P. aquilinum and M. caerulea, but also other grass species (e.g., Deschampsia cespitosa (L.) P. Beauv.) or ericaceous species, which are very common in central and western Europe, may seriously impair seedling establishment (McCarthy et al. 2011; Gaudio et al. 2011). Because of their deep and dense root systems, these species are very difficult to control using traditional MSP methods. The two tools, DS and MS, were specifically designed to reduce plant species that are difficult to control mechanically. In our study, they proved to be very efficient at reducing the cover of P. aquilinum and M. caerulea and provided a two- to four-year time window for the seedlings to grow with reduced competition. These time windows may be considered as sufficient to allow rapid seedling establishment (Wagner and Robinson 2006). They were much longer than those obtained in the Regional Reference treatment, which represented conventional MSP tools (one year in all experimental sites). As expected, these methods were insufficient to allow successful seedling establishment in these site conditions.
After MSP, P. aquilinum may recolonize the prepared area mainly by rapidly resprouting from living roots remaining in the deep soil horizons, or by slowly growing from adjacent unprepared areas with undisturbed plants (Dolling 1999). DS and MS tools were very efficient because they were able to almost completely remove P. aquilinum root systems in the prepared area, preventing any resprouting. M. caerulea mainly recolonizes the prepared area by resprouting from remaining roots, by growing from dormant seeds stored in the upper soil layers or by growing from new seeds dispersed from adjacent unprepared areas (Taylor et al. 2001; Brys et al. 2005). Contrary to traditional tools, the DS and MS tools completely removed the grass root system and the dormant seeds and set them 1 m to the side. After a preparation using the DS or MS tools, new M. caerulea plants mostly originated from dispersed seeds, strongly curtailing its recolonization.
In our study, MSP treatments improved seedling growth and survival in most situations. However, when applied over a sufficient duration (at least three to four years), herbicide treatments outclassed most MSP treatments. Although MSP methods differ greatly from one study to another, the literature globally tends to show that MSP and herbicides both improve seedling growth and survival. Some studies suggest that the best results are obtained by a combination between mechanical and herbicide treatments (Löf et al. 2012) while others demonstrate that herbicide alone performs better than MSP (e.g., Sutherland and Foreman 2000). Our own study tends to show the latter, especially if the herbicide is applied during at least three growing seasons. Since herbicide treatments only reduce vegetation constraints but do not modify soil constraints (as opposed to MSP treatments), our results confirm the major role of competing vegetation on seedling growth and survival even in sites with high soil constraints (drought, waterlogging, and/or compaction).
In addition to removing neighbouring vegetation, MSP also modifies the soil structure (Bock and Van Rees 2002). It may improve soil structure, mainly by loosening and aerating the soil, which is expected to enhance seedling root development (Neaves et al. 2017), but it may also induce soil compaction due to machine traffic (Aust et al. 2004), soil smearing caused by the tool (Kees 2008) or lateral export of soil and organic matter, which all may have detrimental effects on the growth and functioning of seedling roots (Cambi et al. 2017). These changes in the soil structure may, in turn, induce a series of positive and negative side effects: reduction of organic matter (Yildiz et al. 2009), nutrient leaching (Berthrong et al. 2009), nutrient mineralization (Smolander et al. 2000), soil organic carbon losses (Bárcena et al. 2014), soil temperature reduction or elevation (Sutton 1993), and changes in microbial activity (Merino et al. 2004). These factors are all known to affect seedling establishment and growth, and changes in soil characteristics induced by MSP have been shown to possibly last for decades (Sutinen et al. 2019).
The MSP treatments used in our study differed in terms of their direct impacts on soil structure. Visual soil observations performed in the experimental sites (Collet et al. 2020) showed that the lightweight tools, DS and MS, loosened the soil horizons down to a depth of 30 and 45 cm, respectively, and completely removed the upper organic soil horizons. The other MSP tools used in our study reached a depth of only 20 cm and did not remove the organic horizons. Soil compaction on wheel and caterpillar tracks induced by the tool carrier were limited in lightweight MSP treatments, compared to conventional MSP treatments. However, in waterlogged sites, most lightweight and conventional tools induced soil smearing at the soil-tool interface. A companion study (Dassot and Collet 2020) performed in one experimental site of the present study (site BL, with P. aquilinum) showed that root systems of both pines and oaks were slightly larger in the herbicide than in the lightweight treatment after four years, and much smaller in the control.
