Göran Bergqvist  ,
Roger Bergström,
Märtha Wallgren
,
Roger Bergström,
Märtha Wallgren
Recent browsing damage by moose on Scots pine, birch and aspen in young commercial forests – effects of forage availability, moose population density and site productivity
Bergqvist G., Bergström R., Wallgren M. (2014). Recent browsing damage by moose on Scots pine, birch and aspen in young commercial forests – effects of forage availability, moose population density and site productivity. Silva Fennica vol. 48 no. 1 article id 1077. https://doi.org/10.14214/sf.1077
Highlights
- Moose damage was most common on aspen and least common on Scots pine
- Damage to Scots pine increased when the amount of pine browse decreased, moose index and site index increased and when birch was overtopping pine
- Damage to birch increased when moose index increased and damage to aspen increased when the amount of pine and birch browse decreased.
Abstract
Moose browsing damage from the winter preceding the study (recent damage) on Scots pine, birch and aspen was examined in relation to forage availability, an index of moose population density and site productivity in young forests in the hemiboreal zone. Recent damage was observed for 4.1 ± 0.54% (mean ± SE; Scots pine), 16.8 ± 1.89% (birch) and 67.6 ± 13.76% (aspen) of the trees. A multiple regression with five independent variables explained 19% (Scots pine) 14% (birch) and 33% (aspen) of the variation in recent damage. Cover of Scots pine browse was the most important variable for predicting damage to Scots pine and accounted for 44% of the explained variation. When birch was overtopping pine there was a significant increase in damage to pine. Moose index was the only significant variable to explain recent damage to birch, and accounted for 64% of the explained variation. For aspen, damage was negatively correlated to coverage of Scots pine and birch browse, each variable accounting for 38% of the explained variation. For Scots pine, increasing the number of pines ha–1 and performing pre-commercial thinning in such a way that pines are not overtopped may be efficient ways of reducing damage proportions, whereas birch needs to be protected from moose (by a reduction of the moose population or otherwise) in order to escape damage. Increased amounts of Scots pine browse and birch browse may also reduce damage levels to aspen, according to this study.
Keywords
Pinus;
Betula;
Populus;
Alces alces
-
Bergqvist,
Swedish Association for Hunting and Wildlife Management, Öster Malma, SE-611 91 Nyköping, Sweden
E-mail
goran.bergqvist@jagareforbundet.se
- Bergström, Forestry Research Institute of Sweden, Uppsala Science Park, SE-751 83 Uppsala, Sweden; (present) Gropgränd 2 A, SE-753 10 Uppsala, Sweden E-mail larsrogerbergstrom@yahoo.com
- Wallgren, Forestry Research Institute of Sweden, Uppsala Science Park, SE-751 83 Uppsala, Sweden E-mail martha.wallgren@skogforsk.se
Received 12 December 2013 Accepted 14 January 2014 Published 23 January 2014
Views 170513
Available at https://doi.org/10.14214/sf.1077 | Download PDF

1 Introduction
In winter, moose (Alces alces L.) mainly feed on woody plants (Bergström and Hjeljord 1987). In Sweden, the deciduous species rowan (Sorbus aucuparia L.), aspen (Populus tremula L.) and willows (Salix spp.) are ranked among the most preferred species, and they are in turn followed by silver birch (Betula pendula Roth), downy birch (Betula pubescens Ehrh.) and the conifer Scots pine, Pinus sylvestris L. (Månsson et al. 2007b). Although Scots pine ranks intermediate with regard to preference, it is quantitatively the most important winter food species for moose in Sweden (Cederlund et al. 1980; Bergström and Hjeljord 1987).
Of the tree species mentioned above, birch and especially Scots pine is of great economic importance for the Swedish forest industry. Extensive moose browsing is regarded as a problem since it may affect the suitability of trees for future industrial purposes due to lowered wood quality or suppressed volume growth and thereby result in economic losses (Lavsund 1987; Liberg et al. 2010). Highly preferred food species like rowan, aspen and willows are generally of lower economic value for the forestry sector, but are considered valuable for biodiversity (Cassing 2009).
The extent of browsing on young trees is depending on the density of the moose population (Bergström and Vikberg 1992; Hörnberg 2001b) although stand- or tree characteristics, such as stand density (Lyly and Saksa 1992), site productivity (Ball et al. 2000), tree species composition (Danell et al. 1991a; Härkönen et al. 1998; Milligan and Koricheva 2013) or silvicultural treatments (Edenius et al. 2002) may affect this relationship. Earlier browsing history may also be important as the browsing by moose on already browsed trees (in terms of number of browsed trees) is more common than expected from a random use (Bergqvist et al. 2003).
From a forest management standpoint, browsing by herbivores on important tree species can be divided into two main categories: browsing that is expected to affect the quality of the butt log (browsing of the apical leader and breaking or bark stripping of the trunk) and browsing that may affect the growth of the tree but not the quality of the butt log (i.e. browsing of lateral shoots). Damage survey methods commonly used in Sweden today focus on the former category.
Presently, a new system for moose management is being launched in Sweden. Compared to earlier systems, management goals, such as the size of the moose winter population, will be set at a larger spatial scale (for moose management areas; between 50 000 and > 1 million ha) and predictions for the amount of available browse in such areas are being developed in order to aid in decision-making. Increased knowledge regarding effects of browse availability and moose population density on browsing damage to commercially important tree species will therefore be of increasing importance.
The aim of our study was to evaluate effects of browse availability, site productivity and moose population density on recent moose browsing damage for Scots pine, birch and aspen trees in young, commercial forests. For that purpose we used data from a large-scale survey of browsing damage, performed according to standard methods in Sweden, and supplemented with data regarding browse availability and moose pellet counts.
2 Materials and methods
2.1 Survey area
The survey was conducted on forest land owned by the Holmen company, within the counties of Uppsala, Stockholm, Södermanland, Östergötland and Kalmar in Sweden (57°36´–60°10´N, 15°30´–18°40´E, altitude 5–155 m a.s.l.), in the hemiboreal forest zone (sensu Ahti et al. 1968). In this area Holmen owns about 86 000 ha of productive forest land, and at the time of the survey about 11 000 ha (13%) was covered by forest stands with a mean tree height of 1–4 m.
The total moose harvest during the hunting season 2000–2001 was 14 114 animals in the five counties, varying from 1658 in Stockholm to 3974 in Östergötland (Statistical Yearbook of Sweden 2006). This was equivalent to 0.3–0.4 harvested animals km–2 of the areas registered for moose hunting by the County Administrative Boards. Based on these figures, we estimated the moose population in the area at 0.6–0.9 animals km–2 during the winter of 2000–2001.
2.2 Field work
A total of 182 squares of forest land, each 1 x 1 km in size, were randomly distributed over the area (hereafter denoted “tracts”). Within each tract, a maximum of five stands were sampled for the survey. The criteria for a stand to be chosen were: average tree height 1–4 m., stand size at least 0.5 ha and the proportion of deciduous trees other than birch less than 30% based on stem numbers. If more than five stands in a tract fulfilled the conditions, a sub-sampling was performed. In total 450 stands were included in the survey, see Table 1.
| Table 1. Number of stands, plots and total area in the 182 surveyed tracts. Arithmetic mean values with one SE of the mean within brackets. | |||
| Total | per stand | per tract | |
| Stands (no) | 450 | 2.5 (0.09) | |
| Plots (no) | 2052 | 4.6 (0.17) | 11.3 (0.51) |
| Area (ha) | 2134 | 4.7 (0.22) | 11.7 (0.64) |
The survey was conducted between snow melt and leaf flush during April–May 2001 by personnel from the County Forestry Boards. All field workers were experienced in this type of work and attended a one day course before the survey. A grid was superimposed over each stand in such a manner that 1–15 grid intersections occurred within the stand. Circular plots (r = 3.5 m, area 38.5 m2) were systematically laid out at grid intersections. The number of plots per stand increased with increasing stand size and increasing proportion of Scots pine.
On each plot, trees of Scots pine, birch and aspen were checked for recent browsing damage (i.e. from the winter of 2000–2001). For Scots pine, browsing damage included browsing of the apical leader and/or breaking or bark stripping of the trunk, whereas it included browsing of the apical leader or breaking of the trunk for birch and aspen. These types of browsing are expected to negatively affect the height development and/or quality of the butt log, and therefore included in standard damage surveys in Sweden. To be included in the survey, Scots pine and birch trees should be taller than half the average height of the two tallest conifers on the plot, whereas aspen was included when its height was 1–2.5 m. In total, 19 781 trees were included in the survey of recent browsing damage; 10 338 Scots pine, 9110 birch and 333 aspen.
Amounts of available browse of Scots pine, birch, rowan, aspen and willows were estimated as the vertically projected area in the plot that was covered by each species within moose browsing range (approximately 0.3–3.0 m height; see Hörnberg 2001a). Cover of rowan, aspen and willows occurred only at low frequencies and they were all positively and significantly correlated to each other (Spearman correlation, rs = 0.17–0.39, p < 0.02 in all cases). They were therefore pooled together in one group, hereafter denoted “RAW”. Norway spruce (Picea abies L. Karst.) is seldom used as food by moose and was excluded from all analyses (Cederlund et al. 1980).
In addition, moose fecal pellet groups from the winter 2000–2001 were counted on circular plots (r = 5.63 m, area 100 m2), using the same plot centers as for the 38.5 m2 plots. Groups containing at least 20 pellets were counted and at least half of the pellet group should be inside the plot border to be included. The age of a pellet group (i.e. from the winter immediately before the survey or older) was determined based on pellet color and texture, as well as pellet position in relation to ground litter and vegetation (e.g. above or below fallen leaves or grass). A total of 1062 pellet groups were recorded.