Since lightweight tools provided efficient control of the neighbouring vegetation and offered a larger volume of loosened soil around the seedling rooting zone, we expected that they would enhance seedling performance, compared to herbicide treatments. Somewhat surprisingly, seedling growth did not statistically differ between lightweight tools and herbicide treatments in most experimental sites. In the only site where statistically significant differences were observed, differences occurred in favour of the herbicide treatment. These observations suggest an overall effect of MSP on soil characteristics that is negative towards seedling growth.
We think that the intervention of lightweight tools made it possible to adequately control competing vegetation but, in parallel, may have degraded soil structural, chemical and biological fertility. Investigating the impacts of MSP on the various soil properties that may in turn affect seedling establishment and growth over the long term would be necessary to identify the best compromise, allowing sufficient vegetation control while preserving long-term soil fertility.
4.2 Impacts of subsoiling-mounding for pine species
Our study confirmed the positive effect of mounding (tested in treatment DS+MS) on seedling growth and survival compared to an untreated control and other MSP treatments, both in waterlogged and drier sites for pines and oak species, in agreement with previous studies (Bolte and Löf 2010; Sikström et al. 2020). In waterlogged sites, the main factor expected to impair seedling growth is the deficit of oxygen available for the roots (Drew and Lynch 1980). The main objective of mounding is thus to plant seedlings above the natural ground level. Mounding also increases soil temperature and reduces competing vegetation cover, potentially leading to a higher nutrient availability for seedlings (Sutton 1993). Our study shows the importance of reducing the competition caused by vegetation more clearly than the effect of relieving the waterlogging constraint.
In contrast, the DS tool scalps and removes the upper soil and creates a small depression where the water may stagnate in waterlogged sites. In experimental site RE where waterlogging was the most intense, visual observations made during the winter and spring seasons showed that the DS treatment increased the waterlogging constraint by lowering planting spot elevation. Site RE was the only experimental site where DS performed worse than the untreated control, with a lower height and diameter of pine seedlings after four growing seasons. Such detrimental effects of scalping on seedling survival and growth in sites with high water tables have been previously reported (Pearson et al. 2011).
In dry sites, a major threat for young seedlings is the summer drought during the first growing season (Grossnickle 2005). In such sites, mounding may increase summer drought, due to higher soil temperatures and deeper wet soil horizons during the summer. However, our study shows that mounding also performs very well on dry P. aquilinum sites. These results might be explained by a higher volume of decompacted soil after mounding preparation and by a decrease in neighbouring vegetation cover (Sutton 1993).
4.3 Seedling survival responses to MSP
Post-planting stress very often causes high seedling mortality rates during the first years (Burdett 1990). Recently planted seedlings usually suffer from a bad root-to-shoot balance, leading to a greater water stress and an insufficient supply of carbohydrates (Vyse 1981). Site conditions such as neighbouring vegetation cover, soil water content, soil texture and compaction, as well as nursery and transfer to plantation site conditions, may aggravate post-planting stress and negatively impact seedling growth and survival. A few months are generally necessary for seedlings to grow enough roots to be able to sustain normal growth rates.
Our study shows that seedling mortality in the first year is extremely variable depending on treatment, experimental site and year. However, after four years, the variability is lower and mortality rates show clearer trends, where seedlings in the MSP treatments have higher survival rates. We hypothesize that post-planting stress during the first year led to higher seedling sensitivity to various adverse events that occurred in the experimental sites (late frosts, heavy winter frosts, cockchafer attacks, rabbit herbivory, severe summer droughts), as suggested by Cole et al. (2018) and Luoranen et al. (2018). These events acted as a series of filtering effects on seedling survival, the recently transplanted seedlings seeming to be unable to overcome their occurrence (Grossnickle 2012). Each of these events has a low probability to occur, and when it occurs, it induces a high mortality rate, making the overall mortality in each experimental site and each year very difficult to predict.
The idiosyncratic nature of seedling mortality during the first year contrasts with the mortality over four years, which usually shows gradual responses. In our study, treatment effects after four years were quite similar among experimental sites. During this period, adverse events also occurred but they had a lower impact than in the first year and, as a consequence, they acted in a cumulative way on seedling survival. Once the seedlings establish and manage to adapt to the forest ecosystem, most of them are usually able to survive the individual adverse events. Seedling survival then becomes more closely correlated to overall treatment and environmental conditions (Pinto et al. 2011).