Each stand was supplemented with information of site index (expected height of Scots pine or Norway spruce at 100 years age (m), an indirect measure of site productivity) from the forest owner database.
2.3 Calculation of variables
Tract mean values were used in all analyses. Site index was calculated by summing the site indices for all stands in a tract and dividing it by the total number of stands examined in that tract. The number of moose fecal pellet groups (hereafter denoted “moose index” and used as an index of the density of the moose population) was calculated as the total number of pellet groups found in a tract divided by the total number of plots examined in that tract. Similarly, average cover of browse for a species or group of species (i.e., the vertically projected area in the plots that were covered by Scots pine, birch or RAW) was calculated as the sum of percentage cover of that species in all plots in a tract divided by the total number of plots examined in that tract.
Average proportions of trees of a species (Scots pine, birch, aspen) with recent browsing damage for each tract was calculated as the total number of trees with recent browsing damage in a tract divided by the total number of trees of that species examined in the tract, and multiplied by 100.
For calculation of the overall proportion of trees with recent browsing damage for a species, the tract mean values (above) were weighted with the total number of trees of that species examined in that tract.
2.4 Statistical analyses
Spearman rank order correlation was used to test the strength of the relation among species for average browse cover and recent browsing damage, and also between recent browsing damage to Scots pine and the height-relation between pine and birch.
The effects of site index, moose index and browse cover (Scots pine, birch, RAW) on recent browsing damage to Scots pine, birch and aspen were examined using linear regressions. All variables except site index showed negative exponential distributions and were [log10 (n + 1)] transformed before analysis (Sabin and Stafford 1990). Akaike relative weights for independent variables were calculated according to Anderson (2008), and the relative weights were normalized (i.e., to sum up to 1.0 for each model).
Akaike’s Information Criteria with a second-order bias correction (AICc) were calculated, using all possible combinations of independent variables as candidate models, excluding interactive terms. The models were ranked according to the ΔAICc value to find the most parsimonious model for recent browsing damage to each species. All calculations and interpretations were performed in accordance with Anderson (2008). Effects were regarded as significant at p < 0.05. All tests were performed using R 2.13.0 (R Development Core Team 2011).
3 Results
Damage from moose browsing during the previous winter, i.e. recent damage, was observed for 4.1 ± 0.54% (mean ± SE) of the Scots pine trees. Corresponding figures were 16.8 ± 1.89% and 67.6 ± 13.76% for birch and aspen, respectively. Damage proportions were significantly and positively correlated among species; Scots pine vs. birch (rs = 0.28, p < 0.001), Scots pine vs. aspen (rs = 0.42, p = 0.015) and birch vs. aspen (rs = 0.51, p = 0.029).
Scots pine browse was present in 160 (87.9%) of the surveyed tracts, and showed the highest coverage of the species or species groups examined (average cover 7.2 ± 0.51%, average height 1.3 ± 0.04 m; calculation of height only including tracts where such browse was found). Birch browse was found in 174 (95.6%) tracts, with average cover 4.5 ± 0.29% and average height 1.1 ± 0.04 m, whereas RAW species were present in 146 (80.2%) of the tracts, but only in small amounts (average cover 1.0 ± 0.1% and average height 1.0 ± 0.06 m). In a few tracts, coverage of Scots pine and birch browse exceeded 50%, whereas cover of RAW showed a maximum of 20%, Fig. 1. The cover of Scots pine browse was negatively correlated to the cover of birch browse (rs = –0.43, p = 0.006) and RAW browse (rs = –0.32, p < 0.001), whereas the correlation among birch browse and RAW browse was positive and significant (rs = 0.18, p = 0.016).
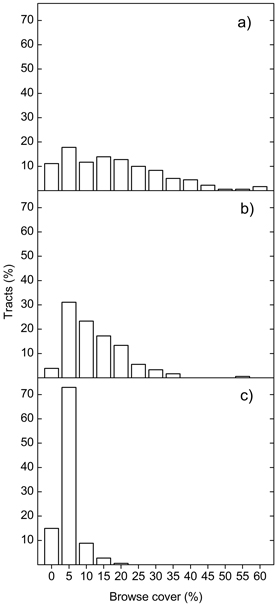
Fig. 1. Frequencies of tracts with different average cover of Scots pine (a), birch (b) and RAW (c) browse. Values on the x-axis denote class upper limits, except for the 0-class. RAW = rowan, aspen, willows.
Depending on species, a multiple regression with five independent variables at the tract scale explained between 14% (birch) and 33% (aspen) of the variation in recent browsing damage. Cover of Scots pine browse was the single most important variable for predicting browsing damage to Scots pine and accounted for 44% of the explained variation, Fig. 2. Recent browsing damage to Scots pine increased when the cover of Scots pine browse decreased, and when moose index and site index increased, Table 2.
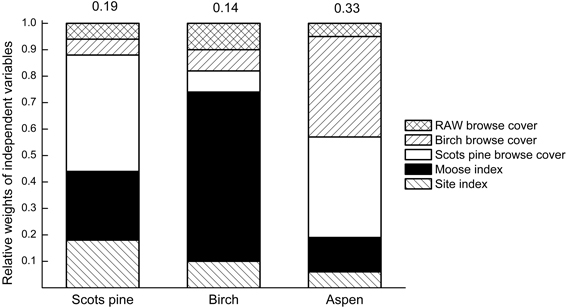
Fig. 2. Akaike relative weights for independent variables (normalized to sum up to 1.0 for each species) in multiple regressions explaining recent browsing damage by moose to Scots pine, birch and aspen. The proportion of the total variance explained by all variables combined (Adj. R2) are given above each bar. RAW = rowan, aspen, willows.
| Table 2. Summary of regression models used to explain recent moose damage to Scots pine, birch and aspen at the tract level. All independent variables except site index were [log10 (n + 1)] transformed before analysis. | ||||
| Variables | df | Coeff. | SE | p |
| Scots pine | ||||
| Intercept | 1 | –0.314 | 0.461 | 0.497 |
| Moose index | 1 | 1.068 | 0.328 | 0.001 |
| Site index | 1 | 0.042 | 0.018 | 0.025 |
| Scots pine cover | 1 | –0.461 | 0.116 | <0.001 |
| Birch cover | 1 | 0.005 | 0.146 | 0.970 |
| RAW cover | 1 | 0.100 | 0.202 | 0.621 |
| Error | 167 | |||
| Total | 173 | |||
| F | 8.735 | <0.001 | ||
| Birch | ||||
| Intercept | 1 | –0.400 | 0.588 | 0.497 |
| Moose index | 1 | 1.793 | 0.410 | <0.001 |
| Site index | 1 | 0.038 | 0.023 | 0.100 |
| Scots pine cover | 1 | 0.135 | 0.144 | 0.351 |
| Birch cover | 1 | –0.158 | 0.198 | 0.424 |
| RAW cover | 1 | –0.295 | 0.243 | 0.226 |
| Error | 162 | |||
| Total | 168 | |||
| F | 6.550 | <0.001 | ||
| Aspen | ||||
| Intercept | 1 | 4.178 | 1.963 | 0.042 |
| Moose index | 1 | 1.503 | 1.277 | 0.249 |
| Site index | 1 | –0.058 | 0.068 | 0.408 |
| Scots pine cover | 1 | –1.513 | 0.546 | 0.010 |
| Birch cover | 1 | –1.715 | 0.592 | 0.007 |
| RAW cover | 1 | 0.272 | 0.707 | 0.703 |
| Error | 26 | |||
| Total | 32 | |||
| F | 3.785 | 0.010 | ||
| Note: RAW = rowan, aspen, willows | ||||
The height relation between Scots pine and birch also affected the damage level of pine. There was a significant increase in recent browsing damage when birch was overtopping pine (rs = 0.23, p = 0.040), Fig. 3. The proportion of pines with recent browsing damage was less than 2% when pine was twice as tall as birch, whereas browsing damage was 16% when birch was at least 50% taller than pine.
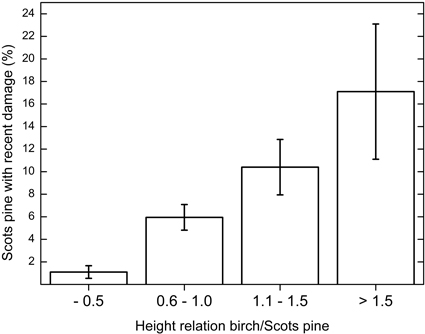
Fig. 3. Proportion of Scots pine with recent browsing damage depending on the height-relation between birch and Scots pine. When the relation is >1.0, birch is overtopping pine. Tracts with both birch and Scots pine are included in the analysis (n = 154). Vertical bars show ± one S.E. of the mean.
Moose index was the most important, and only significant, variable to explain recent browsing damage to birch, and accounted for 64% of the explained variation, Fig. 2. This type of damage increased with an increasing moose index, Table 2. For aspen, browsing damage was negatively correlated to coverage of Scots pine and birch browse, both independent variables being equally important (each accounting for 38% of the explained variation), Fig. 2 and Table 2.