From an experimental perspective, our results clearly illustrate the difficulty to obtain a robust prediction of first-year seedling mortality in relation to MSP methods and experimental site characteristics. Such a goal would require a very large number of experimental site and year replicates and would lead to predictions with large confidence intervals (Sikström et al. 2020). On the contrary, survival after a period of three to five years is a good indicator of the performance of MSP methods, and robust estimations may be obtained by measuring a small number of experimental site or year replicates.
4.4 Conclusion and perspectives
Our study shows that lightweight MSP tools that provide intensive localised preparation perform well in sites where highly competitive and deep-rooted species such as P. aquilinum and M. caerulea may limit seedling establishment and growth, especially for pine species. In these site conditions, there is currently no effective vegetation management method: conventional MSP methods do not provide an effective vegetation control and, in many countries, herbicides are prohibited. Our study shows that the lightweight tools provided better vegetation control than conventional MSP methods (disk harrow, mouldboard plow), consistent with our first hypothesis (H1). In keeping with our second hypothesis (H2), seedling growth and survival after four growing seasons were considerably improved when combining the two lightweight tools (treatments DS+MS) compared to untreated control or to conventional MSP methods (disk harrow, mouldboard plow). However, contrary to the hypothesis (H2), the deep scarifier (treatment DS) did not always lead to better seedling performances than the conventional MSP methods that were tested. Finally, according to our third hypothesis (H3), pine seedling responses to MSP treatments were more pronounced than oak seedling responses.
The effective performances of the tools with regard to seedling establishment and growth are also associated with disturbances in soil properties in the seedling vicinity (removal of the upper soil layers, decompaction of deeper soil horizons and, in the mounding treatment, displacement of soil volume to create the mound), like for any MSP method. Such disturbances may affect long-term soil chemical and physical properties (Sutinen et al. 2019) and should be kept to a minimum to maintain the ecosystem services provided by forest soils (Scott et al. 2014). Lightweight tools may be used for intermittent or spotted soil preparation, which limits the disturbed area in the stand (Sikström et al. 2020). The issue is then to determine how much of the area around the seedlings should be prepared to allow rapid seedling establishment and growth while limiting the overall soil disturbance induced by MSP estimated at stand scale. The choice of a MSP method should be made according to several criteria, including the cost of operations and their impacts on the environment. More investigations are presently required to evaluate the economic and environmental performances of lightweight MSP tools, in addition to their impacts on seedling growth and survival in order to provide a complete evaluation of tool performances.
Openness of research data
Data are available on request from the corresponding author.
Authors‘ contribution
JPiat, CR and CC conceived the research question and designed the experiments. ND, MD, and CC conducted the data analysis. All authors contributed to data acquisition and scientific writing of the work.
Acknowledgements
We thank Florian Vast, Erwin Thirion, Lindsay Godard and all the technicians who provided their technical support (INRAE), Jean-Yves Fraysse (FCBA), as well as the numerous colleagues from the ONF for their help when running the experimental sites and collecting the data. Noémie Pousse (ONF) assisted with soil description and analysis.
Funding
This study was supported by the Ministère de l’Agriculture et de l’Alimentation (agreements E13/2010, E21/2013, E09/2017), the Région Grand-Est (agreement Alsace 871-10-C1) and the Agence de l’Environnement et la Maîtrise de l’Energie (Capsol project).
Conflict of interest
The authors declare that they have no conflict of interest.
References
Ammer C, Balandier P, Bentsen NS, Coll L, Löf M (2011) Forest vegetation management under debate: an introduction. Eur J For Res 130: 1–5. https://doi.org/10.1007/s10342-010-0452-6.
Ampoorter E, De Frenne P, Hermy M, Verheyen K (2011) Effects of soil compaction on growth and survival of tree saplings: a meta-analysis. Basic Appl Ecol 12: 394–402. https://doi.org/10.1016/j.baae.2011.06.003.
Aust WM, Miwa M, Burger JA, Patterson SC, Carter EA (2004) Wet-weather timber harvesting and site preparation effects on Coastal Plain sites: a review. South J Appl For 28: 137–151. https://doi.org/10.1093/sjaf/28.3.137.
Balandier P, Collet C, Miller JH, Reynolds PE, Zedaker SM (2006) Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 79: 3–27. https://doi.org/10.1093/forestry/cpi056.
Bárcena TG, Kiær LP, Vesterdal L, Stefánsdóttir HM, Gundersen P, Sigurdsson BD (2014) Soil carbon stock change following afforestation in Northern Europe: a meta-analysis. Glob Chang Biol 20: 2393–2405. https://doi.org/10.1111/gcb.12576.
Berthrong ST, Jobbágy EG, Jackson RB (2009) A global meta-analysis of soil exchangeable cations, pH, carbon, and nitrogen with afforestation. Ecol Appl 19: 2228–2241. https://doi.org/10.1890/08-1730.1.
Bock MD, Van Rees KCJ (2002) Mechanical site preparation impacts on soil properties and vegetation communities in the Northwest Territories. Can J For Res 32: 1381–1392. https://doi.org/10.1139/x02-067.
Bolte A, Löf M (2010) Root spatial distribution and biomass partitioning in Quercus robur L. seedlings: the effects of mounding site preparation in oak plantations. Eur J For Res 129: 603–612. https://doi.org/10.1007/s10342-010-0360-9.
Boutte B (2015) Suivi spécifique national “plantation de l’année”. Bilan des saisons 2013 et 2014. [Annual national survey of current-year plantations. 2013 and 2014 report]. Département Santé des Forêts, Ministère de l’Agriculture, de l’Agroalimentaire et de la Forêt.
Boutte B (2020) Bilan sanitaire des plantations forestières de l’année 2019. [Sanitary assessment of forest plantations in 2019]. Département Santé des Forêts, Ministère de l’Agriculture et de l’Alimentation.
Brys R, Jacquemyn H, De Blust G (2005) Fire increases aboveground biomass, seed production and recruitment success of Molinia caerulea in dry heathland. Acta Oecol 28: 299–305. https://doi.org/10.1016/j.actao.2005.05.008.
Burdett AN (1990) Physiological processes in plantation establishment and the development of specifications for forest planting stock. Can J For Res 20: 415–427. https://doi.org/10.1139/x90-059.
Burton P, Bedford L, Goldstein M, Osberg M (2000) Effects of disk trench orientation and planting spot position on the ten-year performance of lodgepole pine. New For 20: 23–44. https://doi.org/10.1023/A:1006796412006.
Cambi M, Hoshika Y, Mariotti B, Paoletti E, Picchio R, Venanzi R, Marchi E (2017) Compaction by a forest machine affects soil quality and Quercus robur L. seedling performance in an experimental field. For Ecol Manag 384: 406–414. https://doi.org/10.1016/j.foreco.2016.10.045.
Chytry M, Pysek P, Tichy L, Knollova I, Danihelka J (2005) Invasions by alien plants in the Czech Republic: a quantitative assessment across habitats. Presla 77: 339–354.
Cole E, Lindsay A, Newton M, Bailey JD (2018) Eight-year performance of bareroot Douglas-fir and bareroot and plug western larch seedlings following herbicide applications, northeast Oregon, USA. New For 49: 791–814. https://doi.org/10.1007/s11056-018-9653-5.
Collet C, Vast F, Richter C, Koller R (2020) Cultivation profile: a visual evaluation method of soil structure adapted to the analysis of the impacts of mechanical site preparation in forest plantations. Eur J For Res 140: 65–76. https://doi.org/10.1007/s10342-020-01315-2.
Cribari-Neto F, Zeileis A (2010) Beta regression in R. J Stat Softw 34: 1–24. https://doi.org/10.18637/jss.v034.i02.
Dassot M, Collet C (2020) Effects of different site preparation methods on the root development of planted Quercus petraea and Pinus nigra. New For 52: 17–30. https://doi.org/10.1007/s11056-020-09781-7.
Dolling AHU (1999) The vegetative spread of Pteridium aquilinum in a hemiboreal forest – invasion or revegetation? For Ecol Manag 124: 177–184. https://doi.org/10.1016/S0378-1127(99)00056-0.
Drew MC, Lynch JM (1980) Soil anaerobiosis, microorganisms, and root function. Ann Rev Phytopathol 18: 37–66. https://doi.org/10.1146/annurev.py.18.090180.000345.