According to ΔAICc, the most parsimonious regression models differed among species. However the full model, including all five independent variables, had some empirical support forall species (ΔAICc < 4), Table 3.
| Table 3. Independent variables included and ΔAICc values for predicting recent moose browsing damage on Scots pine, birch and aspen. | ||||||
| Species | Moose index | Site index | Forage cover | ΔAICc | ||
| Scots pine | Birch | RAW | ||||
| Scots pine | X | X | X | 0 | ||
| X | X | X | X | 1.72 | ||
| X | X | X | X | X | 3.72 | |
| Birch | X | 0 | ||||
| X | X | 0.61 | ||||
| X | X | X | 0.75 | |||
| X | X | X | X | 2.24 | ||
| X | X | X | X | X | 3.57 | |
| Aspen | X | X | 0 | |||
| X | X | X | 0.94 | |||
| X | X | X | X | 2.10 | ||
| X | X | X | X | X | 3.92 | |
| Note: RAW = rowan, aspen, willows | ||||||
4 Discussion
In this survey, all tree species examined (Scots pine, birch, aspen) were browsed to some extent. The rank order, in terms of percentage of trees with recent browsing damage, was aspen > birch > Scots pine. This is in line with moose browsing preferences, as reported by Bergström and Hjeljord (1987) and Månsson et al. (2007b). Similar results were reported by Bergström et al. (1995), Jalkanen (2001) and Speed et al. (2013), who also found browsing damage to be more common for silver birch compared to downy birch. Since we did not differentiate between the two birch species in this study, we cannot say if the relatively high damage proportion found for birch was due to a high proportion of silver birch, a high moose index (the only significant variable to explain browsing damage to birch) or some other discrepancy.
When interpreting the results from this study, it should be kept in mind that we have recorded browsing damage according to a Swedish standard survey method, i.e. browsing of lateral shoots is excluded for all species. However, a significant relationship between damage, as defined in this study, and percentage of browsing of all shoots has been demonstrated for Scots pine (Bergqvist et al. 2001), although the fit is not perfect (Lindqvist 2012). Top shoot browsing is the most common of the three types of damage recorded for Scots pine, and accounted for about 75% of all recent damage in a large scale survey (Bergqvist et al. 2001). Hence, we believe that results from our study may be compared to results from other studies where, e.g., lateral shoot browsing is included or breaking and/or bark stripping of the trunk are excluded.
The browse cover recorded in this investigation was larger for pine, but smaller for birch and RAW, compared to similar investigations in Sweden (Bergström et al. 1995; Hörnberg 2001a). It should be noted that those surveys comprised all forest types and also the entire country, whereas the present investigation is restricted to young, intensively managed forests in one region. The intensity of management may affect the occurrence of preferred species, as they were found in greater numbers in young forest owned by private land owners, who left more deciduous trees after pre-commercial thinning, compared to forest companies (Cassing 2009). However, since only stands with a low proportion of deciduous species other than birch were eligible for selection in this study, we cannot draw any further conclusions regarding the amounts of preferred browse found.
According to ΔAICc, the most parsimonious model explaining recent browsing damage differed among species. A model with ΔAICc < 4 may be used although models with ΔAICc > 2 have less empirical support (Burnham and Anderson 2002). Since the full model, using all five independent variables, showed ΔAICc < 4 for all species we suggest that this model may be used instead of using different models for each species. However, since the models only explained 19% (Scots pine), 14% (birch) and 33% (aspen) of the total variation, it is obvious that also other factors affect damage patterns.
Cover of Scots pine browse was the single most important factor explaining recent damage to Scots pine, and accounted for almost half of the explained variation. Similar results have been reported by Månsson et al. (2007a), who investigated total browsing and coverage (all species) at different spatial scales, and Bergström et al. (1995), who investigated browsing to Scots pine in relation to coverage of different species. Per unit cover, the amount of biomass available for browsing is approximately five times bigger for Scots pine compared to birch or RAW (Broman 2005) and, hence, Scots pine may be expected to have a stronger influence on moose habitat use compared to birch or RAW.
In our model, pine cover showed a negative coefficient (i.e. damage decreased as cover increased). We believe that this is due to the fact that we used the percentage of pines with recent damage as the response variable. A general pattern is that the absolute number of browsed/damaged trees or shoots increases with increasing food supply, whereas the proportion decreases (Lyly and Saksa 1992; Heikkilä and Härkönen 1996). As pointed out by Ball and Dahlgren (2002), this fact could be exploited in order to increase the number of undamaged pine trees at a given moose population density by increasing the number of Scots pine to 4000–5000 ha–1 in young stands, compared to recommendations of at least 1700 ha–1. However, at very high stand densities (more than 11 000 trees ha–1) the proportion of pine trees with browsing of the apical leader again increases (Lyly and Saksa 1992; Wallgren et al. 2013). There may be several reasons for this. Very dense stands offer shelter for moose even before canopy closure, which may increase the time spent in such stands (Lyly and Saksa 1992).
The amount of birch browse did not significantly affect Scots pine damage, in agreement with results by Ball and Dahlgren (2002) and Härkönen (1998), but the height relation between the species did. Damage increased when birch overtopped pine. Similar results have been reported previously (e.g. Heikkilä 1990, 1993; Heikkilä and Härkönen 1996; Härkönen 1998; Härkönen et al. 2008b; Nikula et al. 2008). Scots pine is a light demanding tree species, and shading reduces its height- and diameter growth, keeping the trees longer within moose browsing range and, possibly, making them more palatable to moose (Danell et al. 1991b). Hence, in order to avoid moose browsing damage to Scots pine, pre-commercial thinning should be carried out at such a time and in such a way that the pines are not overtopped by birch.