Duncker PS, Raulund-Rasmussen K, Gundersen P, Katzensteiner K, De Jong J, Ravn HP, Smith M, Eckmüllner O, Spiecker H (2012) How forest management affects ecosystem services, including timber production and economic return: synergies and trade-offs. Ecol Soc 17, article id 50. https://doi.org/10.5751/ES-05066-170450.
Fernandez M, Malagoli P, Vernay A, Améglio T, Balandier P (2020) Below-ground nitrogen transfer from oak seedlings facilitates Molinia growth: 15N pulse-chase labelling. Plant Soil 449: 343–356. https://doi.org/10.1007/s11104-020-04473-9.
Fischer HS (2015) On the combination of species cover values from different vegetation layers. Appl Veg Sci 18: 169–170. https://doi.org/10.1111/avsc.12130.
Gaudio N, Balandier P, Philippe G, Dumas Y, Jean F, Ginisty C (2011) Light-mediated influence of three understorey species (Calluna vulgaris, Pteridium aquilinum, Molinia caerulea) on the growth of Pinus sylvestris seedlings. Eur J For Res 130: 77–89. https://doi.org/10.1007/s10342-010-0403-2.
von der Gönna MA (1992) Fundamentals of mechanical site preparation. FRDA Report 178, British Columbia Ministry of Forests.
Grossnickle SC (2005) Importance of root growth in overcoming planting stress. New For 30: 273–294. https://doi.org/10.1007/s11056-004-8303-2.
Grossnickle SC (2012) Why seedlings survive: influence of plant attributes. New For 43: 711–738. https://doi.org/10.1007/s11056-012-9336-6.
Hallsby G, Örlander G (2004) A comparison of mounding and inverting to establish Norway spruce on podzolic soils in Sweden. Forestry 77: 107–117. https://doi.org/10.1093/forestry/77.2.107.
Kees G (2008) Using subsoiling to reduce soil compaction. United States Department of Agriculture Forest Service Technology & Development Program.
Lacey ST, Brennan PD, Parekh J (2001) Deep may not be meaningful: cost and effectiveness of various ripping tine configurations in a plantation cultivation trial in eastern Australia. New For 21: 231–248. https://doi.org/10.1023/A:1012283106140.
Length R (2020) emmeans: estimated marginal means, aka least-squares means. R package version 1.4.7. https://CRAN.R-project.org/package=emmeans.
Löf M, Dey DC, Navarro RM, Jacobs DF (2012) Mechanical site preparation for forest restoration. New For 43: 825–848. https://doi.org/10.1007/s11056-012-9332-x.
Luoranen J, Saksa T, Lappi J (2018) Seedling, planting site and weather factors affecting the success of autumn plantings in Norway spruce and Scots pine seedlings. For Ecol Manag 419–420: 79–90. https://doi.org/10.1016/j.foreco.2018.03.040.
McCarthy N, Bentsen NS, Willoughby I, Balandier P (2011) The state of forest vegetation management in Europe in the 21st century. Eur J For Res 130: 7–16. https://doi.org/10.1007/s10342-010-0429-5.
Merino A, Fernández-López A, Solla-Gullón F, Edeso JM (2004) Soil changes and tree growth in intensively managed Pinus radiata in northern Spain. For Ecol Manag 196: 393–404. https://doi.org/10.1016/j.foreco.2004.04.002.
Neaves CM, Aust WM, Bolding MC, Barrett SM, Trettin CC, Vance E (2017) Soil properties in site prepared loblolly pine (Pinus taeda L.) stands 25years after wet weather harvesting in the lower Atlantic coastal plain. For Ecol Manag 404: 344–353. https://doi.org/10.1016/j.foreco.2017.08.015.
Nilsson U, Luoranen J, Kolström T, Örlander G, Puttonen P (2010) Reforestation with planting in northern Europe. Scand J For Res 25: 283–294. https://doi.org/10.1080/02827581.2010.498384.
Pearson M, Saarinen M, Minkkinen K, Silvan N, Laine J (2011) Mounding and scalping prior to reforestation of hydrologically sensitive deep-peated sites: factors behind Scots pine regeneration success. Silva Fenn 45: 647–667. https://doi.org/10.14214/sf.98.
Pinheiro J, Bates D, DebRoy S, Sarkar D (2019) nlme: linear and nonlinear mixed effects models. R package version 3.1-141. https://CRAN.R-project.org/package=nlme.