Moose index, showing a positive coefficient, was also a significant factor explaining browsing damage to Scots pine. The importance of moose population density on browsing/damage has been shown to be scale-dependent. At small spatial scales (e.g. tree, patch or stand), moose population density is often an important predictor of browsing pressure, whereas environmental factors such as site productivity and amount of young forest suitable for browsing becomes increasingly important as spatial scale increases (Bergström et al. 1995; Månsson et al. 2007a). In some cases, e.g. when comparing different areas over large spatial scales, moose population density does not even rank as a significant factor explaining browsing pressure (Månsson et al. 2007a; Vysinova 2010). However, when comparing forest landscapes in Sweden, Finland and Russian Karelia, Angelstam et al. (2000) reported significant correlations between moose population density and browsing damage to Scots pine. As cover of Scots pine browse, moose index and site index all were significant in the model explaining recent damage to Scots pine, we conclude that the tract scale used in this survey (one km2) can be viewed as an intermediate scale, where both factors related to the moose population and, especially, environmental factors are important.
For browsing on birch, moose index was the only independent variable in the most parsimonious model. Other studies show conflicting results, i.e. a positive correlation with the volume of Scots pine (Cassing et al. 2006) or no significant relationship between browsing on birch and moose index or browse availability (Månsson, 2009). However, Speed et al. (2013) reported a positive relation between browsing on birch and moose population density at the municipal level. Hence, it is possible that these processes are also scale-dependent.
Almost seven out of ten aspen trees were damaged by recent moose browsing in this survey. The damage proportion found is in the same order of magnitude as in other studies (Cassing et al. 2006; Härkönen et al. 2008a). An increased browsing damage to aspen was coupled to a decreased cover of Scots pine and birch, and they were the only significant independent variables in the model. Similar results have been reported by Hörnberg (2001a) who found a negative relationship between available pine forage and browsing on a set of species including aspen and by Cassing et al. (2006) who suggested this to be an effect of aspen growing on more high-productive soils where pine and birch are less abundant.
It should also be noted that there were positive and significant correlations for the proportion of trees with recent browsing damage among all species investigated. We believe that this reflects the large-scale distribution of moose in the relatively large survey area, with the total browsing pressure being generally stronger in some tracts than in others. Moose habitat selection is found to be scale-dependent and, at the landscape scale, moose select winter habitats with high available browse biomass of low quality species such as pine and birch whereas patches with high quality browse such as aspen are selected within the home range (Nikula et al. 2004; van Beest et al. 2010).
4.1 Management implications
The new moose management system in Sweden includes, among other things, the use of standard damage surveys and new methods in order to predict the development of available browse as tools in deciding on management goals. Hence, it is important to gain a better understanding on how available browse and the size of the moose population affect damage levels.
For Scots pine the concern is to produce trees with high timber quality and the survey method used is designed to account for the types of browsing that is expected to affect timber quality. Increasing the number of pines ha–1 may be a more efficient way of reducing damage proportions compared to reducing the moose population, according to results from this study. The amount of deciduous species seems to be of less importance unless the deciduous trees are overtopping pine, in which case damage levels increase.
The results regarding birch shows that, at the spatial scale used in this survey, a reduction of the moose population or other measures to protect birches from browsing are the most efficient way to reduce browsing damage.
For aspen, the main focus in management is that browsing may reduce the recruitment of mature trees, important for conservation (Cassing 2009). Although browsing may play a role, most studies conclude that silvicultural practices and the absence of forest fires seem to be the main causes of changes in aspen abundance (Zakrisson et al. 2007; Edenius et al. 2011; Myking et al. 2011).
Acknowledgements
The study was financed by Holmen, Swedish Association for Hunting and Wildlife Management and Forestry Research Institute of Sweden. We thank all people involved in the field work.
References
Ahti T., Hämet-Ahti L., Jalas J. (1968). Vegetation zones and their sections in Northwestern Europe. Annales Botanica Fennici 5: 169–211.
Anderson R.D. (2008). Model based inference in the life sciences. Springer Verlag, New York. 184 p. ISBN 978-0-387-74073-7.
Angelstam P., Wikberg P.E., Danilov P., Faber W.E., Nygren K. (2000). Effects of moose density on timber quality and biodiversity restoration in Sweden, Finland and Russian Karelia. Alces 36: 133–145. ISSN 0835-5851.
Ball J.P., Dahlgren J. (2002). Browsing damage on pine (P. sylvestris and P. contorta) by a migrating moose (Alces alces) population in winter: relation to habitat composition and road barriers. Scandinavian Journal of Forest Research 17: 427–435. http://dx.doi.org/10.1080/028275802320435441.