Pinto JR, Marshall JD, Dumroese RK, Davis AS, Cobos DR (2011) Establishment and growth of container seedlings for reforestation: a function of stocktype and edaphic conditions. For Ecol Manag 261: 1876–1884. https://doi.org/10.1016/j.foreco.2011.02.010.
Quibel E (2020) Impacts de la préparation du sol lors de la phase de plantation sur la dynamique des matières organiques et le stockage du carbone dans les sols forestiers. [Impacts of soil preparation prior to planting on soil organic matter dynamics in forest soils]. Ph Thesis University of Rouen.
R Core Team (2018) A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Scott DA, Eaton RJ, Foote JA, Vierra B, Boutton TW, Blank GB, Johnsen K (2014) Soil ecosystem services in loblolly pine plantations 15 years after harvest, compaction, and vegetation control. Soil Sci Soc Am J 78: 2032–2040. https://doi.org/10.2136/sssaj2014.02.0086.
Sikström U, Hjelm K, Holt Hanssen K, Saksa T, Wallertz K (2020) Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fenn 54, article id 10172 https://doi.org/10.14214/sf.10172.
Smith C, Johnston M (2001) Managing soil compaction to ensure long-term site productivity in South African forest plantations. Proc Afr Sug Technol Ass 75: 129–136.
Smolander A, Paavolainen L, Mälkönen E (2000) C and N transformations in forest soil after mounding for regeneration. For Ecol Manag 134: 17–28. https://doi.org/10.1016/S0378-1127(99)00242-X.
Sutherland B, Foreman FF (2000) Black spruce and vegetation response to chemical and mechanical site preparation on a boreal mixedwood site. Can J For Res 30: 1561–1570. https://doi.org/10.1139/x00-087.
Sutinen R, Gustavsson N, Hänninen P, Middleton M, Räisänen ML (2019) Impact of mechanical site preparation on soil properties at clear-cut Norway spruce sites on mafic rocks of the Lapland Greenstone Belt. Soil Tillage Res 186: 52–63. https://doi.org/10.1016/j.still.2018.10.013.
Sutton RF (1993) Mounding site preparation: a review of European and North American experience. New For 7: 151–192. https://doi.org/10.1007/BF00034198.
Taylor K, Rowland AP, Jones HE (2001) Molinia caerulea (L.) Moench. J Ecol 89: 126–144. https://doi.org/10.1046/j.1365-2745.2001.00534.x.
Thiffault N, Roy V (2011) Living without herbicides in Québec (Canada): historical context, current strategy, research and challenges in forest vegetation management. Eur J For Res 130: 117–133. https://doi.org/10.1007/s10342-010-0373-4.
du Toit B, Smith CW, Little KM, Boreham G, Pallett RN (2010) Intensive, site-specific silviculture: manipulating resource availability at establishment for improved stand productivity. A review of South African research. For Ecol Manag 259: 1836–1845. https://doi.org/10.1016/j.foreco.2009.07.015.
Vernay A, Balandier P, Guinard L, Améglio T, Malagoli P (2016) Photosynthesis capacity of Quercus petraea (Matt.) saplings is affected by Molinia caerulea (L.) under high irradiance. For Ecol Manag 376: 107–117. https://doi.org/10.1016/j.foreco.2016.05.045.
Vyse A (1981) Growth of young spruce plantations in interior British Columbia. Forest Chron 57: 174–180. https://doi.org/10.5558/tfc57174-4.
Wagner RG, Robinson AP (2006) Critical period of interspecific competition for four northern conifers: 10-year growth response and associated vegetation dynamics. Can J For Res 36: 2474–2485. https://doi.org/10.1139/x06-058.
Wagner RG, Little KM, Richardson B, Mcnabb K (2006) The role of vegetation management for enhancing productivity of the world’s forests. Forestry 79: 57–79. https://doi.org/10.1093/forestry/cpi057.
Yildiz O, Eşen D, Sarginci M (2009) Long-term site productivity effects of different Rhododendron control methods in eastern beech (Fagus orientalis Lipsky) ecosystems in the Western Black Sea region of Turkey. Soil Use Manage 25: 28–33. https://doi.org/10.1111/j.1475-2743.2008.00190.x.
Total of 58 references.