Ball J.P., Danell K., Sunesson P. (2000). Response of a herbivore community to increased food quality and quantity: an experiment with nitrogen fertilizer in a boreal forest. Journal of Applied Ecology 37: 247–255. http://dx.doi.org/10.1046/j.1365-2664.2000.00487.x.
Bergqvist G., Bergström R., Edenius L. (2001). Patterns of stem damage by moose (Alces alces) in young stands of Pinus sylvestris. Scandinavian Journal of Forest Research 16: 363–370. http://dx.doi.org/10.1080/02827580119307.
Bergqvist G., Bergström R., Edenius L. (2003). Effects of moose (Alces alces) rebrowsing on damage development in young stands of Scots pine (Pinus sylvestris). Forest Ecology and Management 176: 397–403. http://dx.doi.org/10.1016/S0378-1127(02)00288-8.
Bergström R., Hjeljord O. (1987). Moose and vegetation interactions in northwestern Europe and Poland. Swedish Wildlife Research, Supplement 1: 213–228. ISSN 0349-5116.
Bergström R., Vikberg M. (1992). Winter browsing on pine and birch in relation to moose population density. Alces, supplement 1: 127–131.
Bergström R., Jernelid H., Lavsund S., Lundberg P., Wallin K. (1995). Älgtäthet – betestryck – fodertillgång – skogstillstånd – skadenivåer – skaderisker. [Moose density – browsing pressure – forage availability – forest composition – forest damage – damage risks]. Projekt Balanserad Älgstam, Final report. Swedish Hunters Organisation and Swedish University of Agricultural Sciences (SLU). 24 p. [In Swedish].
Broman E. (2005). Validation of coverage degree as a measure of moose food resources. Final report. Naturvårdsverket diarie nr: 802-151-03F. 15 p. [In Swedish].
Burnham K.P., Anderson D.R. (2002). Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag. New York. 488 p. ISBN 0-387-95364-7.
Cassing G. (2009). Decidious tree occurrences and large herbivore browsing in multiscale perspectives. Licentiate thesis. Karlstad University Studies 2009:30. 28 p. ISBN 978-91-7063-253-2.
Cassing G., Greenberg L., Mikusinski G. (2006). Moose (Alces alces) browsing in young forest stands in central Sweden: a multiscale perspective. Scandinavian Journal of Forest Research 21: 221–230. http://dx.doi.org/10.1080/02827580600673535.
Cederlund G., Ljungqvist H., Markgren G., Stålfelt F. (1980). Foods of moose and roe-deer at Grimsö in central Sweden. Results of rumen content analysis. Swedish Wildlife Research 11: 169–247.
Danell K., Edenius L., Lundberg P. (1991a). Herbivory and tree stand composition: moose patch use in winter. Ecology 72: 1350–1357. http://dx.doi.org/10.2307/1941107.
Danell K., Niemelä P., Varvikko T., Vuorisalo T. (1991b). Moose browsing on Scots pine along a gradient of plant productivity. Ecology 72: 1624–1633. http://dx.doi.org/10.2307/1940962.
Edenius L., Bergman M., Ericsson G., Danell K. (2002). The role of moose as a disturbance factor in managed boreal forests. Silva Fennica 36: 57–67.
Edenius L., Ericsson G., Kempe G., Bergström R., Danell K. (2011). The effects of changing land use and browsing on aspen abundance and regeneration: a 50-year perspective from Sweden. Journal of Applied Ecology 48: 301–309. http://dx.doi.org/10.1111/j.1365-2664.2010.01923.x.
Härkönen S. (1998). Effects of silvicultural cleaning in mixed pine-deciduous stands on moose damage to Scots pine (Pinus sylvestris). Scandinavian Journal of Forest Research 13: 429–436. http://dx.doi.org/10.1080/02827589809383003.
Härkönen S., Heikkilä R., Faber W.E., Pehrson Å. (1998). The influence of silvicultural cleaning on moose browsing in young Scots pine stands in Finland. Alces 34: 409–422.
Härkönen S., Eerikäinen K., Lähteenmäki R., Heikkilä R. (2008a). Does moose browsing threaten european aspen regeneration in Koli national park, Finland? Alces 44: 31–40.
Härkönen S., Miina J., Saksa T. (2008b). Effect of cleaning methods in mixed pine-deciduous stands on moose damage to Scots pines in southern Finland. Scandinavian Journal of Forest Research 23: 491–500. http://dx.doi.org/10.1080/02827580802491371.
Heikkilä R. (1990). Effect of plantation characteristics on moose browsing on Scots pine. Silva Fennica 24: 341–351.
Heikkilä R. (1993). Effects of food quantity and tree species composition on moose (Alces alces) browsing in Scots pine plantations. Folia Forestalia 815. 18 p. [In Finnish with English summary].
Heikkilä R., Härkönen S. (1996). Moose browsing in young Scots pine stands in relation to forest management. Forest Ecology and Management 88: 179–186. http://dx.doi.org/10.1016/S0378-1127(96)03823-6.
Hörnberg S. (2001a). The relationship between moose (Alces alces) browsing utilization and the occurrence of different forage species in Sweden. Forest Ecology and Management 149: 91–102. http://dx.doi.org/10.1016/S0378-1127(00)00547-8.
Hörnberg S. (2001b). Changes in population density of moose (Alces alces) and damage to forests in Sweden. Forest Ecology and Management 149: 141–151. http://dx.doi.org/10.1016/S0378-1127(00)00551-X.
Jalkanen A. (2001). The probability of moose damage at the stand level in southern Finland. Silva Fennica 35: 159–168.
Lavsund S. (1987). Moose relationships to forestry in Finland, Norway and Sweden. Swedish Wildlife Research, Supplement 1: 229–244. ISSN 0349-5116.
Liberg O., Bergström R., Kindberg J., von Essen H. (2010). Ungulates and their management in Sweden. In: Appollonio M., Andersen R., Putman R. (eds.). European ungulates and their management in the 21 st Century. Cambridge University Press, Cambridge. p. 37–70. ISBN 978-0-521-76061-4.
Lindqvist S. (2012). Moose foraging patterns: implications for the use of browsing indices. Swedish University of Agricultural Sciences, Department of Ecology. Independent project/Degree project 2012:17.
Lyly O., Saksa T. (1992). The effect of stand density on moose damage in young Pinus sylvestris stands. Scandinavian Journal of Forest Research 7: 393–403. http://dx.doi.org/10.1080/02827589209382732.
Månsson J. (2009). Environmental variation and moose Alces alces density as determinants of spatio-temporal heterogeneity in browsing. Ecography 32: 601–612. http://dx.doi.org/10.1111/j.1600-0587.2009.05713.x.
Månsson J., Andrén H., Pehrson Å., Bergström R. (2007a). Moose browsing and forage availability: a scale-dependent relationship? Canadian Journal of Zoology 85: 372–380. http://dx.doi.org/10.1139/Z07-015.
Månsson J., Kalén C., Kjellander P., Andrén H., Smith H. (2007b). Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scandinavian Journal of Forest Research 22: 407–414. http://dx.doi.org/10.1080/02827580701515023.
Milligan H.T., Koricheva J. (2013). Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: an experimental study. Journal of Animal Ecology 82: 739–748. http://dx.doi.org/10.1111/1365-2656.12049.
Myking T., Bohler F., Austerheim G., Solberg E.J. (2011). Life history strategies of aspen (Populus tremula L.) and browsing effects: a literature review. Forestry 84: 61–71. http://dx.doi.org/10.1093/forestry/cpq044.
Nikula A., Heikkinen S., Helle E. (2004). Habitat selection of adult moose Alces alces at two spatial scales in central Finland. Wildlife Biology 10: 121–135.
Nikula A., Hallikainen V., Jalkanen R., Hyppönen M., Mäkitalo K. (2008). Modelling the factors predisposing Scots pine to moose damage in artificially regenerated sapling stands in Finnish lapland. Silva Fennica 42: 587–603.
R Development Core Team. (2011). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.R-project.org.
Sabin T.E., Stafford S.G. (1990). Assessing the need for transformation of response variables. Forest Research Laboratory, Oregon State University, Corvallis, Special Publication 20. 31 p. .
Speed J.D.M., Austrheim G, Hester A.J., Solberg E.J., Tremblay J-P. (2013). Regional-scale alteration of clear-cut forest regeneration caused by moose browsing. Forest Ecology and Management 289: 289–299. http://dx.doi.org/10.1016/j.foreco.2012.10.051.
Statistical Yearbook of Sweden. (2006). Statistiska Centralbyrån. 784 p. ISBN 91-618-1285-4. [In Swedish with English translations].
Wallgren M., Bergström R., Bergqvist G., Olsson M. (2013). Spatial distribution of browsing and tree damage by moose in young pine forest, with implications for the forest industry. Forest Ecology and Management 305: 229–238. http://dx.doi.org/10.1016/j.foreco.2013.05.057.
Van Beest F.M., Mysterud A., Loe L.E., Milner J.M. (2010). Forage quality and depletion as scale-dependent mechanisms driving habitat selection of a large browsing herbivore. Journal of Animal Ecology 79: 910–922. http://dx.doi.org/10.1111/j.1365-2656.2010.01701.x.
Vysinova L. (2010). Determinants of winter browsing intensity on young Scots pine (Pinus sylvestris) by moose (Alces alces) across a bio-geographical gradient in Sweden. Honor thesis 2010:6. Swedish University of Agricultural Sciences, Department of Wildlife, Fish and Environmental Studies. 34 p.
Zakrisson C., Ericsson G., Edenius L. (2007). Effects of browsing on recruitment and mortality of European aspen (Populus tremula L.). Scandinavian Journal of Forest Research 22: 324–332. http://dx.doi.org/10.1080/02827580701442186.
Total of 48 references
